Animal Models for In Vivo Lactation Studies: Anatomy, Physiology and Milk Compositions in the Most Used Non-Clinical Species: A Contribution from the ConcePTION Project

 ,
, 
 ,
,  , , , ,
, , , ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Anatomy of the Mammary Glands
2.1. Humans
2.2. NHPs
2.3. Pigs and Minipigs
2.4. Dogs
2.5. Mice
2.6. Rats
2.7. Rabbits
3. Physiology of Lactation
3.1. Humans
3.2. NHPs
3.3. Pigs and Minipigs
3.4. Dogs
3.5. Mice
3.6. Rats
3.7. Rabbits
4. Colostrum and Milk Composition
4.1. Humans
4.2. NHPs
4.3. Pigs and Minipigs
4.4. Dogs
4.5. Mice
4.6. Rats
4.7. Rabbits
5. Discussion
5.1. Anatomy of the Mammary Gland
5.2. Physiology of Lactation
5.3. Colostrum and Milk Composition
5.4. Practical Considerations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, A. Breast milk as the gold standard for protective nutrients. J. Pediatr. 2010, 156, S3–S7. [Google Scholar] [CrossRef]
- World Health Organization. Breastfeeding. Available online: https://www.who.int/westernpacific/health-topics/breastfeeding (accessed on 1 December 2020).
- Saha, M.R.; Ryan, K.; Amir, L.H. Postpartum women’s use of medicines and breastfeeding practices: A systematic review. Int. Breastfeed. J. 2015, 10, 1–10. [Google Scholar] [CrossRef]
- Hussainy, S.Y.; Dermele, N. Knowledge, attitudes and practices of health professionals and women towards medication use in breastfeeding: A review. Int. Breastfeed. J. 2011, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Nice, F.J.; Luo, A.C. Medications and breast-feeding: Current concepts. J. Am. Pharm. Assoc. 2012, 52, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Dodds, W.J.; Abelseth, M.K. Criteria for selecting the animal to meet the research need. Lab. Anim. Sci. 1980, 30, 460–465. [Google Scholar] [PubMed]
- Davidson, M.K.; Lindsey, J.R.; Davis, J.K. Requirements and selection of an animal model. Isr. J. Med. Sci. 1987, 23, 551–555. [Google Scholar]
- Prior, H.; Baldrick, P.; De Haan, L.; Downes, N.; Jones, K.; Mortimer-Cassen, E.; Kimber, I. Reviewing the utility of two species in general toxicology related to drug development. Int. J. Toxicol. 2018, 37, 121–124. [Google Scholar] [CrossRef]
- Singh, B. The common integument. In Dyce, Sack, and Wensing’s Textbook of Veterinary Anatomy, 5th ed.; Saunders: Philadelphia, PA, USA, 2017; pp. 341–358. ISBN 9780323442640. [Google Scholar]
- Macias, H.; Hinck, L. Mammary gland development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef]
- Cardiff, R.D.; Jindal, S.; Treuting, P.M.; Going, J.J.; Gusterson, B.; Thompson, H.J. 23—Mammary gland. In Comparative Anatomy and Histology, 2nd ed.; Treuting, P.M., Dintzis, S.M., Montine, K.S., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 487–509. ISBN 9780128029008. [Google Scholar]
- Russo, I.H.; Tewari, M.; Russo, J. Morphology and Development of the Rat Mammary Gland. In Integument and Mammary Glands; Jones, T.C., Mohr, U., Hunt, R.D., Eds.; Monographs on Pathology of Laboratory Animals; Springer: Berlin/Heidelberg, Germany, 1989; pp. 233–252. ISBN 9783642837494. [Google Scholar]
- Cardiff, R.D.; Allison, K.H. 4—Mammary Gland. In Comparative Anatomy and Histology; Treuting, P.M., Dintzis, S.M., Eds.; Academic Press: San Diego, CA, USA, 2012; pp. 41–52. ISBN 9780123813619. [Google Scholar]
