
Nutritional Wasting Disorders in Sheep


 , , ,
, , , 
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Metabolic, Motility, and Management Disorders
2.1. Malnutrition
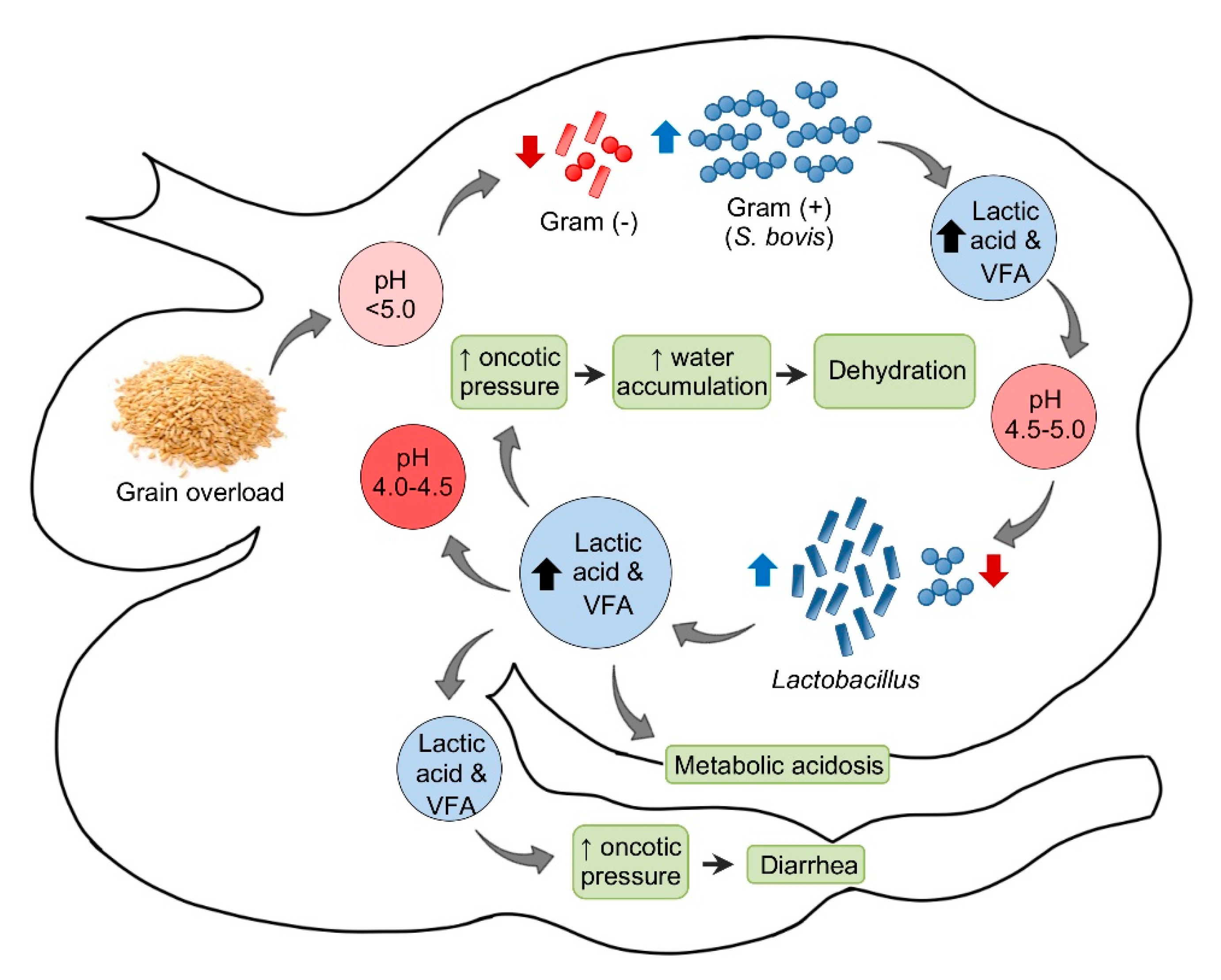
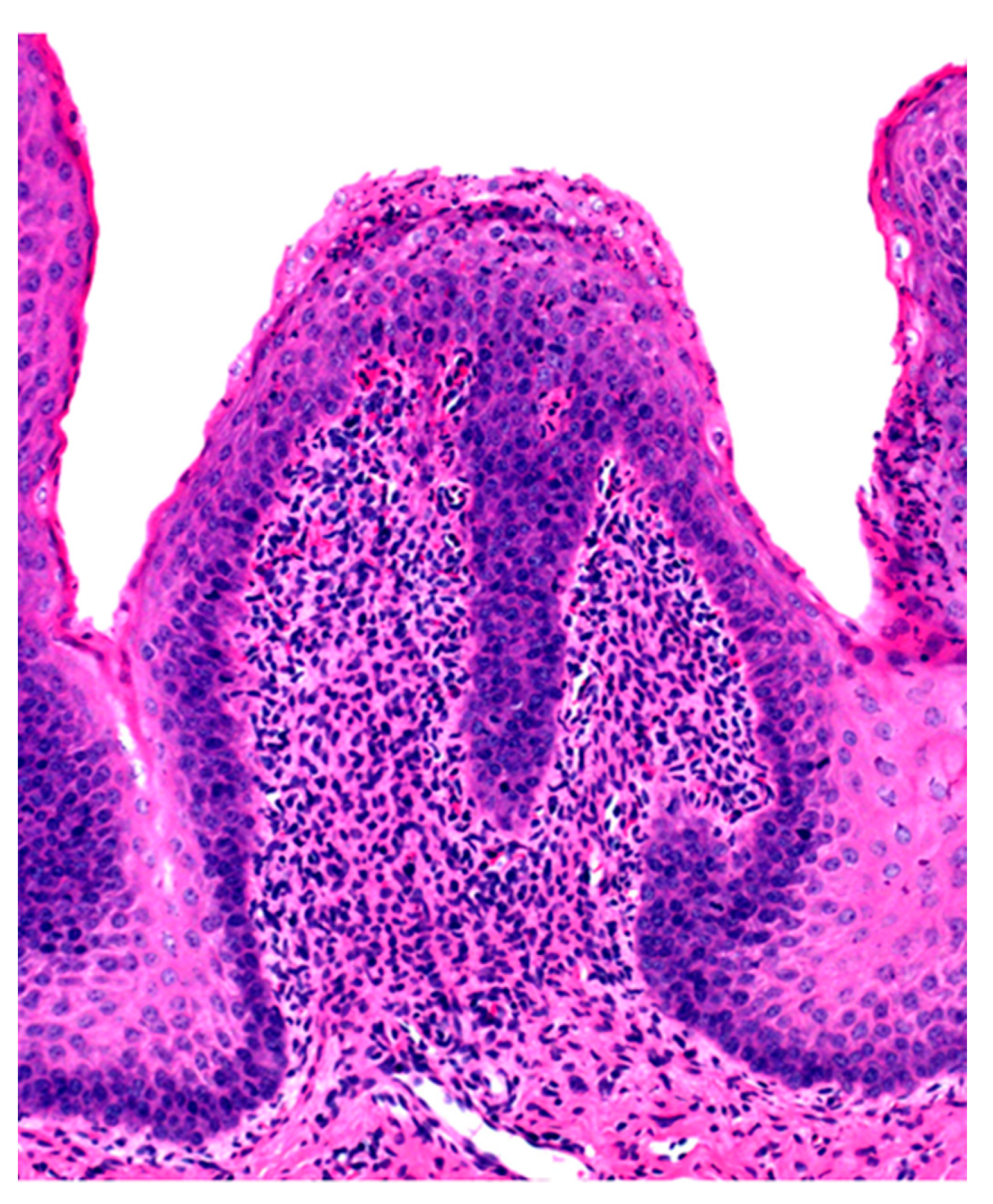
2.2. Subacute and Chronic Ruminal Acidosis
2.3. Vagal Indigestion Syndrome
2.4. Abomasal Emptying Defect
3. Mineral Deficiencies
3.1. Copper Deficiency
3.2. Cobalt Deficiency
3.3. Selenium/Vitamin E Deficiency
3.4. Zinc Deficiency
3.5. Iodine Deficiency
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Morris, S.T. Overview of sheep production systems. In Advances in Sheep Welfare, 1st ed.; Ferguson, D.M., Lee, C., Fisher, A., Eds.; Woodhead Publishing: Kidlington, UK, 2017; pp. 19–35. [Google Scholar]
- Lovatt, F. Developing flock health plans. Practice 2004, 26, 290–295. [Google Scholar] [CrossRef]
- Zajac, A.M. Gastrointestinal nematodes of small ruminants: Life cycle, anthelmintics, and diagnosis. Vet. Clin. N. Am. Food Anim. Pract. 2006, 22, 529–541. [Google Scholar] [CrossRef]
- Baird, G.J.; Fontaine, M.C. Corynebacterium pseudotuberculosis and its role in ovine caseous lymphadenitis. J. Comp. Pathol. 2007, 137, 179–210. [Google Scholar] [CrossRef] [PubMed]
- Windsor, P.A. Paratuberculosis in sheep and goats. Vet. Microbiol. 2015, 181, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Minguijón, E.; Reina, R.; Pérez, M.; Polledo, L.; Villoria, M.; Ramírez, H.; Leginagoikoa, I.; Badiola, J.J.; García-Marín, J.F.; de Andrés, D.; et al. Small ruminant lentivirus infections and diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef]
- Windsor, P.; Whittington, R. Ovine Paratuberculosis Control in Australia Revisited. Animals 2020, 10, 1623. [Google Scholar] [CrossRef]
- Winter, A.C.; Fitzpatrick, J.L. Sheep Welfare: Standards and Practices. In Diseases of Sheep, 4th ed.; Aitken, I.D., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 15–22. [Google Scholar]
- Abecia, J.A.; Sosa, C.; Forcada, F.; Meikle, A. The effect of undernutrition on the establishment of pregnancy in the ewe. Reprod. Nutr. Dev. 2006, 46, 367–378. [Google Scholar] [CrossRef]
- Stubbings, L.A. Ewe Management for Reproduction. In Diseases of Sheep, 4th ed.; Aitken, I.D., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 53–61. [Google Scholar]
