
The Effect of Mother Goat Presence during Rearing on Kids’ Response to Isolation and to an Arena Test



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing, and Management
2.2. Experimental Procedure
2.2.1. Isolation Test at Two Weeks of Age
2.2.2. Isolation and Novel Arena Test at Age Two Months
2.2.3. Behavioral Observations
2.2.4. Heart Rate
2.2.5. Saliva Cortisol
2.3. Statistics
3. Results
3.1. Isolation Test
3.1.1. Heart Rate and Vocalization during Isolation—Comparisons between Treatments at Two Weeks
3.1.2. Heart Rate and Vocalization during Isolation—Comparisons between Treatments at Two Months
3.1.3. Heart Rate and Vocalization during Isolation—Comparisons between Ages
3.1.4. Saliva Cortisol during Isolation—Comparisons between Treatments at Two Weeks, and Two Months
3.2. Novel Arena Test
3.2.1. Heart Rate during the Arena Test—Comparisons between Treatments
3.2.2. Behavior during the Arena Test—Comparisons between Treatments
3.2.3. Heart Rate and Behavior—Reactions to Novel Object
3.2.4. Saliva Cortisol during the Arena Test—Comparisons between and within Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boogaard, B.K.; Oosting, S.J.; Bock, B.B. Defining Sustainability as a Socio-Cultural Concept: Citizen Panels Visiting Dairy Farms in the Netherlands. Livest. Sci. 2008, 117, 24–33. [Google Scholar] [CrossRef]
- Boogaard, B.K.; Oosting, S.J.; Bock, B.B.; Wiskerke, J.S.C. The Sociocultural Sustainability of Livestock Farming: An Inquiry into Social Perceptions of Dairy Farming. Anim. Int. J. Anim. Biosci. 2011, 5, 1458–1466. [Google Scholar] [CrossRef]
- Boogaard, B.K.; Bock, B.B.; Oosting, S.J.; Wiskerke, J.S.C.; van der Zijpp, A.J. Social Acceptance of Dairy Farming: The Ambivalence Between the Two Faces of Modernity. J. Agric. Environ. Ethics 2011, 24, 259–282. [Google Scholar] [CrossRef]
- Ellis, K.; Billington, K.; McNeil, B.; McKeegan, D. Public Opinion on UK Milk Marketing and Dairy Cow Welfare. Anim. Welf. 2009, 18, 267–282. [Google Scholar]
- Ventura, B.A.; von Keyserlingk, M.A.G.; Schuppli, C.A.; Weary, D.M. Views on Contentious Practices in Dairy Farming: The Case of Early Cow-Calf Separation. J. Dairy Sci. 2013, 96, 6105–6116. [Google Scholar] [CrossRef]
- Ventura, B.A.; von Keyserlingk, M.A.G.; Wittman, H.; Weary, D.M. What Difference Does a Visit Make? Changes in Animal Welfare Perceptions after Interested Citizens Tour a Dairy Farm. PLoS ONE 2016, 11, e0154733. [Google Scholar] [CrossRef]
- Johnsen, J.F.; Zipp, K.A.; Kälber, T.; de Passillé, A.M.; Knierim, U.; Barth, K.; Mejdell, C.M. Is Rearing Calves with the Dam a Feasible Option for Dairy Farms?—Current and Future Research. Appl. Anim. Behav. Sci. 2016, 181, 1–11. [Google Scholar] [CrossRef]
