
Differences in a Cage Escape Behaviour between Two Migrating Warblers of Different Stop-Over Strategy

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Cage Escape Experiment
2.3. Statistical Analyses
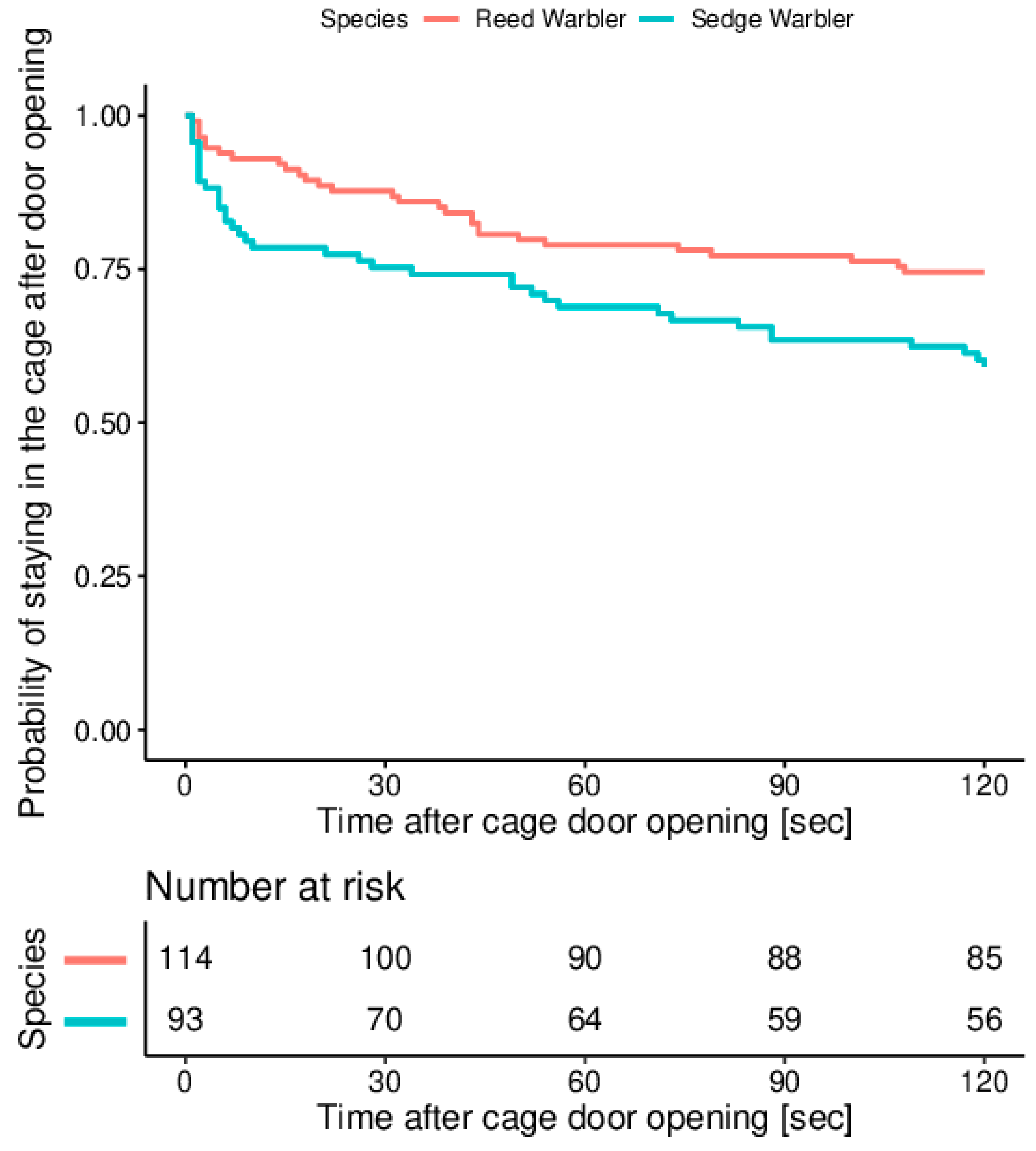
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shettleworth, S.J. Cognition, Evolution, and Behavior; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Zentall, T.; Wasserman, E. The Oxford Handbook of Comparative Cognition; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Iyengar, S.; Parishar, P.; Mohapatra, A.N. Avian cognition and consciousness-from the perspective of neuroscience and behaviour. In Self, Culture and Consciousness: Interdisciplinary Convergences on Knowing and Being; Springer: Singapore, 2018; pp. 23–50. [Google Scholar] [CrossRef]
- Gwinner, E. Circannual Rhythms in the Control of Avian Migrations. Adv. Study Behav. 1986, 16, 191–228. [Google Scholar]
- Leisler, B.; Winkler, H. Morphological Consequences of Migration in Passerines. In Avian Migration; Springer: Berlin/Heidelberg, Germany, 2003; pp. 175–186. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C. Avian movements in a modern world: Cognitive challenges. Anim. Cogn. 2017, 20, 77–86. [Google Scholar] [CrossRef]
- Cramp, S. The Complete Birds of the Western Palearctic on CD-ROM; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Bibby, C.J.; Green, R.E. Autumn Migration Strategies of Reed and Sedge Warblers. Ornis Scand. 1981, 12, 1–12. [Google Scholar] [CrossRef]
- Bibby, C.J.; Green, R.E. Food and fattening of migrating warblers in some french marshlands. Ringing Migr. 1983, 4, 175–184. [Google Scholar] [CrossRef]
- Chernetsov, N.S.; Titov, N. Movement patterns of European Reed Warblers Acrocephalus scirpaceus and Sedge Warblers A. schoenobaenus before and during autumn migration. Ardea 2001, 89, 509–515. [Google Scholar]
- Chernetsov, N.; Manukyan, A. Foraging strategy of the Sedge Warbler (Acrocephalus schoenobaenus) on migration. Vogelwarte 2000, 40, 189–197. [Google Scholar]
- Kerbiriou, C.; Bargain, B.; Le Viol, I.; Pavoine, S. Diet and fuelling of the globally threatened aquatic warbler at autumn migration stopover as compared with two congeners. Anim. Conserv. 2011, 14, 261–270. [Google Scholar] [CrossRef]
- Koskimies, P.; Saurola, P. Autumn migration strategies of the Sedge Warbler Acrocephalus schoenobaenus in Finland: A preliminary report. Ornis Fenn. 1985, 62, 145–152. [Google Scholar]