- Morley, J.E.; Thomas, D.R.; Wilson, M.M. Cachexia: Pathophysiology and clinical relevance. Am. J. Clin. Nutr. 2006, 83, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Gerdin, J.A.; McDonough, S.P.; Reisman, R.; Scarlett, J. Circumstances, Descriptive Characteristics, and Pathologic Findings in Dogs Suspected of Starving. Vet. Pathol. 2016, 53, 1087–1094. [Google Scholar] [CrossRef]
- Russel, A. Body condition scoring of sheep. Practice 1984, 6, 91–93. [Google Scholar]
- Bush, R.D.; Toribio, J.A.; Windsor, P.A. The impact of malnutrition and other causes of losses of adult sheep in 12 flocks during drought. Aust. Vet. J. 2006, 84, 254–260. [Google Scholar] [CrossRef]
- Ruiz de Arcaute Rivero, M.; Ferrer-Mayayo, L.-M.; Lacasta, D.; González, J.; Las Heras, M.; Borobia, M.; Ramos, J.J. Prevalence of dental and mandibular disorders in culled sheep in Spain. Aust. Vet. J. 2020, 98, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Whiting, T.L.; Postey, R.C.; Chestley, S.T.; Wruck, G.C. Explanatory model of cattle death by starvation in Manitoba: Forensic evaluation. Can. Vet. J. 2012, 53, 1173–1180. [Google Scholar]
- Inadequate feeding and grazing leads to illthrift and death in cattle and sheep. Vet. Rec. 2018, 183, 51. [CrossRef] [PubMed]
- Uzal, F.A.; Plattner, B.L.; Hostetter, J.M. Alimentary System. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 2, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 1–257. [Google Scholar]
- Beeler-Marfisi, J.; Gallastegui Menoyo, A.; Beck, A.; König, J.; Hewson, J.; Bienzle, D. Gelatinous marrow transformation and hematopoietic atrophy in a miniature horse stallion. Vet. Pathol. 2011, 48, 451–455. [Google Scholar] [CrossRef]
- Meyerholtz, K.A.; Wilson, C.R.; Everson, R.J.; Hooser, S.B. Quantitative assessment of the percent fat in domestic animal bone marrow. J. Forensic Sci. 2011, 56, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Murden, D.; Hunnam, J.; De Groef, B.; Rawlin, G.; McCowan, C. Comparison of methodologies in determining bone marrow fat percentage under different environmental conditions. J. Vet. Diagn. Investig. 2017, 29, 83–90. [Google Scholar] [CrossRef]
- Sherman, D.M. Unexplained weight loss in sheep and goats. A guide to differential diagnosis, therapy, and management. Vet. Clin. N. Am. Large Anim. Pract. 1983, 5, 571–590. [Google Scholar] [CrossRef]
- Coop, R.L.; Holmes, P.H. Nutrition and parasite interaction. Int. J. Parasitol. 1996, 26, 951–962. [Google Scholar] [CrossRef]
- Van Metre, D.C.; Tyler, J.W.; Stehman, S.M. Diagnosis of enteric disease in small ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 87–115. [Google Scholar]