- Honvo-Houeto, E.; Truchet, S. Indirect immunofluorescence on frozen sections of mouse mammary gland. J. Vis. Exp. 2015, 1, 53179. [Google Scholar] [CrossRef]
- Silver, I.A. Symposium on mammary neoplasia in the dog and cat—I the anatomy of the mammary gland of the dog and cat. J. Small Anim. Pract. 1966, 7, 689–696. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Rasotto, R.; Zappulli, V.; Goldschmidt, M.H. Development, anatomy, histology, lymphatic drainage, clinical features, and cell differentiation markers of canine mammary gland neoplasms. Vet. Pathol. 2010, 48, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, S.C.; Akers, R.M. Mammary gland anatomy. In Encyclopedia of Dairy Sciences; Fuquay, J.W., Fox, P.F., McSweeney, P.L.H., Eds.; Academic Press: San Diego, CA, USA, 2011; Volume 3. [Google Scholar]
- Szendrő, Z.; Szendrő, K.; Zotte, A.D. Management of reproduction on small, medium and large rabbit farms: A review. Asian Australas. J. Anim. Sci. 2012, 25, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Rutteman, T. Mammary glands. In Medical History and Physical Examination in Companion Animals; Rijnberk, A., van Sluijs, F.J., Eds.; Saunders: Philadelphia, PA, USA, 2008; pp. 132–134. [Google Scholar]
- Davis, S.R. Triennial lactation symposium/bolfa: Mammary growth during pregnancy and lactation and its relationship with milk yield1. J. Anim. Sci. 2017, 95, 5675–5688. [Google Scholar] [CrossRef]
- Pospieszny, N.; Poznanski, W.; Rzasa, A.; Zawada, Z. The anatomical structure of sow’s udder—A different point of view. In Book of Abstracts of the 55th Annual Meeting of The European Association For Animal Production; van der Honing, Y., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2004; p. 292. ISBN 9789076998459. [Google Scholar]
- Cline, J.M.; Wood, C.E. The Mammary Glands of Macaques. Toxicol. Pathol. 2008, 36, 130S–141S. [Google Scholar] [CrossRef] [PubMed]
- Neville, M.C. Anatomy and physiology of lactation. Pediatr. Clin. N. Am. 2001, 48, 13–34. [Google Scholar] [CrossRef]
- Gorden, P.J.; Timms, L.L. Lactation. In Duke’s Physiology of Domestic Animals; Reece, W.O., Ed.; Wiley Blackwell: Hoboken, NJ, USA, 2015; pp. 694–714. [Google Scholar]
- Djonov, V.; Andres, A.C.; Ziemiecki, A. Vascular remodelling during the normal and malignant life cycle of the mammary gland. Microsc. Res. Tech. 2001, 52, 182–189. [Google Scholar] [CrossRef]
- Hassiotou, F.; Geddes, D. Anatomy of the human mammary gland: Current status of knowledge. Clin. Anat. 2012, 26, 29–48. [Google Scholar] [CrossRef]
- Inman, J.L.; Robertson, C.; Mott, J.D.; Bissell, M.J. Mammary gland development: Cell fate specification, stem cells and the microenvironment. Development 2015, 142, 1028–1042. [Google Scholar] [CrossRef]
- Stevenson, A.J.; Vanwalleghem, G.; Stewart, T.A.; Condon, N.D.; Lloyd-Lewis, B.; Marino, N.; Putney, J.W.; Scott, E.K.; Ewing, A.D.; Davis, F.M. Multiscale imaging of basal cell dynamics in the functionally mature mammary gland. Proc. Natl. Acad. Sci. USA 2020, 117, 26822–26832. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, D.T.; Kent, J.C.; Hartmann, R.A.; Hartmann, P.E. Anatomy of the lactating human breast redefined with ultrasound imaging. J. Anat. 2005, 206, 525–534. [Google Scholar] [CrossRef]
- Russo, J.; Russo, I.H. Development of the human breast. Maturitas 2004, 49, 2–15. [Google Scholar] [CrossRef]