- Schaub, M.; Jenni, L. Body mass of six long-distance migrant passerine species along the autumn migration route. J. Ornithol. 2000, 141, 441–460. [Google Scholar] [CrossRef]
- Schaub, M.; Jenni, L. Fuel deposition of three passerine bird species along the migration route. Oecologia 2000, 122, 306–317. [Google Scholar] [CrossRef]
- Stȩpniewska, K.; Ozarowska, A.; Busse, P.; Zehtindjiev, P.; Ilieva, M.; Hnatyna, O.; Meissner, W. Fuelling strategies differ among juvenile Sedge and Reed Warblers along the eastern European flyway during autumn migration. Ornis Fenn. 2018, 95, 103–114. [Google Scholar]
- BirdLife International and Handbook of the Birds of the World. Bird Species Distribution Maps of the World. Version 7.0. 2018. Available online: http://datazone.birdlife.org (accessed on 10 September 2020).
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An Ecoregion-Based Approach to Protecting Half the Terrestrial Realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef]
- Formella, M.; Busse, P. Directional preferences of the Reed Warbler (Acrocephalus scirpaceus) and the Sedge Warbler (A. schoenobaenus) on autumn migration at Lake Drużno (N Poland). Ring 2002, 24, 15–29. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C. Learning and cognition in resident and migrant birds. In Behavioral and Chemical Ecology; Zhang, W., Liu, H., Eds.; Nova Science Publishers: New York, NY, USA, 2010; pp. 155–172. [Google Scholar]
- Mettke-Hofmann, C. Cognitive ecology: Ecological factors, life-styles, and cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 5, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.L.K.; Nilsson, J.Å.; Alerstam, T.; Bäckman, J. Migratory and resident blue tits Cyanistes caeruleus differ in their reaction to a novel object. Naturwissenschaften 2010, 97, 981–985. [Google Scholar] [CrossRef]
- Jakubas, D.; Wojczulanis-Jakubas, K. Sex-and age-related differences in the timing and body condition of migrating Reed Warblers Acrocephalus scirpaceus and Sedge Warblers Acrocephalus schoenobaenus. Naturwissenschaften 2010, 97, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Nitecki, C.; Jakubas, D.; Typiak, J. Awifauna jeziora Druzno. Nitecki, C. Jezioro Druzno—Monografia Przyrodnicza; Wydawnictwo Mantis: Olsztyn, Poland, 2013; pp. 113–190. [Google Scholar]