- Kleen, J.L.; Hooijer, G.A.; Rehage, J.; Noordhuizen, J.P. Subacute ruminal acidosis (SARA): A review. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2003, 50, 406–414. [Google Scholar] [CrossRef]
- Braun, U.; Rihs, T.; Schefer, U. Ruminal lactic acidosis in sheep and goats. Vet. Rec. 1992, 130, 343–349. [Google Scholar] [CrossRef]
- Lorentz, I. Subacute Ruminal Acidosis. In Merck Veterinary Manual. 2015. Available online: https://www.merckvetmanual.com/digestive-system/diseases-of-the-ruminant-forestomach/subacute-ruminal-acidosis (accessed on 29 November 2020).
- Randall, L.V.; Green, M.J.; Huxley, J.N. Use of statistical modelling to investigate the pathogenesis of claw horn disruption lesions in dairy cattle. Vet. J. 2018, 238, 41–48. [Google Scholar] [CrossRef]
- Cole, N.; Richardson, L.; Stock, R. Postmortem ruminal changes in sheep and steers. Vet. Clin. Nutr. 1998, 5, 14–17. [Google Scholar]
- Nordlund, K.V.; Garrett, E.F.; Oetzel, G.R. Herd-based rumenocentesis-a clinical approach to the diagnosis of sub-acute rumen acidosis. Comp. Cont. Educ. Pract. Vet. 1995, 17, 48–56. [Google Scholar]
- Mgasa, M.; Arnbjerg, J. Influence of diet on forestomach structure and occurrence of digital diseases in adult goats. Small Rumin. Res. 1993, 10, 63–73. [Google Scholar] [CrossRef]
- Constable, P.D.; Hinchcliff, K.W.; Done, S.H.; Grünberg, W.; Radostits, O.M. Diseases of the Alimentary Tract-Ruminant. In Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 11th ed.; Elsevier: St. Louis, MO, USA, 2017; pp. 436–621. [Google Scholar]
- Hoflund, S. Investigation in functional disorders of bovine stomach caused by lesions of the nervous vagus. Svensk. Vet. Tidskr. 1940. Cited in Whitlock, R.H. In Veterinary Gastroenterology; Anderson, N.V., Ed.; Lea and Febiger: Philadelphia, PA, USA, 1980. [Google Scholar]
- Malbert, C.H.; Ruckebusch, Y. Vagal influences on the phasic and tonic components of the motility of the ovine stomach and gastroduodenal area. Neurogastroent. Motil. 1989, 1, 15–20. [Google Scholar]
- Foster, D. Disorders of Rumen Distension and Dysmotility. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 499–512. [Google Scholar] [CrossRef]
- Lacasta, D.; Climent, M.; Ferrer, L.M.; Ramos, J.J.; Figueras, L.; Borobia, M. Vagus indigestion resulting from a Cysticercus tenuicollis cyst in an adult ewe. Small Rumin. Res. 2013, 110, 62–64. [Google Scholar] [CrossRef]