- Agenäs, S. Editorial: We Need to Bring the Calves Back to the Dairy Cows. J. Dairy Res. 2017, 84, 239. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Busch, G.; Weary, D.M.; Spiller, A.; Keyserlingk, M.A.G. von American and German Attitudes towards Cow-Calf Separation on Dairy Farms. PLoS ONE 2017, 12, e0174013. [Google Scholar] [CrossRef] [PubMed]
- Mikuš, T.; Marzel, R.; Mikuš, O. Early Weaning: New Insights on an Ever-Persistent Problem in the Dairy Industry. J. Dairy Res. 2020, 87, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Placzek, M.; Christoph-Schulz, I.; Barth, K. Public Attitude towards Cow-Calf Separation and Other Common Practices of Calf Rearing in Dairy Farming—A Review. Org. Agric. 2020. [Google Scholar] [CrossRef]
- Boivin, X.; Braastad, B.O. Effects of Handling during Temporary Isolation after Early Weaning on Goat Kids’ Later Response to Humans. Appl. Anim. Behav. Sci. 1996, 48, 61–71. [Google Scholar] [CrossRef]
- Bergamasco, L.; Macchi, E.; Facello, C.; Badino, P.; Odore, R.; Pagliasso, S.; Bellino, C.; Osella, M.C.; Re, G. Effects of Brief Maternal Separation in Kids on Neurohormonal and Electroencephalographic Parameters. Appl. Anim. Behav. Sci. 2005, 93, 39–52. [Google Scholar] [CrossRef]
- Sevi, A.; Casamassima, D.; Pulina, G.; Pazzona, A. Factors of Welfare Reduction in Dairy Sheep and Goats. Ital. J. Anim. Sci. 2009, 8, 81–101. [Google Scholar] [CrossRef]
- Miranda-de la Lama, G.; Mattiello, S. The Importance of Social Behaviour for Goat Welfare in Livestock Farming. Small Rumin. Res. 2010, 90, 1–10. [Google Scholar] [CrossRef]
- von Winblad Walter, L.; Lidfors, L.; Madej, A.; Dahlborn, K.; Hydbring-Sandberg, E. Cardiovascular, Endocrine and Behavioural Responses to Suckling and Permanent Separation in Goats. Acta Vet. Scand. 2010, 52, 51. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, H.; Delgadillo, J.; Flores, J.; Rodríguez, A.; Serafín, N.; Kann, G.; Marnet, P.; Poindron, P. Effects of Restricted Nursing on Milk Production and Collection, Kid Growth and Plasma Prolactin and Growth Hormone Concentrations in Dairy Goats. Anim. Int. J. Anim. Biosci. 2007, 1, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Högberg, M. Keeping Goats and Kids Together. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2 May 2016. [Google Scholar]
- Delgado-Pertíñez, M.; Guzmán-Guerrero, J.L.; Mena, Y.; Castel, J.M.; González-Redondo, P.; Caravaca, F.P. Influence of Kid Rearing Systems on Milk Yield, Kid Growth and Cost of Florida Dairy Goats. Small Rumin. Res. 2009, 81, 105–111. [Google Scholar] [CrossRef]
- Delgado-Pertíñez, M.; Guzmán-Guerrero, J.L.; Caravaca, F.P.; Castel, J.M.; Ruiz, F.A.; González-Redondo, P.; Alcalde, M.J. Effect of Artificial vs. Natural Rearing on Milk Yield, Kid Growth and Cost in Payoya Autochthonous Dairy Goats. Small Rumin. Res. 2009, 84, 108–115. [Google Scholar] [CrossRef]