- Chernetsov, N. Habitat distribution during the post-breeding and post-fledging period in the Reed Warbler Acrocephalus scirpaceus and Sedge Warbler A. schoenobaenus depends on food abundance. Ornis Svecica 1998, 8, 77–82. [Google Scholar]
- Kozłowska, A.; Stępniewska, K.; Stępniewski, K.; Busse, P. Dynamics of Autumn Migration of the Acrocephalus Warblers through the Polish Baltic Coast. Ring 2010, 31, 15–43. [Google Scholar] [CrossRef]
- Wojczulanis-Jakubas, K.; Wietrzykowski, J.; Jakubas, D. Response of Reed Warbler and Sedge Warbler to acoustic playback in relation to age, sex, and body condition. J. Ornithol. 2016, 157, 137–143. [Google Scholar] [CrossRef]
- Ellrich, H.; Salewski, V.; Fiedler, W. Morphological sexing of passerines: Not valid over larger geographical scales. J. Ornithol. 2010, 151, 449–458. [Google Scholar] [CrossRef]
- Wojczulanis-Jakubas, K.; Jakubas, D. Predicting the sex of the Sedge Warbler (Acrocephalus schoenobaenus) by discriminant analysis. Ornis Fenn. 2011, 88, 90–97. [Google Scholar]
- Kassambra, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves using ggplot2. R package version 0.4.8. 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Mettke-Hofmann, C.; Lorentzen, S.; Schlicht, E.; Schneider, J.; Werner, F. Spatial Neophilia and Spatial Neophobia in Resident and Migratory Warblers (Sylvia). Ethology 2009, 115, 482–492. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C.; Gwinner, E. Differential assessment of environmental information in a migratory and a nonmigratory passerine. Anim. Behav. 2004, 68, 1079–1086. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C.; Wink, M.; Braun, M.; Winkler, H. Residency and a Broad Feeding Spectrum are Related to Extensive Spatial Exploration in Parrots. Behav. Ecol. 2012, 23, 1365–1371. [Google Scholar] [CrossRef]
- Godfrey-Smith, P. Environmental complexity and the evolution of cognition. In The Evolution of Intelligence; Sternberg, R., Kaufman, J., Eds.; Lawrence Erlbaum: Mahwah, NJ, USA, 2002; pp. 233–249. [Google Scholar] [CrossRef]
- Bernays, E.A.; Funk, D.J. Specialists make faster decisions than generalists: Experiments with aphids. Proc. R. Soc. B Biol. Sci. 1999, 266, 151–156. [Google Scholar] [CrossRef]
- Bernays, E.A. The value of being a resource specialist: Behavioral support for a neural hypothesis. Am. Nat. 1998, 151, 451–464. [Google Scholar] [CrossRef]