- Soares, G.S.L.; Firmino, P.R.; de Alencar Santos, D., Jr.; Frade, M.T.S.; Simoes, S.V.D.; de Miranda Neto, E.G. Unusual vagal indigestion in goat associated with liver abscess by Corynebacterium pseudotuberculosis. Vet. Zootec. 2017, 24, 691–697. [Google Scholar]
- Pruden, S.J.; McAllister, M.M.; Schultheiss, P.C.; O’Toole, D.; Christensen, D.E. Abomasal Emptying Defect of Sheep may be an Acquired Form of Dysautonomia. Vet. Pathol. 2004, 41, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Sharp, M.W.; Collings, D.F. Ovine abomasal enlargement and scrapie. Vet. Rec. 1987, 120, 215. [Google Scholar] [CrossRef]
- Gabb, K.; Lofstedt, J.; Bildfell, R. Abomasal emptying defect in a ewe of predominantly Dorset breeding. Vet. Rec. 1992, 131, 127–128. [Google Scholar] [CrossRef]
- Smith, R.M.; Underwood, W.J.; Petersen, G.C.; Harms, R.H. Abomasal dilatation and impaction in two Hampshire rams. Vet Rec. 1992, 130, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.L.; George, L.W.; East, N.E. Abomasal dilatation and emptying defect in a flock of Suffolk ewes. J. Am. Vet. Med. Assoc. 1988, 193, 1534–1536. [Google Scholar]
- Rings, D.M.; Welker, F.H.; Hull, B.L.; Kersting, K.W.; Hoffsis, G.F. Abomasal emptying defect in Suffolk sheep. J. Am. Vet. Med. Assoc. 1984, 185, 1520–1522. [Google Scholar]
- Suttle, N.F. Mineral Nutrition of Livestock, 4th ed.; CABI: Oxfordshire, UK, 2010. [Google Scholar]
- Suttle, N.F.; Jones, D.G. Micronutrient imbalance. In Diseases of Sheep, 4th ed.; Aitken, I.D., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 377–394. [Google Scholar]
- Hill, G.M.; Shannon, M.C. Copper and Zinc Nutritional Issues for Agricultural Animal Production. Biol. Trace Elem. Res. 2019, 188, 148–159. [Google Scholar] [CrossRef]
- Cantile, C.Y.; Youssef, S. Nervous system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volumen 1, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 250–406. [Google Scholar]
- Hall, J.O. Molybdenum. In Veterinary Toxicology, 3rd ed.; Gupta, R.C., Ed.; Elsevier: London, UK, 2018; pp. 463–467. [Google Scholar]
- De Sousa, I.K.; Hamad Minervino, A.H.; Sousa Rdos, S.; Chaves, D.F.; Soares, H.S.; Barros Ide, O.; de Araujo, C.A.; Junior, R.A.; Ortolani, E.L. Copper deficiency in sheep with high liver iron accumulation. Vet. Med. Int. 2012, 2012, 207950. [Google Scholar] [CrossRef]