- Godden, S.M.; Fetrow, J.P.; Feirtag, J.M.; Green, L.R.; Wells, S.J. Economic Analysis of Feeding Pasteurized Nonsaleable Milk versus Conventional Milk Replacer to Dairy Calves. J. Am. Vet. Med. Assoc. 2005, 226, 1547–1554. [Google Scholar] [CrossRef] [PubMed]
- Wagenaar, J.P.T.M.; Langhout, J. Practical Implications of Increasing ‘Natural Living’ through Suckling Systems in Organic Dairy Calf Rearing. NJAS Wagening. J. Life Sci. 2007, 54, 375–386. [Google Scholar] [CrossRef]
- Meagher, R.K.; Beaver, A.; Weary, D.M.; von Keyserlingk, M.A.G. Invited Review: A Systematic Review of the Effects of Prolonged Cow–Calf Contact on Behavior, Welfare, and Productivity. J. Dairy Sci. 2019, 102, 5765–5783. [Google Scholar] [CrossRef]
- Högberg, M.; Dahlborn, K.; Hydbring-Sandberg, E.; Hartmann, E.; Andrén, A. Milk Processing Quality of Suckled/Milked Goats: Effects of Milk Accumulation Interval and Milking Regime. J. Dairy Res. 2016, 83, 173–179. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.H. Leavers and Stayers: Maternal Post-Partum Strategies in Feral Goats. Appl. Anim. Behav. Sci. 1984, 12, 233–243. [Google Scholar] [CrossRef]
- Lickliter, R.E. Hiding Behavior in Domestic Goat Kids. Appl. Anim. Behav. Sci. 1984, 12, 245–251. [Google Scholar] [CrossRef]
- Shackleton, D.; Shank, C. A Review of the Social Behavior of Feral and Wild Sheep and Goats. J. Anim. Sci. 1984, 58, 500–509. [Google Scholar] [CrossRef]
- Andersen, I.L.; Roussel, S.; Ropstad, E.; Braastad, B.O.; Steinheim, G.; Janczak, A.M.; Jørgensen, G.m.; Bøe, K.E. Social Instability Increases Aggression in Groups of Dairy Goats, but with Minor Consequences for the Goats’ Growth, Kid Production and Development. Appl. Anim. Behav. Sci. 2008, 114, 132–148. [Google Scholar] [CrossRef]
- O’Brien, P.H. Feral Goat Social Organization: A Review and Comparative Analysis. Appl. Anim. Behav. Sci. 1988, 21, 209–221. [Google Scholar] [CrossRef]
- Lickliter, R.E. Activity Patterns and Companion Preferences of Domestic Goat Kids. Appl. Anim. Behav. Sci. 1987, 19, 137–145. [Google Scholar] [CrossRef]
- Allan, C.J.; Holst, P.J.; Hinch, G.N. Behaviour of Parturient Australian Bush Goats. I. Doe Behaviour and Kid Vigour. Appl. Anim. Behav. Sci. 1991, 32, 55–64. [Google Scholar] [CrossRef]
- Bungo, T.; Shimojo, M.; Nakano, Y.; Okano, K.; Masuda, Y.; Goto, I. Relationship between Nursing and Suckling Behaviour in Tokara Native Goats. Appl. Anim. Behav. Sci. 1998, 59, 357–362. [Google Scholar] [CrossRef]
- Saunders, F.; McElligott, A.; Safi, K.; Hayden, T. Mating Tactics of Male Feral Goats (Capra Hircus): Risks and Benefits. Acta Ethologica 2013, 8, 103–110. [Google Scholar] [CrossRef]
- Briefer, E.; McElligott, A.G. Mutual Mother–Offspring Vocal Recognition in an Ungulate Hider Species (Capra Hircus). Anim. Cogn. 2011, 14, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Briefer, E.F.; Padilla de la Torre, M.; McElligott, A.G. Mother Goats Do Not Forget Their Kids’ Calls. Proc. R. Soc. B Biol. Sci. 2012, 279, 3749–3755. [Google Scholar] [CrossRef]
