
Unlocking the Role of a Genital Herpesvirus, Otarine Herpesvirus 1, in California Sea Lion Cervical Cancer

 , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. PCR and qPCR
2.3. Viral Genome Sequencing
2.4. Viral Phylogenetic Analysis
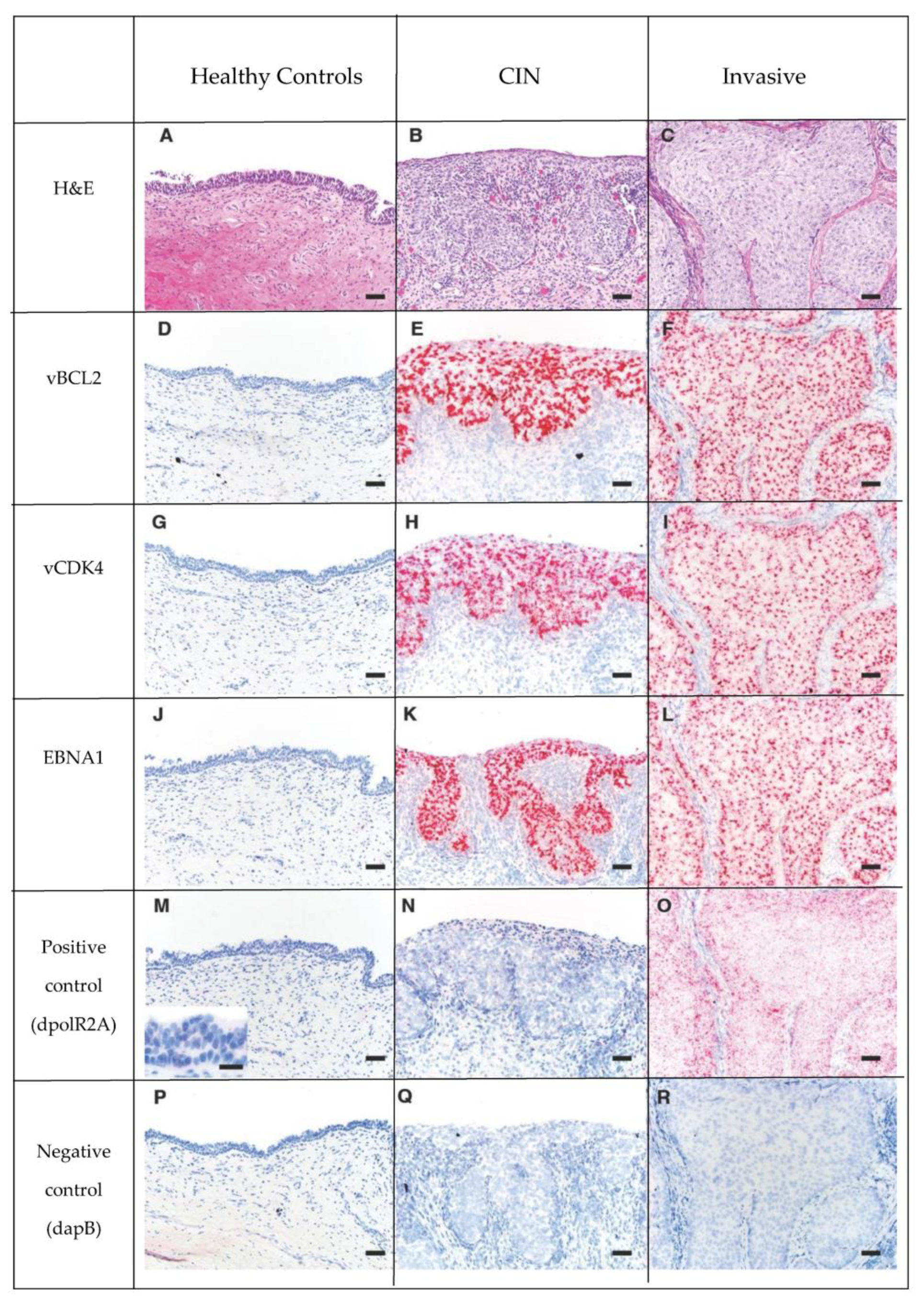
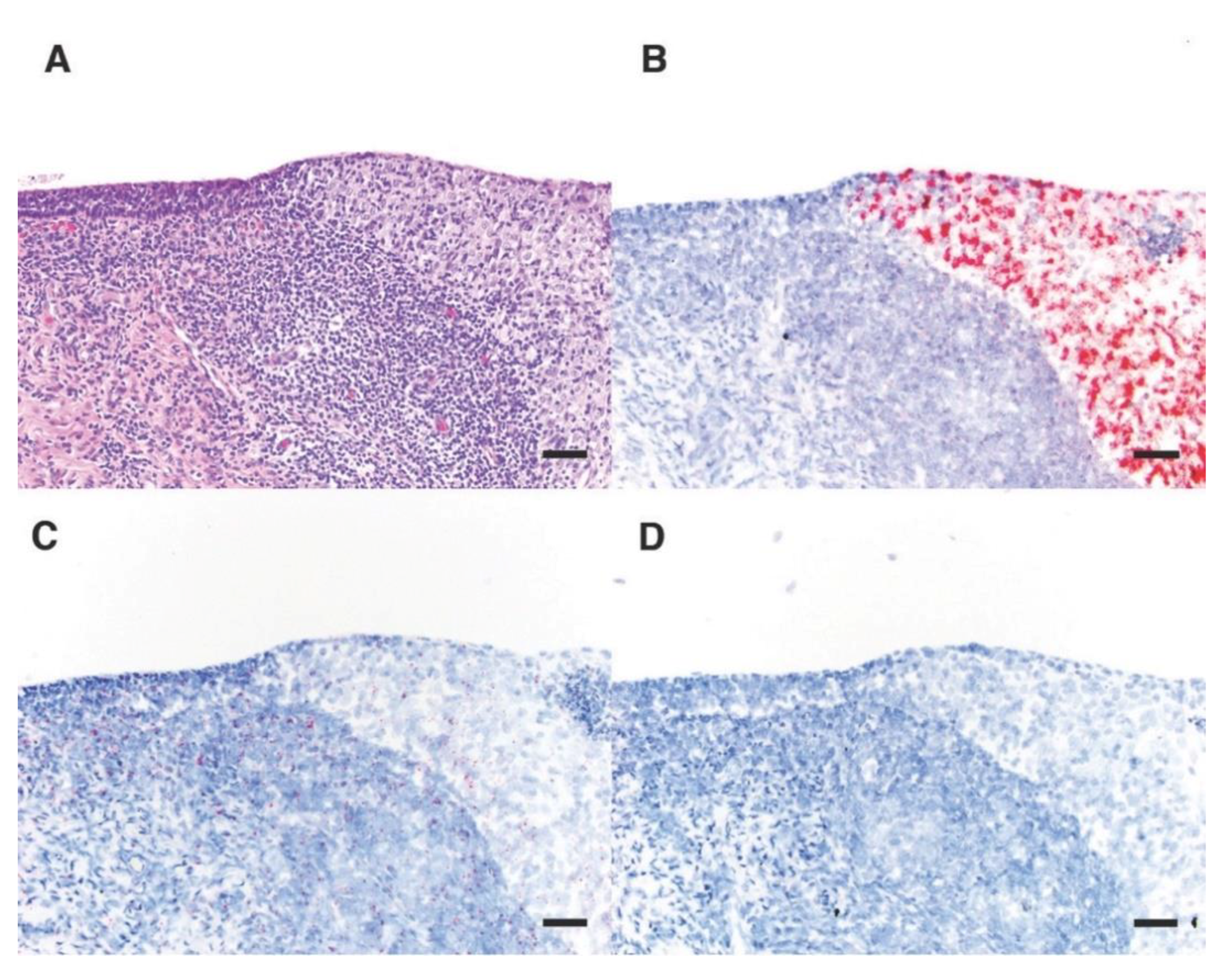
2.5. RNA In Situ Hybridization
3. Results
3.1. Histologic Determination of Urogenital Carcinoma and Control Cases
3.2. PCR and qPCR
3.2.1. OtHV1 PCR and qPCR Validation
3.2.2. OtHV1 qPCR Testing and Viral Quantification
3.3. OtHV1 and OtHV4 Sequencing and Annotation
3.4. Phylogenetic Analysis
3.5. OtHV1 mRNA Expression in Urogenital Carcinoma and Control Cases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gulland, F.M.D.; Trupkiewicz, J.G.; Spraker, T.R.; Lowenstine, L.J. Metastatic carcinoma of probable transitional cell origin in 66 free-living california sea lions (Zalophus californianus), 1979 TO 1994. J. Wildl. Dis. 1996, 32, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Deming, A.C.; Colegrove, K.M.; Duignan, P.J.; Hall, A.J.; Wellehan, J.J.F.X.; Gulland, F.M.D. Prevalence of Urogenital Carcinoma in Stranded California Sea Lions (Zalophus californianus) from 2005–15. J. Wildl. Dis. 2018, 54, 581–586. [Google Scholar] [CrossRef]
- Greig, D.J.; Gulland, F.M.D.; Kreuder, C. A Decade of Live California Sea Lion (Zalophus californianus) Strandings Along the Central California Coast: Causes and Trends, 1991–2000. Aquat. Mamm. 2005, 31, 11–22. [Google Scholar] [CrossRef]
- King, D.P.; Hure, M.C.; Goldstein, T.; Aldridge, B.M.; Gulland, F.M.D.; Saliki, J.T.; Buckles, E.L.; Lowenstine, L.J.; Stott, J.L. Otarine herpesvirus-1: A novel gammaherpesvirus associated with urogenital carcinoma in California sea lions (Zalophus californianus). Veter. Microbiol. 2002, 86, 131–137. [Google Scholar] [CrossRef]
- Lipscomb, T.P.; Scott, D.P.; Garber, R.L.; Krafft, A.E.; Tsai, M.M.; Lichy, J.H.; Taubenberger, J.K.; Schulman, F.Y.; Gulland, F.M.D. Common Metastatic Carcinoma of California Sea Lions (Zalophus californianus): Evidence of Genital Origin and Association with Novel Gammaherpesvirus. Veter. Pathol. 2000, 37, 609–617. [Google Scholar] [CrossRef]