- Miller, A.D.; Zachary, J.F. Nervous system. In Pathologic Basis of Veterinary Disease, 6th ed.; Zachary, J.F., Ed.; Elsevier: St. Louis, MO, USA, 2017; pp. 805–907. [Google Scholar]
- Mauldin, E.A.; Peters-Kennedy, J. Integumentary system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 1, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 509–736. [Google Scholar]
- Ataollahi, F.; Mohri, M.; Seifi, H.A.; Pingguan-Murphy, B.; Wan Abas, W.A.; Osman, N.A. Evaluation of copper concentration in subclinical cases of white muscle disease and its relationship with cardiac troponin I. PLoS ONE 2013, 8, e56163. [Google Scholar] [CrossRef] [PubMed]
- Suttle, N.F.; Angus, K.W.; Nisbet, D.I.; Field, A.C. Osteoporosis in copper-depleted lambs. J. Comp. Pathol. 1972, 82, 93–97. [Google Scholar] [CrossRef]
- Craig, L.E.D.; Dittmer, K.E.; Thompson, K.G. Bones and Joints. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 1, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 16–163. [Google Scholar]
- Vandevelde, M.; Higgins, R.; Oevermann, A. Metabolic-toxic diseases. In Veterinary Neuropathology: Essentials of Theory and Practice; Wiley Blackwell: Ames, IA, USA, 2012; pp. 118–119. [Google Scholar]
- Summers, B.A.; Cummings, J.F.; de Lahunta, A. Degenerative Diseases of the Central Nervous System. In Veterinary Neuropathology; Mosby Inc.: St. Louis, MO, USA, 1995; pp. 273–277. [Google Scholar]
- Kennedy, D.G.; Young, P.B.; Blanchflower, W.J.; Scott, J.M.; Weir, D.G.; Molloy, A.M.; Kennedy, S. Cobalt-vitamin B12 deficiency causes lipid accumulation, lipid peroxidation and decreased alpha-tocopherol concentrations in the liver of sheep. Int. J. Vitam. Nutr. Res. 1994, 64, 270–276. [Google Scholar] [PubMed]
- Mitchell, L.M.; Robinson, J.J.; Watt, R.G.; McEvoy, T.G.; Ashworth, C.J.; Rooke, J.A.; Dwyer, C.M. Effects of cobalt/vitamin B12 status in ewes on ovum development and lamb viability at birth. Reprod. Fertil. Dev. 2007, 19, 553–562. [Google Scholar] [CrossRef]
- Gruner, T.M.; Sedcole, J.R.; Furlong, J.M.; Sykes, A.R. A critical evaluation of serum methylmalonic acid and vitamin B12 for the assessment of cobalt deficiency of growing lambs in New Zealand. N. Zeal. Vet. J. 2004, 52, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.; McConnell, S.; Anderson, H.; Kennedy, D.G.; Young, P.B.; Blanchflower, W.J. Histopathologic and ultrastructural alterations of white liver disease in sheep experimentally depleted of cobalt. Vet. Pathol. 1997, 34, 575–584. [Google Scholar] [CrossRef]