- Hurley, W.L. Review: Mammary gland development in swine: Embryo to early lactation. Animals 2019, 13, s11–s19. [Google Scholar] [CrossRef]
- Van Klompenberg, M.K.; Manjarin, R.; Trott, J.F.; McMicking, H.F.; Hovey, R.C. Late gestational hyperprolactinemia accelerates mammary epithelial cell differentiation that leads to increased milk yield1. J. Anim. Sci. 2013, 91, 1102–1111. [Google Scholar] [CrossRef]
- Van Klompenberg, M.; Manjarín, R.; Donovan, C.E.; Trott, J.F.; Hovey, R.C. Regulation and localization of vascular endothelial growth factor within the mammary glands during the transition from late gestation to lactation. Domest. Anim. Endocrinol. 2016, 54, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Orfanou, D.C.; Pourlis, A.; Ververidis, H.N.; Mavrogianni, V.S.; Taitzoglou, I.A.; Boscos, C.M.; Fthenakis, G.C. Histological features in the mammary glands of female dogs throughout lactation. Anat. Histol. Embryol. 2010, 39, 473–478. [Google Scholar] [CrossRef]
- Cardy, R.H. Sexual dimorphism of the normal rat mammary gland. Vet. Pathol. 1991, 28, 139–145. [Google Scholar] [CrossRef]
- Hughes, K.; Watson, C.J. Sinus-like dilatations of the mammary milk ducts, Ki67 expression, and CD3-positive T lymphocyte infiltration, in the mammary gland of wild European rabbits during pregnancy and lactation. J. Anat. 2018, 233, 266–273. [Google Scholar] [CrossRef]
- Hughes, K. Comparative mammary gland postnatal development and tumourigenesis in the sheep, cow, cat and rabbit: Exploring the menagerie. Semin. Cell Dev. Biol. 2020. [Google Scholar] [CrossRef]
- Lu, M.-H.; Anderson, R.R. Growth of the mammary gland during pregnancy and lactation in the rabbit. Biol. Reprod. 1973, 9, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Truchet, S.; Honvo-Houéto, E. Physiology of milk secretion. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 367–384. [Google Scholar] [CrossRef]
- Pang, W.W.; Hartmann, P.E. Initiation of human lactation: Secretory differentiation and secretory activation. J. Mammary Gland Biol. Neoplasia 2007, 12, 211–221. [Google Scholar] [CrossRef]
- Kulski, J.K.; Hartmann, P.E. Changes in human milk composition during the initiation of lactation. Aust. J. Exp. Biol. Med. Sci. 1981, 59, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Sjaastad, O.V.; Sand, O.; Hove, K. Reproduction. In Physiology of Domestic Animals; Scandinavian Veterinary Press: Oslo, Norway, 2010; pp. 683–734. [Google Scholar]
- Ostrom, K.M. A review of the hormone prolactin during lactation. Prog. Food Nutr. Sci. 1990, 14, 1–43. [Google Scholar]
- Ben-Jonathan, N.; LaPensee, C.R.; LaPensee, E.W. What can we learn from rodents about prolactin in humans? Endocr. Rev. 2007, 29, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Sriraman, N.K. The nuts and bolts of breastfeeding: Anatomy and physiology of lactation. Curr. Probl. Pediatr. Adolesc. Health Care 2017, 47, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P. The endocrine system. In Dukes’ Physiology of Domestic Animals; Reece, W.O., Ed.; Wiley Blackwell: Hoboken, NJ, USA, 2015; pp. 617–654. [Google Scholar]
