
Distinct Pattern of NPY in Gastro–Entero–Pancreatic System of Goat Kids Fed with a New Standardized Red Orange and Lemon Extract (RLE)

 ,
,  , ,
, ,  , ,
, ,  ,
, 

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animals
2.2. Tissue Sampling
2.3. Single Immunostaining
Double Immunostaining
2.4. Controls of Specificity
2.5. Image Acquisition
2.6. Cell Counting
3. Results
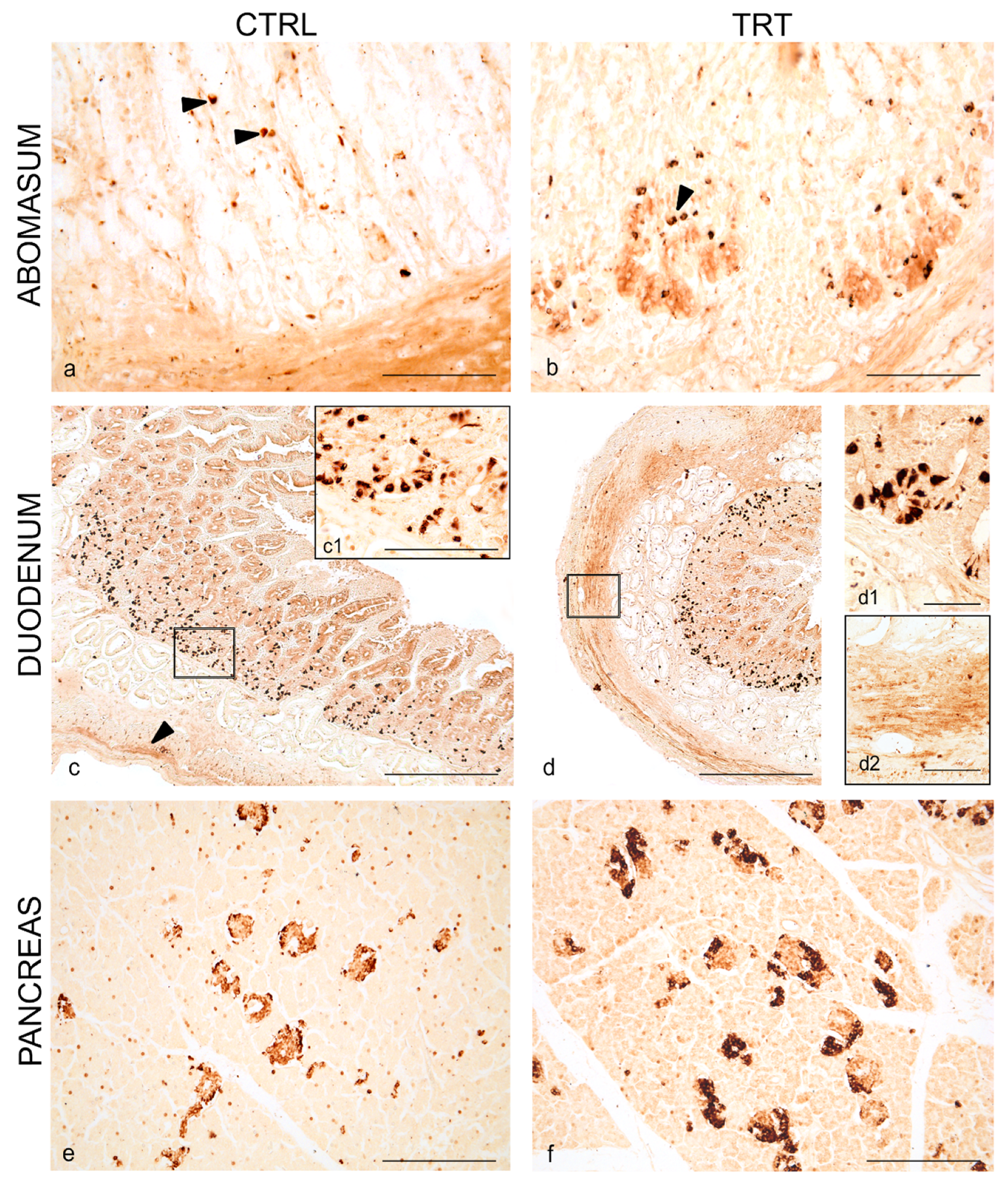
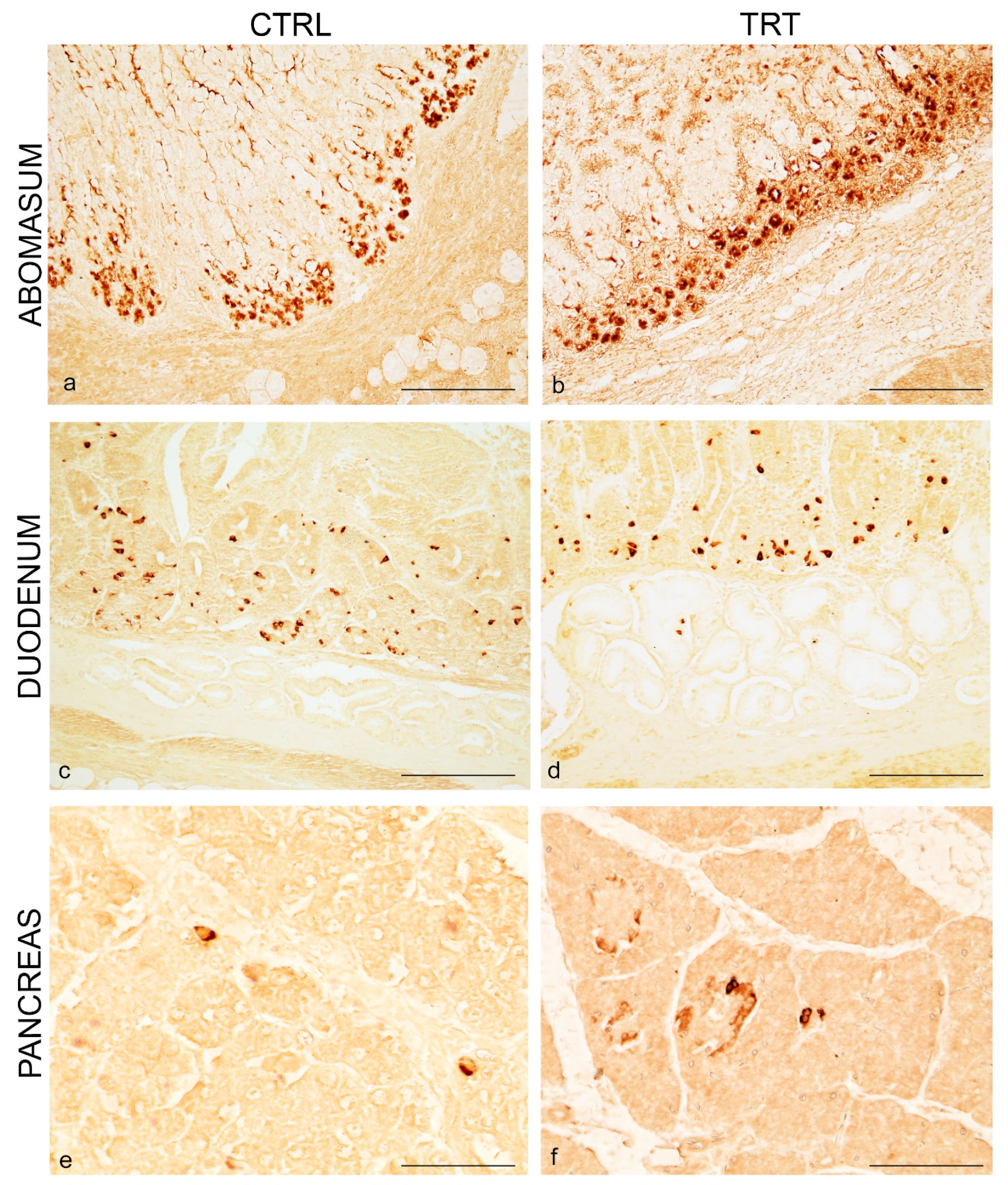
3.1. Distribution of NPY and 5-HT in the Abomasum, Duodenum and Pancreas of Control and Treated Animals
3.2. Co-Localization of NPY and 5-HT in the Abomasum, Duodenum and Pancreas
3.3. NPY Cell Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A review of antibiotic use in food animals: Perspective, policy, and potential. Publ. Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef]
- Ma, Z.; Lee, S.; Jeong, K.C. Mitigating antibiotic resistance at the livestock-environment Interface: A review. J. Microbiol. Biotechnol. 2019, 29, 1683–1692. [Google Scholar] [CrossRef]
- Cisowska, A.; Wojnicz, D.; Hendrich, A.B. Anthocyanins as antimicrobial agents of natural plant origin. Nat. Prod. Commun. 2011, 6, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wu, B.; Fu, W.; Reddivari, L. The anti-inflammatory effects of dietary anthocyanins against ulcerative colitis. Int. J. Mol. Sci. 2019, 20, 2588. [Google Scholar] [CrossRef]
- Hassimotto, N.M.; Moreira, V.; do Nascimento, N.G.; Souto, P.C.; Teixeira, C.; Lajolo, F.M. Inhibition of carrageenan-induced acute inflammation in mice by oral administration of anthocyanin mixture from wild mulberry and cyanidin-3-glucoside. Biomed. Res. Int. 2013, 2013, 146716. [Google Scholar] [CrossRef] [PubMed]
- Taverniti, V.; Fracassetti, D.; Del Bo’, C.; Lanti, C.; Minuzzo, M.; Klimis-Zacas, D.; Riso, P.; Guglielmetti, S. Immunomodulatory effect of a wild blueberry anthocyanin-rich extract in human Caco-2 intestinal cells. J. Agric. Food Chem. 2014, 62, 8346–8351. [Google Scholar] [CrossRef] [PubMed]