- Sutherland, R.J.; Cordes, D.O.; Carthew, G.C. Ovine white liver disease--An hepatic dysfunction associated with vitamin B12 deficiency. N. Zeal. Vet. J. 1979, 27, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.M.; Stalker, M.J. Liver and biliary system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 2, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 258–352. [Google Scholar]
- Abou-Zeina, H.A.A.; Zaghawa, A.A.; Nasr, S.M.; Keshta, H.G.E. Effects of dietary cobalt deficiency on performance, blood and rumen metabolites and liver pathology in sheep. Glob. Vet. 2008, 2, 182–191. [Google Scholar]
- Vellema, P.; van den Ingh, T.S.; Wouda, W. Pathological changes in cobalt-supplemented and non-supplemented twin lambs in relation to blood concentrations of methylmalonic acid and homocysteine. Vet. Q. 1999, 21, 93–98. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Malevu, T.D.; Sochor, J.; Baron, M.; Melcova, M.; Zidkova, J.; et al. A Summary of New Findings on the Biological Effects of Selenium in Selected Animal Species-A Critical Review. Int. J. Mol. Sci. 2017, 18, 2209. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.J.; Valentine, B.A. Muscle and Tendon. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 1, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 16–249. [Google Scholar]
- Valentine, B.A. Skeletal muscle. In Pathologic Basis of Veterinary Disease; Zachary, J.F., Ed.; Elsevier: St. Louis, MO, USA, 2017; pp. 908–953. [Google Scholar]
- Kozat, S. Serum T3 and T4 concentrations in lambs with nutritional myodegeneration. J. Vet. Intern. Med. 2007, 21, 1135–1137. [Google Scholar] [CrossRef]
- Awawdeh, M.S.; Eljarah, A.H.; Ababneh, M.M. Multiple injections of vitamin E and selenium improved the reproductive performance of estrus-synchronized Awassi ewes. Trop. Anim. Health Prod. 2019, 51, 1421–1426. [Google Scholar] [CrossRef]
- Miller, L.M.; Gal, A. Cardiovascular system and lymphatic vessels. In Pathologic Basis of Veterinary Disease; Zachary, J.F., Ed.; Elsevier: St. Louis, MO, USA, 2017; pp. 561–616. [Google Scholar]
- Yildirim, S.; Ozkan, C.; Huyut, Z.; Cinar, A. Detection of Se, Vit. E, Vit. A, MDA, 8-OHdG, and CoQ10 Levels and Histopathological Changes in Heart Tissue in Sheep with White Muscle Disease. Biol. Trace Elem. Res. 2019, 188, 419–423. [Google Scholar] [CrossRef]