- Vieira, A.M.; de Almeida Brasiel, P.G.; Ferreira, M.S.; Mateus, K.; Figueiredo, M.S.; Lisboa, P.C.; De Moura, E.G.; do Amaral Corrêa, J.O.; Lopes, F.C.F.; Da Silva, P.H.F.; et al. Maternal soybean diet during lactation alters breast milk composition and programs the lipid profile in adult male rat offspring. Endocrine 2018, 60, 272–281. [Google Scholar] [CrossRef]
- Görs, S.; Kucia, M.; Langhammer, M.; Junghans, P.; Metges, C. Technical note: Milk composition in mice—Methodological aspects and effects of mouse strain and lactation day. J. Dairy Sci. 2009, 92, 632–637. [Google Scholar] [CrossRef] [PubMed]
- McClellan, H.L.; Miller, S.J.; Hartmann, P.E. Evolution of lactation: Nutritionv.protection with special reference to five mammalian species. Nutr. Res. Rev. 2008, 21, 97–116. [Google Scholar] [CrossRef]
- Peaker, M.; Taylor, J.C. Milk secretion in the rabbit: Changes during lactation and the mechanism of ion transport. J. Physiol. 1975, 253, 527–545. [Google Scholar] [CrossRef] [PubMed]
- Chastant-Maillard, S.; Aggouni, C.; Albaret, A.; Fournier, A.; Mila, H. Canine and feline colostrum. Reprod. Domest. Anim. 2017, 52, 148–152. [Google Scholar] [CrossRef]
- Oftedal, O.T. Lactation in the dog: Milk composition and intake by puppies. J. Nutr. 1984, 114, 803–812. [Google Scholar] [CrossRef]
- Kooistra, H.S.; Okkens, A.C. Secretion of prolactin and growth hormone in relation to ovarian activity in the dog. Reprod. Domest. Anim. Zuchthyg. 2001, 36, 115–119. [Google Scholar] [CrossRef]
- Mol, J.A.; Selman, P.J.; Sprang, E.P.; Van Neck, J.W.; Oosterlaken-Dijksterhuis, M.A. The role of progestins, insulin-like growth factor (IGF) and IGF-binding proteins in the normal and neoplastic mammary gland of the bitch: A review. J. Reprod. Fertil. Suppl. 1997, 51, 339–344. [Google Scholar] [PubMed]
- Farmer, C.; Devillers, N.; Rooke, J.A.; Le Dividich, J. Colostrum production in swine: From the mammary glands to the piglets. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2006, 3, 16. [Google Scholar] [CrossRef]
- Hurley, W.; Farmer, C. Composition of sow colostrum and milk. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 193–230. [Google Scholar]
- Morris, M.; Stevens, S.W.; Adams, M.R. Plasma oxytocin during pregnancy and lactation in the cynomolgus monkey. Biol. Reprod. 1980, 23, 782–787. [Google Scholar] [CrossRef]
- McManaman, J.L.; Neville, M.C. Mammary physiology and milk secretion. Adv. Drug Deliv. Rev. 2003, 55, 629–641. [Google Scholar] [CrossRef]
- Wickes, I.G. A history of infant feeding. Arch. Dis. Child. 1953, 28, 151–158. [Google Scholar] [CrossRef]
- Sjaastad, O.V.; Sand, O.; Hove, K. The endocrine system. In Physiology of Domestic Animals; Scandinavian Veterinary Press: Oslo, Norway, 2010; pp. 219–258. [Google Scholar]
- Concannon, P.; Tsutsui, T.; Shille, V. Embryo development, hormonal requirements and maternal responses during canine pregnancy. J. Reprod. Fertil. Suppl. 2001, 57, 169–179. [Google Scholar]
- Hennighausen, L.; Robinson, G.W. Think globally, act locally: The making of a mouse mammary gland. Genes Dev. 1998, 12, 449–455. [Google Scholar] [CrossRef]
- Topper, Y.J.; Freeman, C.S. Multiple hormone interactions in the developmental biology of the mammary gland. Physiol. Rev. 1980, 60, 1049–1106. [Google Scholar] [CrossRef]
- Jahn, G.A.; Edery, M.; Bélair, L.; Kelly, P.A.; Djiane, J. Prolactin receptor gene expression in rat mammary gland and liver during pregnancy and lactation. Endocrinology 1991, 128, 2976–2984. [Google Scholar] [CrossRef] [PubMed]