- Poindron, P.; Gilling, G.; Hernandez, H.; Serafin, N.; Terrazas, A. Early Recognition of Newborn Goat Kids by Their Mother: I. Nonolfactory Discrimination. Dev. Psychobiol. 2003, 43, 82–89. [Google Scholar] [CrossRef]
- Poindron, P.; Lévy, F.; Keller, M. Maternal Responsiveness and Maternal Selectivity in Domestic Sheep and Goats: The Two Facets of Maternal Attachment. Dev. Psychobiol. 2007, 49, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Poindron, P.; Terrazas, A.; Montes, M.D.L.L.N.; Serafín, N.; Hernández, H. Sensory and Physiological Determinants of Maternal Behavior in the Goat (Capra Hircus). Horm. Behav. 2007, 52, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Carbonaro, D.A.; Friend, T.H.; Dellmeier, G.R.; Nuti, L.C. Behavioral and Physiological Responses of Dairy Goats to Isolation. Physiol. Behav. 1992, 51, 297–301. [Google Scholar] [CrossRef]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.-C.; Canali, E.; Jones, R.B. A Critical Review of Fear Tests Used on Cattle, Pigs, Sheep, Poultry and Horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef]
- Siebert, K.; Langbein, J.; Schön, P.-C.; Tuchscherer, A.; Puppe, B. Degree of Social Isolation Affects Behavioural and Vocal Response Patterns in Dwarf Goats (Capra Hircus). Appl. Anim. Behav. Sci. 2011, 131. [Google Scholar] [CrossRef]
- Högberg, M.; Winblad von Walter, L.; Hydbring-Sandberg, E.; Forkman, B.; Dahlborn, K. Growth Rate and Behaviour in Early, Daily or Not Separated Kids and the Corresponding Milk Production of Their Goats. Small Rumin. Res. 2010, 90, 1–10. [Google Scholar]
- Friard, O.; Gamba, M. BORIS: A Free, Versatile Open-Source Event-Logging Software for Video/Audio Coding and Live Observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Manteuffel, G.; Puppe, B.; Schön, P.C. Vocalization of Farm Animals as a Measure of Welfare. Appl. Anim. Behav. Sci. 2004, 88, 163–182. [Google Scholar] [CrossRef]
- O’Bryan, L.R.; Abaid, N.; Nakayama, S.; Dey, T.; King, A.J.; Cowlishaw, G.; Rubenstein, D.I.; Garnier, S. Contact Calls Facilitate Group Contraction in Free-Ranging Goats (Capra Aegagrus Hircus). Front. Ecol. Evol. 2019, 7, 73. [Google Scholar] [CrossRef]
- Alvarez, L.; Nava, R.A.; Ramírez, A.; Ramírez, E.; Gutiérrez, J. Physiological and Behavioural Alterations in Disbudded Goat Kids with and without Local Anaesthesia. Appl. Anim. Behav. Sci. 2009, 117, 190–196. [Google Scholar] [CrossRef]
- Chojnacki, R.; Vas, J.; Andersen, I. The Effects of Prenatal Stocking Densities on the Fear Responses and Sociality of Goat ( Capra Hircus) Kids. PLoS ONE 2014, 9, e94253. [Google Scholar] [CrossRef]
- Price, E.O.; Thos, J. Behavioral Responses to Short-Term Social Isolation in Sheep and Goats. Appl. Anim. Ethol. 1980, 6, 331–339. [Google Scholar] [CrossRef]