- Humann-Ziehank, E.; Renko, K.; Mueller, A.S.; Roehrig, P.; Wolfsen, J.; Ganter, M. Comparing functional metabolic effects of marginal and sufficient selenium supply in sheep. J. Trace. Elem. Med. Biol. 2013, 27, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Yaghmaie, P.; Ramin, A.; Asri-Rezaei, S.; Zamani, A. Evaluation of glutathion peroxidase activity, trace minerals and weight gain following administration of selenium compounds in lambs. Vet. Res. Forum 2017, 8, 133–137. [Google Scholar] [PubMed]
- Gunes, V.; Ozcan, K.; Citil, M.; Onmaz, A.C.; Erdogan, H.M. Detection of myocardial degeneration with point-of-care cardiac troponin assays and histopathology in lambs with white muscle disease. Vet. J. 2010, 184, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Molín, J.; Mendonça, F.S.; Henderson, E.E.; Nyaoke, A.A.; Ramírez, G.A.; Navarro, M.A.; Uzal, F.A.; Asín, J. Toxic wasting disorders in sheep. Animals 2021, 11, 229. [Google Scholar]
- Kojouri, G.A.; Shirazi, A. Serum concentrations of Cu, Zn, Fe, Mo and Co in newborn lambs following systemic administration of vitamin E and selenium to the pregnant ewes. Small Rumin. Res. 2007, 70, 136–139. [Google Scholar] [CrossRef]
- Smith, A.D.; Panickar, K.S.; Urban, J.F.; Dawson, H.D. Impact of Micronutrients on the Immune Response of Animals. Annu. Rev. Anim. Biosci. 2018, 6, 227–254. [Google Scholar] [CrossRef] [PubMed]
- White, C.L.; Martin, G.B.; Hynd, P.I.; Chapman, R.E. The effect of zinc deficiency on wool growth and skin and wool follicle histology of male Merino lambs. Br. J. Nutr. 1994, 71, 425–435. [Google Scholar] [CrossRef]
- Cousins, R.J.; Blanchard, R.K.; Moore, J.B.; Cui, L.; Green, C.L.; Liuzzi, J.P.; Cao, J.; Bobo, J.A. Regulation of Zinc Metabolism and Genomic Outcomes. J. Nutr. 2003, 133, 1521S–1526S. [Google Scholar] [CrossRef] [PubMed]
- Kwun, I.S.; Cho, Y.E.; Lomeda, R.A.R.; Kwon, S.T.; Kim, Y.; Beattie, J.H. Marginal zinc deficiency in rats decreases leptin expression independently of food intake and corticotrophin-releasing hormone in relation to food intake. Br. J. Nutr. 2007, 98, 485–489. [Google Scholar] [CrossRef]
- Martin, G.B.; White, C.L.; Markey, C.M.; Blackberry, M.A. Effects of dietary zinc deficiency on the reproductive system of young male sheep: Testicular growth and the secretion of inhibin and testosterone. J. Reprod. Fertil. 1994, 101, 87–96. [Google Scholar] [CrossRef]
- Droke, E.A.; Spears, J.W.; Spears, J.W. In vitro and in vivo immunological measurements in growing lambs fed diets deficient, marginal or adequate in zinc. J. Nutr. Immunol. 1993, 2, 71–90. [Google Scholar] [CrossRef]