- Koiter, T.R.; Moes, H.; Valkhof, N.; Wijkstra, S. Interaction of late pregnancy and lactation in rats. J. Reprod. Fertil. 1999, 115, 341–347. [Google Scholar] [CrossRef][Green Version]
- Langer, P. Differences in the composition of colostrum and milk in eutherians reflect differences in immunoglobulin transfer. J. Mammal. 2009, 90, 332–339. [Google Scholar] [CrossRef]
- Rocca, M.; Morford, L.L.; Blanset, D.L.; Halpern, W.G.; Cavagnaro, J.; Bowman, C.J. Applying a weight of evidence approach to the evaluation of developmental toxicity of biopharmaceuticals. Regul. Toxicol. Pharmacol. 2018, 98, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Moffat, G.J.; Retter, M.W.; Kwon, G.; Loomis, M.; Hock, M.B.; Hall, C.; Bussiere, J.; Lewis, E.M.; Chellman, G.J. Placental transfer of a fully human IGG2 monoclonal antibody in the cynomolgus monkey, rat, and rabbit: A comparative assessment from during organogenesis to late gestation. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2014, 101, 178–188. [Google Scholar] [CrossRef]
- Chastant, S.; Mila, H. Passive immune transfer in puppies. Anim. Reprod. Sci. 2019, 207, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Rooke, J.; Bland, I. The acquisition of passive immunity in the new-born piglet. Livest. Prod. Sci. 2002, 78, 13–23. [Google Scholar] [CrossRef]
- Keen, C.L.; Lönnerdal, B.; Clegg, M.; Hurley, L.S. Developmental changes in composition of rat milk: Trace elements, minerals, protein, carbohydrate and fat. J. Nutr. 1981, 111, 226–236. [Google Scholar] [CrossRef]
- Maertens, L.; Lebas, F.; Szendrő, Z. Rabbit milk: A review of quantity, quality and non-dietary affecting factors. World Rabbit Sci. 2010, 14, 205–230. [Google Scholar] [CrossRef]
- Adkins, Y.; Lepine, A.J.; Lonnerdal, B. Changes in protein and nutrient composition of milk throughout lactation in dogs. Am. J. Vet. Res. 2001, 62, 1266–1272. [Google Scholar] [CrossRef]
- Csapó, J.; Martin, T.; Csapó-Kiss, Z.; Házas, Z. Protein, fats, vitamin and mineral concentrations in porcine colostrum and milk from parturition to 60 days. Int. Dairy J. 1996, 6, 881–902. [Google Scholar] [CrossRef]
- Lönnerdal, B.; Keen, C.L.; Glazier, C.E.; Anderson, J. A longitudinal study of rhesus monkey (Macaca mulatta) milk composition: Trace elements, minerals, protein, carbohydrate, and fat. Pediatr. Res. 1984, 18, 911–914. [Google Scholar] [CrossRef]
- Mangel, L.; Ovental, A.; Batscha, N.; Arnon, M.; Yarkoni, I.; Dollberg, S. Higher fat content in breastmilk expressed manually: A randomized trial. Breastfeed. Med. 2015, 10, 352–354. [Google Scholar] [CrossRef]
- Yamawaki, N.; Yamada, M.; Kan-No, T.; Kojima, T.; Kaneko, T.; Yonekubo, A. Macronutrient, mineral and trace element composition of breast milk from Japanese women. J. Trace Elem. Med. Biol. 2005, 19, 171–181. [Google Scholar] [CrossRef]
- Kociszewska-Najman, B.; Borek-Dzieciol, B.; Szpotanska-Sikorska, M.; Wilkos, E.; Pietrzak, B.; Wielgos, M. The creamatocrit, fat and energy concentration in human milk produced by mothers of preterm and term infants. J. Matern. Neonatal Med. 2012, 25, 1599–1602. [Google Scholar] [CrossRef]
- Yonekubo, A.; Honda, S.; Okano, M.; Takahashi, K.; Yamamoto, Y. Dietary fish oil alters rat milk composition and liver and brain fatty acid composition of fetal and neonatal rats. J. Nutr. 1993, 123, 1703–1708. [Google Scholar] [CrossRef] [PubMed]