- Baciadonna, L.; Briefer, E.F.; Favaro, L.; McElligott, A.G. Goats Distinguish between Positive and Negative Emotion-Linked Vocalisations. Front. Zool. 2019, 16, 25. [Google Scholar] [CrossRef]
- Olsson, K.; Hydbring-Sandberg, E. Exposure to a Dog Elicits Different Cardiovascular and Behavioral Effects in Pregnant and Lactating Goats. Acta Vet. Scand. 2011, 53, 60. [Google Scholar] [CrossRef]
- Théoret-Gosselin, R.; Hamel, S.; Côté, S.D. The Role of Maternal Behavior and Offspring Development in the Survival of Mountain Goat Kids. Oecologia 2015, 178, 175–186. [Google Scholar] [CrossRef]
- Napolitano, F.; De Rosa, G.; Sevi, A. Welfare Implications of Artificial Rearing and Early Weaning in Sheep. Appl. Anim. Behav. Sci. 2008, 110, 58–72. [Google Scholar] [CrossRef]
- Boissy, A.; Bouissou, M.-F. Assessment of Individual Differences in Behavioural Reactions of Heifers Exposed to Various Fear-Eliciting Situations. Appl. Anim. Behav. Sci. 1995, 46, 17–31. [Google Scholar] [CrossRef]
- Müller, R.; Schrader, L. Behavioural Consistency during Social Separation and Personality in Dairy Cows. Behaviour 2005, 142, 1289–1306. [Google Scholar] [CrossRef]
- Roelofs, K. Freeze for Action: Neurobiological Mechanisms in Animal and Human Freezing. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160206. [Google Scholar] [CrossRef] [PubMed]
- Alm, P. Stuttering, Emotions, and Heart Rate during Anticipatory Anxiety: A Critical Review. J. Fluen. Disord. 2004, 29, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.; Barth, K.; Hillmann, E.; Palme, R.; Futschik, A.; Waiblinger, S. Mother Rearing of Dairy Calves: Reactions to Isolation and to Confrontation with an Unfamiliar Conspecific in a New Environment. Appl. Anim. Behav. Sci. 2013, 147, 43–54. [Google Scholar] [CrossRef]
- Bartolomé, E.; Azcona, F.; Cañete-Aranda, M.; Perdomo-González, D.I.; Ribes-Pons, J.; Terán, E.M. Testing Eye Temperature Assessed with Infrared Thermography to Evaluate Stress in Meat Goats Raised in a Semi-Intensive Farming System: A Pilot Study. Arch. Anim. Breed. 2019, 62, 199–204. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Shutt, D.A. Effects of Management Practices on Cortisol, β-Endorphin and Behaviour in Young Goats. Proc. Aust. Soc. Anim. Prod. 1990, 18, 224–227. [Google Scholar]
- Kannan, G.; Terrill, T.; Kouakou, B.; Gazal, O.; Gelaye, S.; Amoah, E.; Samaké, S. Transportation of Goats: Effects on Physiological Stress Responses and Live Weight Loss. J. Anim. Sci. 2000, 78, 1450–1457. [Google Scholar] [CrossRef]
- Kannan, G.; Terrill, T.; Kouakou, B.; Gelaye, S.; Amoah, E. Simulated Pre Slaughter Holding and Isolation Effect on Stress Responses and Livemass Shrinkage in Meat Goats. J. Anim. Sci. 2002, 80, 1771–1780. [Google Scholar] [CrossRef]
- Alvarez, L.; Gutiérrez, J. A First Description of the Physiological and Behavioural Responses to Disbudding in Goat Kids. Anim. Welf. 2010, 19, 55–59. [Google Scholar]