- Greenberg, R. The role of neophobia and neophilia in the development of innovative behaviour of birds. In Animal Innovation; Reader, S.M., Laland, K.N., Eds.; Oxford University Press: Oxford, UK, 2003; pp. 175–196. [Google Scholar]
- Feenders, G.; Klaus, K.; Bateson, M. Fear and exploration in European starlings (Sturnus vulgaris): A comparison of hand-reared and wild-caught birds. PLoS ONE 2011, 6, e19074. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Both, C.; Drent, P.J.; Van Oers, K.; Van Noordwijk, A.J. Repeatability and heritability of exploratory behaviour in great tits from the wild. Anim. Behav. 2002, 64, 929–938. [Google Scholar] [CrossRef]
- Saino, N.; Szép, T.; Ambrosini, R.; Romano, M.; Møller, A.P. Ecological conditions during winter affect sexual selection and breeding in a migratory bird. Proc. R. Soc. B Biol. Sci. 2004, 271, 681–686. [Google Scholar] [CrossRef]
- Norris, D.R.; Taylor, C.M. Predicting the consequences of carry-over effects for migratory populations. Biol. Lett. 2006, 2, 148–151. [Google Scholar] [CrossRef]




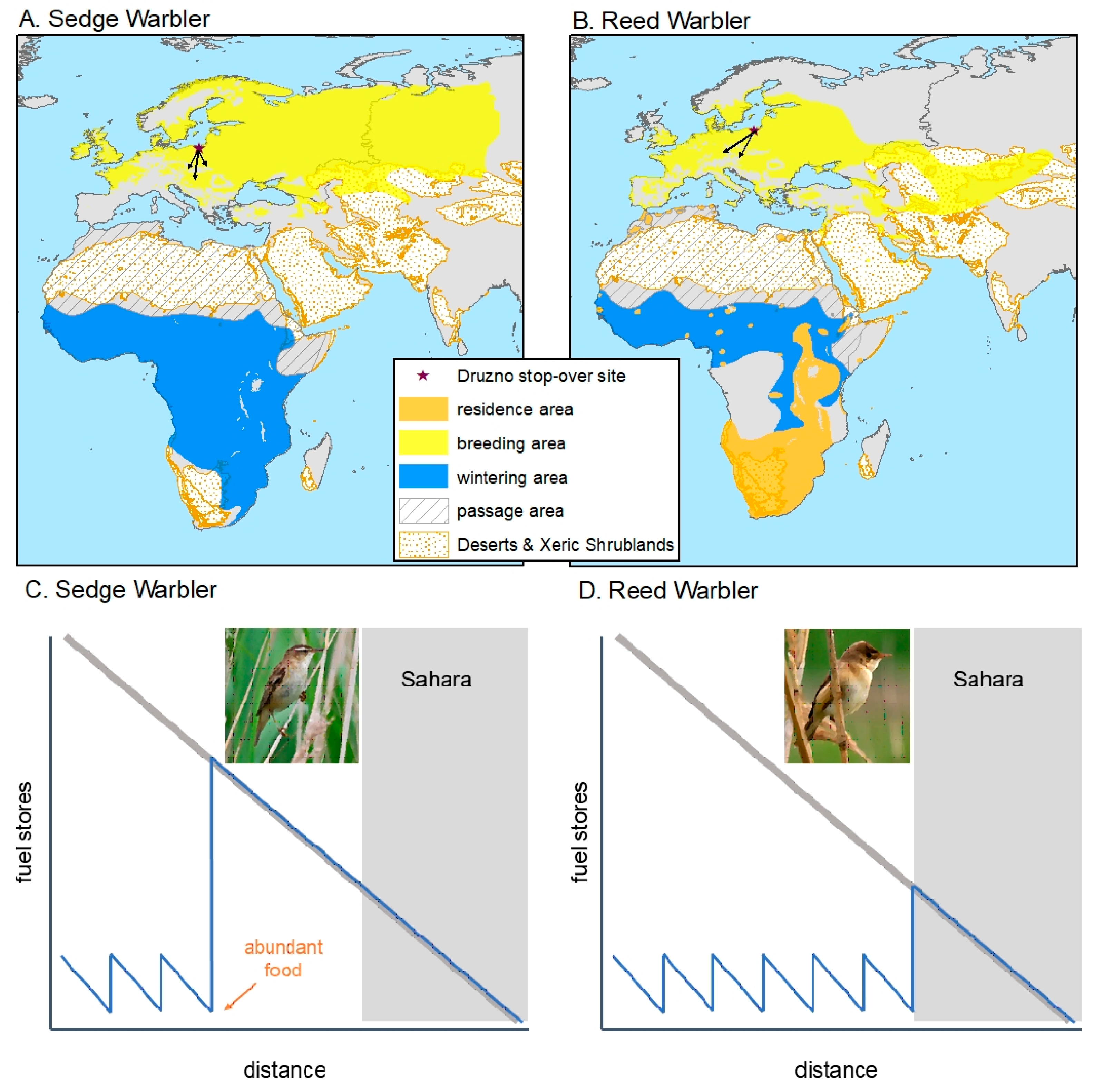{kind=link}
{kind=link}
{kind=link}
| Species | Escaped | Non-Escaped | Total |
|---|---|---|---|
| Sedge Warbler | 38 (41%) | 55 (59%) | 93 (100%) |
| Reed Warbler | 29 (25%) | 85 (75%) | 114 (100%) |
| Total | 67 | 140 | 207 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakubas, D.; Wojczulanis-Jakubas, K.; Powers, A.; Frazier, T.; Bottomley, M.; Kraszpulski, M. Differences in a Cage Escape Behaviour between Two Migrating Warblers of Different Stop-Over Strategy. Animals 2021, 11, 639. https://doi.org/10.3390/ani11030639
Jakubas D, Wojczulanis-Jakubas K, Powers A, Frazier T, Bottomley M, Kraszpulski M. Differences in a Cage Escape Behaviour between Two Migrating Warblers of Different Stop-Over Strategy. Animals. 2021; 11(3):639. https://doi.org/10.3390/ani11030639
Chicago/Turabian StyleJakubas, Dariusz, Katarzyna Wojczulanis-Jakubas, Alexis Powers, Troy Frazier, Michael Bottomley, and Michał Kraszpulski. 2021. "Differences in a Cage Escape Behaviour between Two Migrating Warblers of Different Stop-Over Strategy" Animals 11, no. 3: 639. https://doi.org/10.3390/ani11030639
APA StyleJakubas, D., Wojczulanis-Jakubas, K., Powers, A., Frazier, T., Bottomley, M., & Kraszpulski, M. (2021). Differences in a Cage Escape Behaviour between Two Migrating Warblers of Different Stop-Over Strategy. Animals, 11(3), 639. https://doi.org/10.3390/ani11030639