- Weinberg, R.A. The Biology of Cancer, 2nd ed.; Garland Science: New York, NY, USA, 2014; Volume 1. [Google Scholar]
- Ylitalo, G.M.; Stein, J.E.; Hom, T.; Johnson, L.L.; Tilbury, K.L.; Hall, A.J.; Rowles, T.; Greig, D.; Lowenstine, L.J.; Gulland, F.M. The role of organochlorines in cancer-associated mortality in California sea lions (Zalophus californianus). Mar. Pollut. Bull. 2005, 50, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, N.; Gulland, F.M.D.; Ylitalo, G.M.; Delong, R.L.; Mazet, J.A. Sentinel California sea lions provide insight into legacy organochlorine exposure trends and their association with cancer and infectious disease. One Health 2015, 1, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Browning, H.M.; Acevedo-Whitehouse, K.; Gulland, F.M.D.; Hall, A.J.; Finlayson, J.; Dagleish, M.P.; Billington, K.J.; Colegrove, K.; Hammond, J.A. Evidence for a genetic basis of urogenital carcinoma in the wild California sea lion. Proc. R. Soc. B Biol. Sci. R. Soc. 2014, 281, 20140240. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Whitehouse, K.; Gulland, F.M.D.; Greig, D.J.; Amos, W. Disease susceptibility in California sea lions. Nat. Cell Biol. 2003, 422, 35. [Google Scholar] [CrossRef]
- Buckles, E.L.; Lowenstine, L.J.; Delong, R.L.; Melin, S.R.; Vittore, R.K.; Wong, H.-N.; Ross, G.L.; Leger, J.S.; Greig, D.J.; Duerr, R.S.; et al. Age-prevalence of Otarine Herpesvirus-1, a tumor-associated virus, and possibility of its sexual transmission in California sea lions. Veter. Microbiol. 2007, 120, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Barragán-Vargas, C.; Montano-Frías, J.; Rosales, G.Á.; Godínez-Reyes, C.R.; Acevedo-Whitehouse, K. Transformation of the genital epithelial tract occurs early in California sea lion development. R. Soc. Open Sci. 2016, 3, 150419. [Google Scholar] [CrossRef]
- Parkin, D.M. The global health burden of infection-associated cancers in the year 2002. Int. J. Cancer 2006, 118, 3030–3044. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin-Drubin, M.E.; Munger, K. Viruses associated with human cancer. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2008, 1782, 127–150. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-J.; You, S.-L.; Hsu, W.-L.; Yang, H.-I.; Lee, M.-H.; Chen, H.-C.; Chen, Y.-Y.; Liu, J.; Hu, H.-H.; Lin, Y.-J.; et al. Epidemiology of Virus Infection and Human Cancer. Toxic. Assess. 2021, 217, 13–45. [Google Scholar] [CrossRef]