- Alvarez, L.; De Luna, J.B.; Gamboa, D.; Reyes, M.; Sánchez, A.; Terrazas, A.; Rojas, S.; Galindo, F. Cortisol and Pain-Related Behavior in Disbudded Goat Kids with and without Cornual Nerve Block. Physiol. Behav. 2015, 138, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Zipp, K.A.; Knierim, U. Physical Development, Ease of Integration into the Dairy Herd and Performance of Primiparous Dairy Cows Reared with Full Whole-Day, Half-Day or No Mother-Contact as Calves. J. Dairy Res. 2020, 87, 154–156. [Google Scholar] [CrossRef] [PubMed]
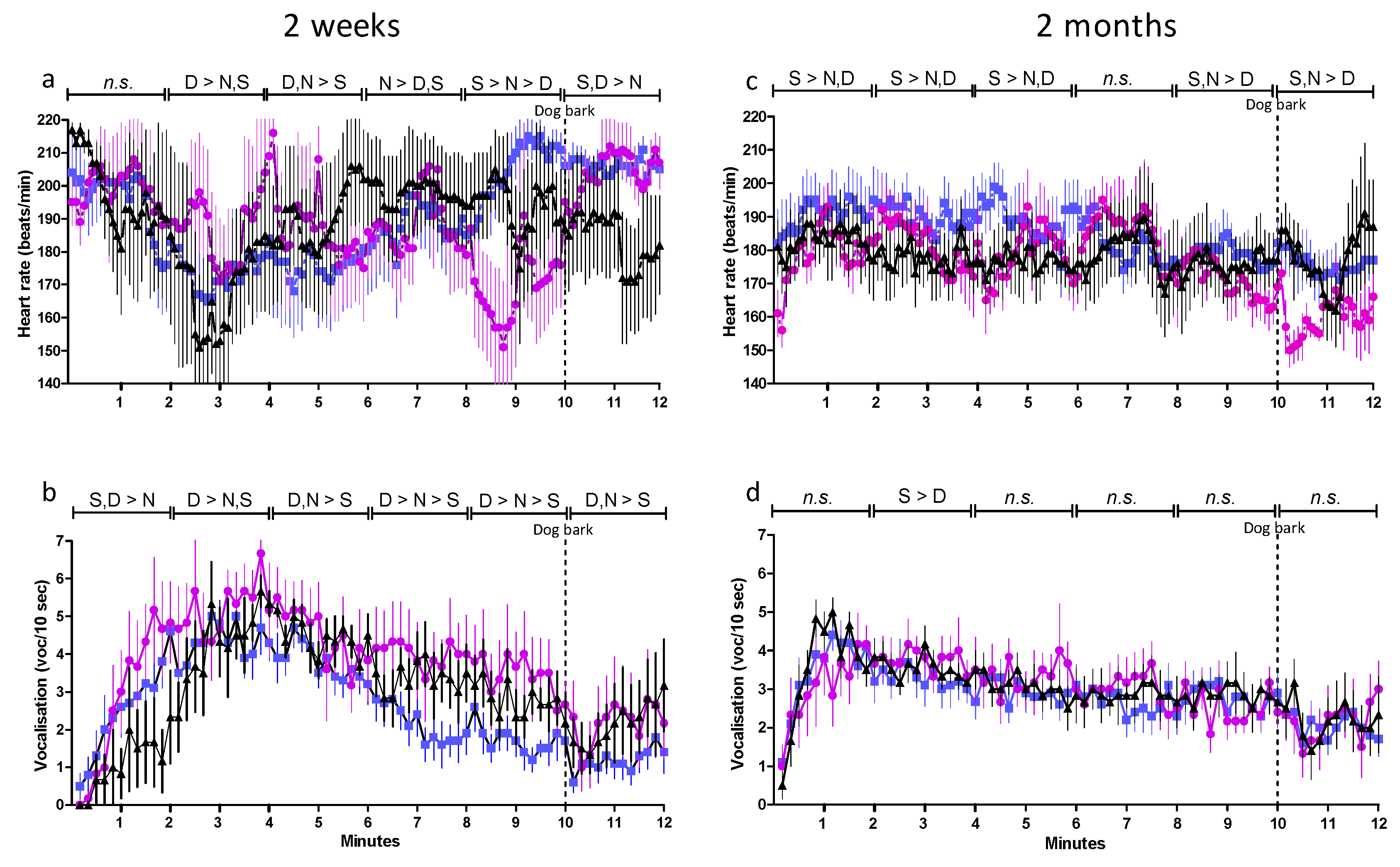
 ) DAY-SEP kids (
) DAY-SEP kids ( ) and NON-SEP kids (
) and NON-SEP kids ( ) during 12-min isolation test with the sound of a dog bark at 10 min. SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h–15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.
) DAY-SEP kids () and NON-SEP kids () during 12-min isolation test with the sound of a dog bark at 10 min. SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h–15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.
) during 12-min isolation test with the sound of a dog bark at 10 min. SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h–15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.
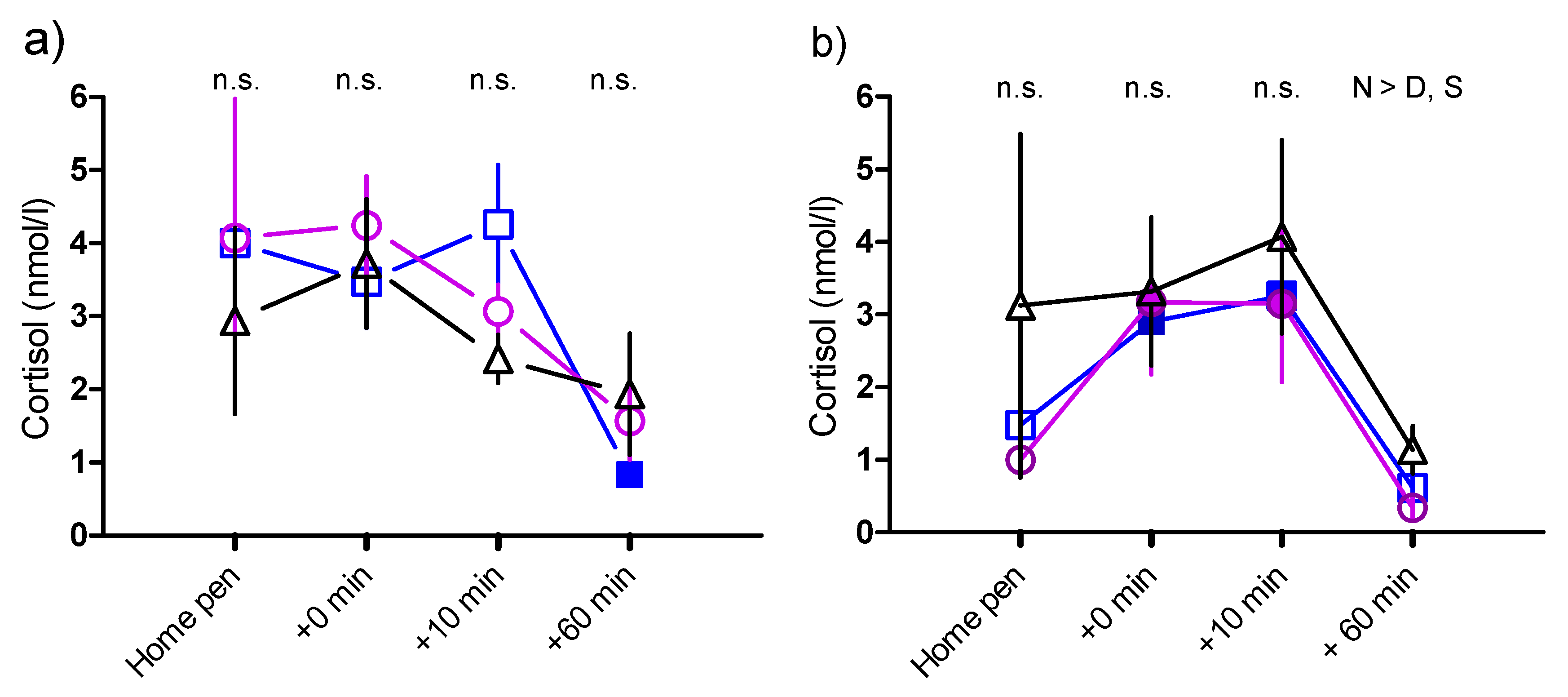
) DAY-SEP kids () and NON-SEP kids () during 12-min isolation test with the sound of a dog bark at 10 min. SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h–15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05. ) DAY-SEP kids () and NON-SEP kids () at 2 weeks (a) and 2 months (b) of age in home pen before isolation (Home pen), immediately after isolation, but before reunion (+0 min), 10 min after reunion with mother respectively kids group (+10 min), and 60 min after reunion (+60 min). SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. Filled symbols indicate that values are significantly different from the first sample. The level of significance was set at p ≤ 0.05.
) DAY-SEP kids () and NON-SEP kids () at 2 weeks (a) and 2 months (b) of age in home pen before isolation (Home pen), immediately after isolation, but before reunion (+0 min), 10 min after reunion with mother respectively kids group (+10 min), and 60 min after reunion (+60 min). SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. Filled symbols indicate that values are significantly different from the first sample. The level of significance was set at p ≤ 0.05.
) DAY-SEP kids () and NON-SEP kids () at 2 weeks (a) and 2 months (b) of age in home pen before isolation (Home pen), immediately after isolation, but before reunion (+0 min), 10 min after reunion with mother respectively kids group (+10 min), and 60 min after reunion (+60 min). SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. Filled symbols indicate that values are significantly different from the first sample. The level of significance was set at p ≤ 0.05.