- Bicudo, M.O.; Ribani, R.H.; Beta, T. Anthocyanins, phenolic acids and antioxidant properties of Juçara fruits (Euterpe edulis M.) along the on-tree ripening process. Plant Foods Hum. Nutr. 2014, 69, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Anthocyanins induce the activation of phase II enzymes through the antioxidant response element pathway against oxidative stress-induced apoptosis. J. Agric. Food Chem. 2007, 55, 9427–9435. [Google Scholar] [CrossRef]
- Morais, C.A.; Vera de Rosso, V.; Estadella, D.; Pellegrini Pisani, L. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Igwe, E.O.; Charlton, K.E.; Probst, Y.C.; Kent, K.; Netzel, M.E. A systematic literature review of the effect of anthocyanins on gut microbiota populations. J. Hum. Nutr. Diet. 2018, 32, 53–62. [Google Scholar] [CrossRef]
- Hosoda, K.; Eruden, B.; Matsuyama, H.; Shioya, S. Effect of anthocyanin-rich corn silage on digestibility, milk production and plasma enzyme activities in lactating dairy cows. Anim. Sci. J. 2012, 83, 453–459. [Google Scholar] [CrossRef]
- Hosoda, K.; Sasahara, H.; Matsushita, K.; Tamura, Y.; Miyaji, M.; Matsuyama, H. Anthocyanin and proanthocyanidin contents, antioxidant activity and in situ degradability of black and red rice grains. Asian Australas J. Anim. Sci. 2018, 31, 1213–1220. [Google Scholar] [CrossRef]
- Tian, X.; Xin, H.; Paengkoum, P.; Paengkoum, S.; Ban, C.; Sorasak, T. Effects of anthocyanin-rich purple corn (Zea mays L.) stover silage on nutrient utilization, rumen fermentation, plasma antioxidant capacity, and mammary gland gene expression in dairy goats. J. Anim. Sci. 2019, 97, 1384–1397. [Google Scholar] [CrossRef]
- Jobstl, E.; O’Connell, J.; Fairclough, J.P.; Williamson, M.P. Molecular model for astringency produced by polyphenol/protein interactions. Biomacromolecules 2004, 5, 942–949. [Google Scholar] [CrossRef]