- Honda, T. Potential inks between Hepadnavirus and Bornavirus sequences in the host genome and cancer. Front. Microbiol. 2017, 8, 2537. [Google Scholar] [CrossRef]
- Iizasa, H.; Nanbo, A.; Nishikawa, J.; Jinushi, M.; Yoshiyama, H. Epstein-Barr Virus (EBV)-associated Gastric Carcinoma. Viruses 2012, 4, 3420–3439. [Google Scholar] [CrossRef]
- Lancaster, W.D.; Olson, C. Animal papillomaviruses. Microbiol. Rev. 1982, 46, 191–207. [Google Scholar] [CrossRef]
- Jha, H.C.; Banerjee, S.; Robertson, E.S. The Role of Gammaherpesviruses in Cancer Pathogenesis. Pathogens 2016, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Klein, G. Herpesviruses and Oncogenesis. Proc. Natl. Acad. Sci. USA 1972, 69, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Rous, P. A transmissible avian neoplasm (sarcoma of the common fowl). 1910. Clin. Orthop. Relat. Res. 1993, 12, 696–705. [Google Scholar]
- Ewald, P. 99th Dahlem Conference on Infection, Inflammation and Chronic Inflammatory Disorders: Symbionts and immunopathology in chronic diseases: Insights from evolution. Clin. Exp. Immunol. 2010, 160, 27–34. [Google Scholar] [CrossRef]
- Renne, R.; Barry, C.; Dittmer, D.P.; Compitello, N.; Brown, P.O.; Ganem, D. Modulation of Cellular and Viral Gene Expression by the Latency-Associated Nuclear Antigen of Kaposi’s Sarcoma-Associated Herpesvirus. J. Virol. 2001, 75, 458–468. [Google Scholar] [CrossRef]
- Looker, K.J.; Magaret, A.S.; May, M.T.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global and Regional Estimates of Prevalent and Incident Herpes Simplex Virus Type 1 Infections in 2012. PLoS ONE 2015, 10, e0140765. [Google Scholar] [CrossRef]
- Looker, K.J.; Magaret, A.S.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global Estimates of Prevalent and Incident Herpes Simplex Virus Type 2 Infections in 2012. PLoS ONE 2015, 10, e114989. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.A.E.; Heuvel, D.V.D.; Bouthoorn, S.H.; Jaddoe, V.W.V.; Hooijkaas, H.; Raat, H.; Fraaij, P.L.; Van Zelm, M.C.; Moll, H.A. Determinants of Ethnic Differences in Cytomegalovirus, Epstein-Barr Virus, and Herpes Simplex Virus Type 1 Seroprevalence in Childhood. J. Pediatr. 2016, 170, 126–134.e6. [Google Scholar] [CrossRef] [PubMed]