- Auestad, N.; Korsak, R.A.; Bergstrom, J.D.; Edmond, J. Milk-substitutes comparable to rat’s milk; their preparation, composition and impact on development and metabolism in the artificially reared rat. Br. J. Nutr. 1989, 61, 495–518. [Google Scholar] [CrossRef]
- Oftedal, O.T.; Iverson, S.J. Comparative analysis of nonhuman milks. A phylogenetic variation in the gross composition of milks. In Handbook of Milk Composition; Jensen, R.G., Ed.; Food Science and Technology; Academic Press: Cambridge, MA, USA, 1995; pp. 749–789. ISBN 9780123844309. [Google Scholar]
- Lönnerdal, B.; Keen, C.L.; Hurley, L.S.; Fisher, G.L. Developmental changes in the composition of Beagle dog milk. Am. J. Vet. Res. 1981, 42, 662–666. [Google Scholar] [PubMed]
- Luick, J.R.; Parker, H.R.; Andersen, A.C. Composition of beagle dog milk. Am. J. Physiol. Content 1960, 199, 731–732. [Google Scholar] [CrossRef]
- Nishikawa, I.; Kawanishi, G.; Cho, F.; Honjo, S.; Hatakeyama, T.; Wako, H. Chemical composition of cynomolgus monkey milk. Exp. Anim. 1976, 25, 253–264. [Google Scholar] [CrossRef][Green Version]
- Goto, K.; Fukuda, K.; Senda, A.; Saito, T.; Kimura, K.; Glander, K.E.; Hinde, K.; Dittus, W.; Milligan, L.A.; Power, M.L.; et al. Chemical characterization of oligosaccharides in the milk of six species of new and old world monkeys. Glycoconj. J. 2010, 27, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Hinde, K.; Power, M.L.; Oftedal, O.T. Rhesus macaque milk: Magnitude, sources, and consequences of individual variation over lactation. Am. J. Phys. Anthropol. 2009, 138, 148–157. [Google Scholar] [CrossRef]
- Osthoff, G.; Hugo, A.; De Wit, M.; Nguyen, T.; Seier, J. Milk composition of captive vervet monkey (Chlorocebus pygerythrus) and rhesus macaque (Macaca mulatta) with observations on gorilla (Gorilla gorilla gorilla) and white handed gibbon (Hylobates lar). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 152, 332–338. [Google Scholar] [CrossRef]
- Holt, C.; Jenness, R. Interrelationships of constituents and partition of salts in milk samples from eight species. Comp. Biochem. Physiol. Part A Physiol. 1984, 77, 275–282. [Google Scholar] [CrossRef]
- Young, B.E.; Borman, L.L.; Heinrich, R.; Long, J.; Pinney, S.; Westcott, J.; Krebs, N.F. Effect of pooling practices and time postpartum of milk donations on the energy, macronutrient, and zinc concentrations of resultant donor human milk pools. J. Pediatr. 2019, 214, 54–59. [Google Scholar] [CrossRef]
- Alves Peixoto, R.R.; Codo, C.R.B.; Sanches, V.L.; Guiraldelo, T.C.; Da Silva, F.F.; Ribessi, R.L.; Marba, S.T.M.; Cadore, S. Trace mineral composition of human breast milk from Brazilian mothers. J. Trace Elem. Med. Biol. 2019, 54, 199–205. [Google Scholar] [CrossRef]
- Keikha, M.; Bahreynian, M.; Saleki, M.; Kelishadi, R. Macro- and micronutrients of human milk composition: Are they related to maternal diet? A comprehensive systematic review. Breastfeed. Med. 2017, 12, 517–527. [Google Scholar] [CrossRef]
- Tilden, C.D.; Oftedal, O.T. Milk composition reflects pattern of maternal care in prosimian primates. Am. J. Primatol. 1997, 41, 195–211. [Google Scholar] [CrossRef]