- Valli, V.E.O.; Kiupel, M.; Bienzle, D.; Wood, R.D. Hematopoietic system. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 3, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 102–268. [Google Scholar]
- Knowles, S.O.; Grace, N.D. A practical approach to managing the risks of iodine deficiency in flocks using thyroid-weight: Birthweight ratios of lambs. N. Zeal. Vet. J. 2007, 55, 314–318. [Google Scholar] [CrossRef]
- Rosol, T.J.; Gröne, A. Endocrine glands. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 3, 6th ed.; Maxie, M.G., Ed.; W.B. Saunders: St. Louis, MO, USA, 2016; pp. 269–357. [Google Scholar]
- Follett, B.K.; Potts, C. Hypothyroidism affects reproductive refractoriness and the seasonal oestrous period in Welsh Mountain ewes. J. Endocrinol. 1990, 127, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.J.D.; Croser, E.L.; Milne, M.E.; Hodge, P.J.; Webb Ware, J.K. An outbreak of severe iodine-deficiency goitre in a sheep flock in north-east Victoria. Aust. Vet. J. 2012, 90, 235–239. [Google Scholar] [CrossRef]
- Sargison, N.D.; West, D.M.; Clark, R.G. An investigation of the possible effects of subclinical iodine deficiency on ewe fertility and perinatal lamb mortality. N. Zeal. Vet. J. 1997, 45, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Hill, R. Rapeseed meal in the diets of ruminants. Nutr. Abstr. Rev. B 1991, 61, 139–155. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asín, J.; Ramírez, G.A.; Navarro, M.A.; Nyaoke, A.C.; Henderson, E.E.; Mendonça, F.S.; Molín, J.; Uzal, F.A. Nutritional Wasting Disorders in Sheep. Animals 2021, 11, 501. https://doi.org/10.3390/ani11020501
Asín J, Ramírez GA, Navarro MA, Nyaoke AC, Henderson EE, Mendonça FS, Molín J, Uzal FA. Nutritional Wasting Disorders in Sheep. Animals. 2021; 11(2):501. https://doi.org/10.3390/ani11020501
Chicago/Turabian StyleAsín, Javier, Gustavo A. Ramírez, Mauricio A. Navarro, Akinyi C. Nyaoke, Eileen E. Henderson, Fábio S. Mendonça, Jéssica Molín, and Francisco A. Uzal. 2021. "Nutritional Wasting Disorders in Sheep" Animals 11, no. 2: 501. https://doi.org/10.3390/ani11020501
APA StyleAsín, J., Ramírez, G. A., Navarro, M. A., Nyaoke, A. C., Henderson, E. E., Mendonça, F. S., Molín, J., & Uzal, F. A. (2021). Nutritional Wasting Disorders in Sheep. Animals, 11(2), 501. https://doi.org/10.3390/ani11020501