- Verma, M. Cancer epidemiology. Host susceptibility factors. In Methods in Molecular Biology; Humana: New York, NY, USA, 2009; Volume 1, p. 1. [Google Scholar]
- Pipas, J.M.; Damania, B.; SpringerLink. DNA Tumor Viruses; Springer Science Business Media: New York, NY, USA, 2009; p. 1. [Google Scholar]
- Hernández-Ramírez, R.U.; Shiels, M.S.; Dubrow, R.; Engels, E.A. Cancer risk in HIV-infected people in the USA from 1996 to 2012: A population-based, registry-linkage study. Lancet HIV 2017, 4, e495–e504. [Google Scholar] [CrossRef]
- Frappier, L. EBNA1 and host factors in Epstein–Barr virus latent DNA replication. Curr. Opin. Virol. 2012, 2, 733–739. [Google Scholar] [CrossRef]
- Mesri, E.A.; Cesarman, E.; Boshoff, C. Kaposi’s sarcoma and its associated herpesvirus. Nat. Rev. Cancer 2010, 10, 707–719. [Google Scholar] [CrossRef]
- Grinde, B. Herpesviruses: Latency and reactivation—Viral strategies and host response. J. Oral Microbiol. 2013, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Falkow, S. Molecular Koch’s postulates applied to microbial pathogenicity. Rev. Infect. Dis. 1988, 10, S274–S276. [Google Scholar] [CrossRef] [PubMed]
- Fredericks, D.N.; Relman, D.A. Sequence-based identification of microbial pathogens: A reconsideration of Koch’s postulates. Clin. Microbiol. Rev. 1996, 9, 18–33. [Google Scholar] [CrossRef]
- Gulland, F.M.D.; Hall, A.J.; Ylitalo, G.M.; Colegrove, K.M.; Norris, T.; Duignan, P.J.; Halaska, B.; Whitehouse, K.A.; Lowenstine, L.J.; Deming, A.C.; et al. Persistent contaminants and herpesvirus OtHV1 are positively associated with cancer in wild California sea lions (Zalophus californianus). Front. Mar. Sci. 2020, 7, 1093. [Google Scholar] [CrossRef]
- Colegrove, K.M.; Gullanda, F.M.D.; Naydan, D.K.; Lowenstine, L.J. Tumor morphology and immunohistochemical expression of estrogen receptor, progesterone receptor, p53, and ki67 in urogenital carcinomas of California sea lions (Zalophus californianus). Veter. Pathol. 2009, 46, 642–655. [Google Scholar] [CrossRef]