- Power, M.L.; Oftedal, O.T.; Tardif, S.D. Does the milk of callitrichid monkeys differ from that of larger anthropoids? Am. J. Primatol. 2002, 56, 117–127. [Google Scholar] [CrossRef]
- Knight, C.H.; Maltz, E.; Docherty, A.H. Milk yield and composition in mice: Effects of litter size and lactation number. Comp. Biochem. Physiol. Part A Physiol. 1986, 84, 127–133. [Google Scholar] [CrossRef]
- Riley, L.G.; Zubair, M.; Thomson, P.C.; Holt, M.; Xavier, S.P.; Wynn, P.C.; Sheehy, P.A. Lactational performance of Quackenbush Swiss line 5 mice. J. Anim. Sci. 2006, 84, 2118–2125. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ragueneau, S. Early development in mice. IV: Quantity and gross composition of milk in five inbred strains. Physiol. Behav. 1987, 40, 431–435. [Google Scholar] [CrossRef]
- Nicholas, K.R.; Hartmann, P.E. Milk secretion in the rat: Progressive changes in milk composition during lactation and weaning and the effect of diet. Comp. Biochem. Physiol. Part A Physiol. 1991, 98, 535–542. [Google Scholar] [CrossRef]
- Akers, R.M.; Denbow, D.M. Lactation and animal agriculture. In Anatomy & Physiology of Domestic Animals; Balckwell Pub-lishing: Hoboken, NJ, USA, 2008; pp. 475–500. [Google Scholar]
- European Parliament; European Council. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes Text with EEA Relevance. 47; European Parliament: Brussels, Belgium; European Council: Brussels, Belgium, 2010. [Google Scholar]
- Hasiwa, N. Critical evaluation of the use of dogs in biomedical research and testing in Europe. ALTEX 2011, 28, 326–340. [Google Scholar] [CrossRef] [PubMed]


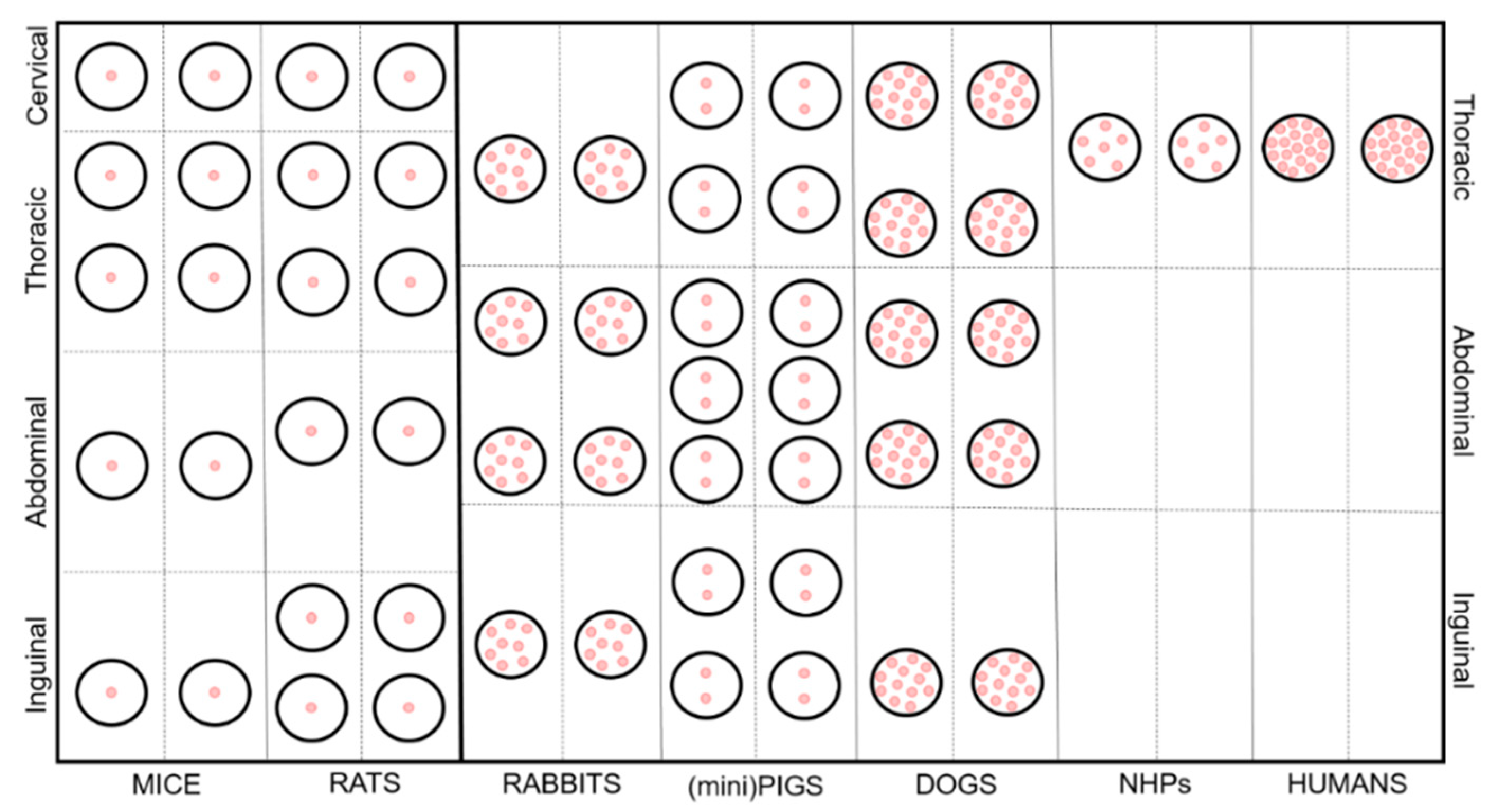{kind=link}
| Species | Number of Glands | Number of Teats/Nipples | Number of Canals per Gland | Location | References |
|---|---|---|---|---|---|
| Rats | 12 | 12 | 1 | cervical (1 pr) thoracic/pectoral (2 pr) abdominal (1 pr) inguinal (2 pr) | [11,12] |
| Mice | 10 | 10 | 1 | cervical (1 pr) thoracic/pectoral (2 pr) abdominal (1 pr) inguinal (1 pr) | [11,13,14,15,16] |
| Rabbits | 8–10 | 8–10 | 6–10 | thoracic (1 pr) abdominal (2 pr) inguinal (1 pr) | [17,18] |
| Dogs | 8–10 | 8–10 | 7–16 | thoracic (2 pr) abdominal/inguinal (2–3 pr) | [16,19] |
| (Mini)Pigs | 10–18 | 12–18 | 1–3 | thoracic (2 pr) abdominal (3 pr) inguinal (2 pr) | [20,21] |
| Non-Human Primates (NHPs) | 2 | 2 | 5–7 | pectoral (1 pr) | [22] |
| Humans | 2 | 2 | 10–25 | pectoral (1 pr) | [11,23] |
| Species | Duration of Colostrum Production | Yield of Colostrum | Duration of Milk Production | Yield of Milk | References |
|---|---|---|---|---|---|
| Rats | / | / | ~21 days | / | [47] |
| Mice | / | / | ~18 days | 0.1–0.5 mL | [48] |
| Rabbits | / | / | 4–5 weeks | 100–200 g/day | [49,50] |
| Dogs | 48 h | 270 mL/day | ~8–10 weeks | ~1000 mL/day | [51,52,53,54] |
| (mini)Pigs | 24 h | ~3.75 kg/day * | ~8 weeks | 4500–5700 g/day * | [55,56] |
| NHPs | / | / | ~12 months | / | [22,57] |
| Humans | 96 h | ~500 mL/day | ~6 months | ~800 mL/day | [39,58] |
| Species | Dry Matter % | Protein % | Casein % | Whey Protein % | Fat % | Lactose % | Fe µg/mL | Cu µg/mL | Zn µg/mL | Mn µg/mL | Mg µg/mL | Ca µg/mL | P µg/mL | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rats | / | 8.6–9.1 | / | / | 13.6–15.7 | 2.3–2.6 | 8.1–9.2 | 8.6–9.8 | 13.3–14.2 | 0.3–0.4 | 168–180 | 755–829 | / | [71] |