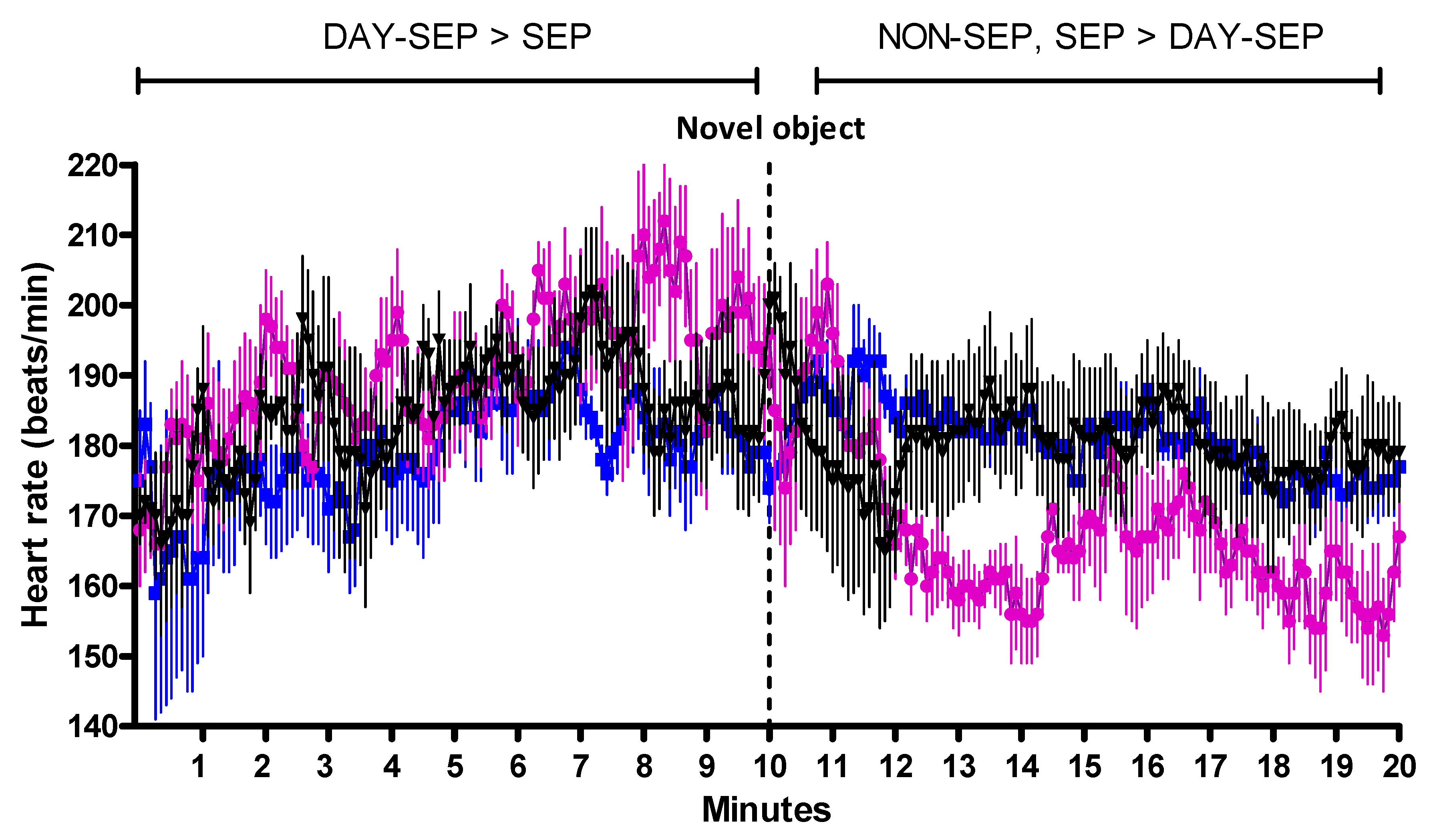
) DAY-SEP kids () and NON-SEP kids () at 2 weeks (a) and 2 months (b) of age in home pen before isolation (Home pen), immediately after isolation, but before reunion (+0 min), 10 min after reunion with mother respectively kids group (+10 min), and 60 min after reunion (+60 min). SEP = (S) 10 kids separated from their mother, DAY-SEP = (D) 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = (N) 6 kids kept together with mothers. > indicates significant differences between treatments stated in the figure. Filled symbols indicate that values are significantly different from the first sample. The level of significance was set at p ≤ 0.05. ) DAY-SEP kids () and NON-SEP kids () during an arena test. SEP = 10 kids separated from their mother, DAY-SEP = 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = 6 kids kept together with mothers. The arena test lasted for 20 min, with a novel object appearing after 10 min. For purposes of comparison, the arena test was divided into two periods; before novel object (PRE = minute 1–10) and after novel object (POST = minute 11–20). > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.
) DAY-SEP kids () and NON-SEP kids () during an arena test. SEP = 10 kids separated from their mother, DAY-SEP = 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = 6 kids kept together with mothers. The arena test lasted for 20 min, with a novel object appearing after 10 min. For purposes of comparison, the arena test was divided into two periods; before novel object (PRE = minute 1–10) and after novel object (POST = minute 11–20). > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.
) DAY-SEP kids () and NON-SEP kids () during an arena test. SEP = 10 kids separated from their mother, DAY-SEP = 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = 6 kids kept together with mothers. The arena test lasted for 20 min, with a novel object appearing after 10 min. For purposes of comparison, the arena test was divided into two periods; before novel object (PRE = minute 1–10) and after novel object (POST = minute 11–20). > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.
) DAY-SEP kids () and NON-SEP kids () during an arena test. SEP = 10 kids separated from their mother, DAY-SEP = 6 kids separated from their mother between 7.30 h and 15.00 h, and NON-SEP = 6 kids kept together with mothers. The arena test lasted for 20 min, with a novel object appearing after 10 min. For purposes of comparison, the arena test was divided into two periods; before novel object (PRE = minute 1–10) and after novel object (POST = minute 11–20). > indicates significant differences between treatments stated in the figure. The level of significance was set at p ≤ 0.05.