- González-Molina, E.; Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Natural bioactive compounds of citrus limon for food and health. J. Pharm. Biomed. Anal. 2010, 51, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Yamamoto, K.; Morimitsu, Y.; Osawa, T. Isolation of C-glucosylflavone from lemon peel and antioxidative activity of flavonoid compounds in lemon fruit. J. Agr. Food Chem. 1997, 45, 4619–4623. [Google Scholar] [CrossRef]
- Del Río, J.A.; Fuster, M.D.; Gómez, P.; Porras, I.; García-Lidón, A.; Ortuño, A. Citrus limon: A source of flavonoids of pharmaceutical interest. Food Chem. 2004, 84, 457–461. [Google Scholar] [CrossRef]
- Amenta, M.; Ballistreri, G.; Fabroni, S.; Romeo, F.V.; Spina, A.; Rapisarda, P. Qualitative and nutraceutical aspects of lemon fruits grown on the mountainsides of the Mount Etna: A first step for a protected designation of origin or protected geographical indication application of the brand name ‘Limone dell’Etna’. Food Res. Int. 2015, 74, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Fabroni, S.; Emma, R.; Ballistreri, G.; Amenta, M.; Currenti, W.; Rinzivillo, C.; Rapisarda, P. A new standardized phytoextract from red orange and lemon wastes (red orange and lemon extract) reduces basophil degranulation and activation. Nat. Prod. Res. 2020, 1–6. [Google Scholar] [CrossRef]
- Carota, G.; Raffaele, M.; Amenta, M.; Ballistreri, G.; Fabroni, S.; Rapisarda, P.; Vanella, L.; Sorrenti, V. In vitro effects of bioflavonoids rich lemon extract on pre-adipocyte differentiation. Nat. Prod. Res. 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, H.; Takenoya, F.; Hirako, S.; Wada, N.; Kintaka, Y.; Inoue, S.; Ota, E.; Ogawa, T.; Shioda, S. Neuronal circuits involving neuropeptide Y in hypothalamic arcuate nucleus-mediated feeding regulation. Neuropeptides 2012, 46, 285–289. [Google Scholar] [CrossRef]
- Garcia, A.; Masot, J.; Franco, A.; Gazquez, A.; Redondo, E. Immunohistochemical evaluation of the goat forestomach during prenatal development. J. Vet Sci. 2014, 15, 35–43. [Google Scholar] [CrossRef]