- Colegrove, K.M.; Gulland, F.M.; Naydan, D.K.; Lowenstine, L.J. The normal genital tract of the female California sea lion (Zalophus californianus): Cyclic changes in histomorphology and hormone receptor distribution. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2009, 292, 1801–1817. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, A. Tumors of the cervix, vagina, and vulva. Atlas of tumor pathology. Third series, Fascicle 4. R. J. Kurman, H. J. Norris and E. J. Wilkinson, Armed Forces Institute of Pathology, Washington, DC, 1992. No. of pages: 262. Price: $45. ISBN: 11 881041 02 6. J. Pathol. 1993, 171, 245. [Google Scholar] [CrossRef]
- Cortés-Hinojosa, G.; Gulland, F.M.D.; Delong, R.; Gelatt, T.; Archer, L.; Wellehan, J.J.F.X. A novel gammaherpesvirus in northern fur seals (Callorhinus ursinus) is closely related to the california sea lion (Zalophus californianus) carcinoma-associated otarine herpesvirus-1. J. Wildl. Dis. 2016, 52, 88–95. [Google Scholar] [CrossRef]
- Frappier, L. Contributions of Epstein–Barr Nuclear Antigen 1 (EBNA1) to Cell Immortalization and Survival. Viruses 2012, 4, 1537–1547. [Google Scholar] [CrossRef]
- Cheng, E.H.; Nicholas, J.; Bellows, D.S.; Hayward, G.S.; Guo, H.G.; Reitz, M.S.; Hardwick, J.M. A Bcl-2 homo-log encoded by Kaposi sarcoma-associated virus, human herpesvirus 8, inhibits apoptosis but does not hetero-dimerize with Bax or Bak. Proc. Natl. Acad. Sci. USA 1997, 94, 690–694. [Google Scholar] [CrossRef]
- Mittnacht, S.; Boshoff, C. Viral cyclins. Rev. Med. Virol. 2000, 10, 175–184. [Google Scholar] [CrossRef]
- Davison, A.J.; Subramaniam, K.; Kerr, K.; Jacob, J.M.; Landrau-Giovannetti, N.; Walsh, M.T.; Wells, R.S.; Waltzek, T.B. Genome Sequence of a Gammaherpesvirus from a Common Bottlenose Dolphin (Tursiops truncatus). Genome Announc. 2017, 5, 31. [Google Scholar] [CrossRef] [PubMed]
- Damania, B. Oncogenic γ-herpesviruses: Comparison of viral proteins involved in tumorigenesis. Nat. Rev. Genet. 2004, 2, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Browning, H.M.; Gulland, F.; Hammond, J.A.; Colegrove, K.M.; Hall, A.J. Common cancer in a wild animal: The California sea lion (Zalophus californianus) as an emerging model for carcinogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140228. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer/Probe Name | Primers (5′-3′) | Reference |