{kind=link}
{kind=link}
{kind=link}
| Goat | Lactation | Kids, nr and Gender | Treatment |
|---|---|---|---|
| 1 | 3 | 1, ♂ | DAY-SEP |
| 2, ♂ | SEP | ||
| 2 | 1 | 3, ♀ | NON-SEP |
| 4, ♂ | SEP | ||
| 3 | 1 | 5, ♂ | DAY-SEP |
| 6, ♂ | SEP | ||
| 7, ♂ | SEP | ||
| 4 | 3 | 8, ♀ | NON-SEP |
| 9 ♂ | SEP | ||
| 5 | 2 | 10, ♀ | DAY-SEP |
| 6 1 | 2 | 11, ♂ | SEP |
| 12, ♂ | SEP | ||
| 7 | 2 | 13, ♀ | NON-SEP |
| 14, ♀ | SEP | ||
| 8 | 3 | 15, ♀ | NON-SEP |
| 16, ♂ | SEP | ||
| 9 | 3 | 17, ♂ | DAY-SEP |
| 18, ♂ | SEP | ||
| 10 | 2 | 19, ♀ | NON-SEP |
| 11 2 | 3 | 20, ♀ | DAY-SEP |
| 21, ♀ | DAY-SEP | ||
| 12 | 2 | 22, ♂ | NON-SEP |
| Behavior | Description | Continuous Recording |
|---|---|---|
| Vocalization | The kid making sound, bleating | Counts per 10 s |
| Number of squares entered | New square was counted when entering an aligned square with one forefoot | Counts per 10 s |
| Jumping/rearing | Jumping against wall with all four feet, or rearing against wall with forefeet | Counts per 10 s |
| Startle reaction | Number of squares entered, and time from novel object until the kids were standing still more than 5 s | Counts of squares entered and seconds |
| Latency to sniff novel object | Latency between novel object on the floor and sniffing with muzzle less than 5 cm from novel object | Seconds |
| Instantaneous sampling | ||
| Location in arena | Wall: Located in one of the squares close to the wall | Proportion of observations |
| Standing | Standing on all four feet | Proportion of observations |
| Locomotion | Moving except jumping (walking, trotting, running or climbing bars) Standing still and walking/running are mutually exclusive | Proportion of observations |
| Exploring | Sniffing with the muzzle less than 5 cm from wall or floor | Proportion of observations |
| Treatment | SEP | DAY-SEP | NON-SEP | |||||
|---|---|---|---|---|---|---|---|---|
| Period | PRE | POST | PRE | POST | PRE | POST | p-Values 1 | p-Values 2 |
| Vocalizations No./10 s | 2.49 ± 0.07 a | 0.80 ± 0.04 c | 2.10 ± 0.09 b | 0.65 ± 0.06 c | 2.40 ± 0.07 ab | 0.41 ± 0.04 c | PRE 0.03 POST n.s. | SEP 0.0001 D-S 0.0001 N-S 0.0001 |
| Locomotion (proportion of observations) | 0.32 ± 0.01 a | 0.23 ± 0.01 c | 0.40 ± 0.02 b | 0.10 ± 0.01 c | 0.35 ± 0.02 ab | 0.15 ± 0.01 c | PRE 0.034 POST n.s. | SEP 0.0001 D-S 0.0001 N-S 0.0001 |
| Squares entered No./minute | 5.50 ± 0.37 a | 2.90 ± 0.3 c | 7.90 ± 0.52 b | 1.30 ± 0.35 c | 6.50 ± 0.50 ab | 2.10 ± 0.35 c | PRE 0.048 POST n.s. | SEP 0.0001 D-S 0.0001 N-S 0.0001 |
| Jumps No./minute | 0.62 ± 0.10 a | 0.10 ± 0.03 c | 1.70 ± 0.24 b | 0.02 ± 0.02 c | 1.00 ± 0.20 ab | 0.06 ± 0.03 c | PRE 0.0006 POST n.s. | SEP 0.0001 D-S 0.0001 N-S 0.0001 |
| Located by the Wall (proportion of observations) | 0.48 ± 0.02 a | 0.55 ± 0.02 c | 0.60 ± 0.02 b | 0.82 ± 0.02 d | 0.48 ± 0.02 ab | 0.63 ± 0.02 c | PRE 0.02 POST SEP vs D-S 0.002 POST D-S vs N-S 0.03 | SEP 0.002 D-S 0.0001 N-S 0.0002 |
| Exploring (proportion of observations) | 0.34 ± 0.02 a | 0.27 ± 0.01 b | 0.32 ± 0.02 a | 0.13 ± 0.01 c | 0.37 ± 0.02 a | 0.23 ± 0.02 bc | PRE n.s. POST 0.02 | SEP 0.002 D-S 0.0001 N-S 0.0001 |
| Treatment | SEP | DAY-SEP | NON-SEP |
|---|---|---|---|
| Home pen | 1.0 ± 0.2 a | 1.2 ± 0.2 a | 0.9 ± 0.3 a |
| After arena test | 1.9 ± 0.5 b | 2.5 ± 0.7 ab | 1.9 ± 0.7 ab |
| +60 min | 0.8 ± 0.1 a | 1.0 ± 0.3 a | 0.7 ± 0.3 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winblad von Walter, L.; Forkman, B.; Högberg, M.; Hydbring-Sandberg, E. The Effect of Mother Goat Presence during Rearing on Kids’ Response to Isolation and to an Arena Test. Animals 2021, 11, 575. https://doi.org/10.3390/ani11020575
Winblad von Walter L, Forkman B, Högberg M, Hydbring-Sandberg E. The Effect of Mother Goat Presence during Rearing on Kids’ Response to Isolation and to an Arena Test. Animals. 2021; 11(2):575. https://doi.org/10.3390/ani11020575
Chicago/Turabian StyleWinblad von Walter, Louise, Björn Forkman, Madeleine Högberg, and Eva Hydbring-Sandberg. 2021. "The Effect of Mother Goat Presence during Rearing on Kids’ Response to Isolation and to an Arena Test" Animals 11, no. 2: 575. https://doi.org/10.3390/ani11020575
APA StyleWinblad von Walter, L., Forkman, B., Högberg, M., & Hydbring-Sandberg, E. (2021). The Effect of Mother Goat Presence during Rearing on Kids’ Response to Isolation and to an Arena Test. Animals, 11(2), 575. https://doi.org/10.3390/ani11020575