| Mice | / | / | / | / | / | / | / | / | / | / | / | / | / | / |
| Rabbits | 31.4–33.7 | 13.5–15.9 | / | / | 13.7–20.4 | 1.6–2.1 | / | / | / | / | / | / | / | [72] |
| Dogs | / | 12.4–16.2 | 60.7 | 39.3 | 13.2 | 1.7 | 3.7 | 1.3 | 5 | / | 128 | 1363 | 935 | [73] |
| (mini)Pigs | 20.1–26.7 | 7.7–16.6 | 1.5–3.4 | 7.9–14.8 | 6.4–8 | 2.8–3.9 | / | / | / | / | 100 | 800 | 1000 | [56,74] |
| NHPs | / | 2.2–2.7 | / | / | 4.3–6.3 | 7.7–7.9 | 0.9–2.6 | 2–4.1 | 3.5–6.8 | / | 37.5–61.7 | 324–347 | / | [75] |
| Humans | 11.92 | 2.6 | 0.4 | 1.18 | 3 | 5.8 | 1.1 | 0.4 | 4.8 | 0.01 | 32 | 293 | 159 | [76,77,78] |
| Species | Dry Matter % | Protein % | Casein % | Whey Protein % | Fat % | Lactose % | Fe µg/mL | Cu µg/mL | Zn µg/mL | Mn µg/mL | Mg µg/mL | Ca µg/mL | P µg/mL | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rats | 27.9–32.8 | 8.9–9.7 | 6.4–8 | 0.9–2.5 | 14–15.9 | 1.1–4.1 | 4–7 | 1.7–7 | 9-55 | / | 158–192 | 2849– 3206 | 1600– 2720 | [79,80] |
| Mice | 36.3–39.4 | 10.1–12.7 | / | / | 19.3–22.9 | 2.4–2.8 | / | / | / | / | / | / | / | [48] |
| Rabbits | 31.2 | 10.3 | / | / | 15.2 | 1.8 | 0.003 § | 0.002 § | 0.02–0.03 § | 0.0001 § | 0.35–0.45 § | 2.71–5.36 § | 2.44–3.28 § | [72,81] |
| Dogs | 22.7–26 | 4.3–9.8 | 65.8–75.4 * | 26.4–34.2 * | 2.4–13.4 | 29.3–40.2 | 1.8–13.1 | 0.9–2 | 4.1–9.6 | 0.1-0.2 | 55.8–104.3 | 1366–2440 | 914–1401 | [52,73,81,82,83] |
| (mini)Pigs | 18.8–22.7 | 5–7.5 | 2.7–3.6 | 2.4–5.4 | 7–10.1 | 4.3–5.6 | / | / | / | / | 105 | 2000 | 1420 | [56,74] |
| NHPs | 12.2–14 | 1.3–2.3 | 45 * | 55 * | 3.3–6.2 | 4.8–9.1 | / | / | / | / | 34 | 380 | 152 | [81,84,85,86,87] |
| Humans | 12.6 | 1.2 | 0.3 | 0.7 | 4.1 | 7 | 0.5–1.8 | 0.2–5.2 | 0.7–3.8 | 0.01-0.03 | 25–33 | 230–310 | 130–190 | [88,89,90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventrella, D.; Ashkenazi, N.; Elmi, A.; Allegaert, K.; Aniballi, C.; DeLise, A.; Devine, P.J.; Smits, A.; Steiner, L.; Forni, M.; et al. Animal Models for In Vivo Lactation Studies: Anatomy, Physiology and Milk Compositions in the Most Used Non-Clinical Species: A Contribution from the ConcePTION Project. Animals 2021, 11, 714. https://doi.org/10.3390/ani11030714
Ventrella D, Ashkenazi N, Elmi A, Allegaert K, Aniballi C, DeLise A, Devine PJ, Smits A, Steiner L, Forni M, et al. Animal Models for In Vivo Lactation Studies: Anatomy, Physiology and Milk Compositions in the Most Used Non-Clinical Species: A Contribution from the ConcePTION Project. Animals. 2021; 11(3):714. https://doi.org/10.3390/ani11030714
Chicago/Turabian StyleVentrella, Domenico, Nurit Ashkenazi, Alberto Elmi, Karel Allegaert, Camilla Aniballi, Anthony DeLise, Patrick John Devine, Anne Smits, Lilach Steiner, Monica Forni, and et al. 2021. "Animal Models for In Vivo Lactation Studies: Anatomy, Physiology and Milk Compositions in the Most Used Non-Clinical Species: A Contribution from the ConcePTION Project" Animals 11, no. 3: 714. https://doi.org/10.3390/ani11030714
APA StyleVentrella, D., Ashkenazi, N., Elmi, A., Allegaert, K., Aniballi, C., DeLise, A., Devine, P. J., Smits, A., Steiner, L., Forni, M., Bouisset-Leonard, M., & Bacci, M. L. (2021). Animal Models for In Vivo Lactation Studies: Anatomy, Physiology and Milk Compositions in the Most Used Non-Clinical Species: A Contribution from the ConcePTION Project. Animals, 11(3), 714. https://doi.org/10.3390/ani11030714