|---|---|---|
| OtGHV1_polF | CTTCGCATGGGTGGACTACT | This Study |
| OtGHV1_polR | TCATGCCTACTAGCAGCAGC | This Study |
| OtGHV1qPCRF | TCCCACGCTGTTTCGAATG | This Study |
| OtGHV1qPCRR | AGCTCCGAGTCGTGTACACAGTAT | This Study |
| OtGHV1_Probe | TCGCGCTCGCATCGGCA | This Study |
| OtGHV4F2 | TCCACAATGATACTGGATGAAGA | Cortés-Hinojosa et al., 2016 |
| OtGHV4R4 | CTAGAATTGCACGACGCTGT | Cortés-Hinojosa et al., 2016 |
| QPCR_OtGHV4F2 | CTTCAACATTAGCTCCGGATT | Cortés-Hinojosa et al., 2016 |
| QPCR_OtGHV4R2 | CTTTACGCTTTGTTAGCCATGT | Cortés-Hinojosa et al., 2016 |
| QPCR_OtGHV4probe2 | AAAAAGCCATATATGTCAATCGCTACTATCAAA | Cortés-Hinojosa et al., 2016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deming, A.C.; Wellehan, J.F.X.; Colegrove, K.M.; Hall, A.; Luff, J.; Lowenstine, L.; Duignan, P.; Cortés-Hinojosa, G.; Gulland, F.M.D. Unlocking the Role of a Genital Herpesvirus, Otarine Herpesvirus 1, in California Sea Lion Cervical Cancer. Animals 2021, 11, 491. https://doi.org/10.3390/ani11020491
Deming AC, Wellehan JFX, Colegrove KM, Hall A, Luff J, Lowenstine L, Duignan P, Cortés-Hinojosa G, Gulland FMD. Unlocking the Role of a Genital Herpesvirus, Otarine Herpesvirus 1, in California Sea Lion Cervical Cancer. Animals. 2021; 11(2):491. https://doi.org/10.3390/ani11020491
Chicago/Turabian StyleDeming, Alissa C., James F. X. Wellehan, Kathleen M. Colegrove, Ailsa Hall, Jennifer Luff, Linda Lowenstine, Pádraig Duignan, Galaxia Cortés-Hinojosa, and Frances M. D. Gulland. 2021. "Unlocking the Role of a Genital Herpesvirus, Otarine Herpesvirus 1, in California Sea Lion Cervical Cancer" Animals 11, no. 2: 491. https://doi.org/10.3390/ani11020491
APA StyleDeming, A. C., Wellehan, J. F. X., Colegrove, K. M., Hall, A., Luff, J., Lowenstine, L., Duignan, P., Cortés-Hinojosa, G., & Gulland, F. M. D. (2021). Unlocking the Role of a Genital Herpesvirus, Otarine Herpesvirus 1, in California Sea Lion Cervical Cancer. Animals, 11(2), 491. https://doi.org/10.3390/ani11020491