- Garcia, A.; Masot, J.; Franco, A.; Gazquez, A.; Redondo, E. Histomorphometric and immunohistochemical study of the goat abomasum during prenatal development. Histol. Histopathol. 2013, 28, 1639–1649. [Google Scholar] [PubMed]
- Badshah, H.; Ullah, I.; Kim, S.E.; Kim, T.; Lee, H.Y.; Kim, M.O. Anthocyanins attenuate body weight gain via modulating neuropeptide Y and GABAB1 receptor in rats hypothalamus. Neuropeptides 2013, 47, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Ibars, M.; Aragonès, G.; Ardid-Ruiz, A.; Gibert-Ramos, A.; Arola-Arnal, A.; Suárez, M.; Bladé, C. Seasonal consumption of polyphenol-rich fruits affects the hypothalamic leptin signaling system in a photoperiod-depend mode. Sci. Rep. 2018, 8, 3572. [Google Scholar] [CrossRef]
- Wu, X.; Pittman 3rd, H.E.; Prior, R.L. Fate of anthocyanins and antioxidant capacity in contents of the gastrointestinal tract of weanling pigs following black raspberry consumption. J. Agric. Food Chem. 2006, 54, 583–589. [Google Scholar] [CrossRef]
- Damiano, S.; Lombari, P.; Salvi, E.; Papale, M.; Giordano, A.; Amenta, M.; Ballistreri, G.; Fabroni, S.; Rapisarda, P.; Capasso, G.; et al. A red orange and lemon by-products extract rich in anthocyanins inhibits the progression of diabetic nephropathy. J. Cell. Physiol. 2019, 234, 23268–23278. [Google Scholar] [CrossRef]
- Maggiolino, A.; Lorenzo, J.M.; Quiñones, J.; Latorre, M.A.; Blando, F.; Centoducati, G.; Dahl, E.; De Palo, P. Effects of dietary supplementation with Pinus taeda hydrolyzed lignin on in vivo performances, in vitro nutrient apparent digestibility, and gas emission in beef steers. Anim. Feed Sci. Technol. 2019, 255, 114217. [Google Scholar] [CrossRef]
- Dall’Aglio, C.; Scocco, P.; Maranesi, M.; Petrucci, L.; Acuti, G.; De Felice, E.; Mercati, F. Immunohistochemical identification of resistin in the uterus of ewes subjected to different diets: Preliminary results. Eur. J. Histochem. 2019, 63, 3020. [Google Scholar] [CrossRef]
- Wessel, G.M.; McClay, D.R. Two embryonic, tissue-specific molecules identified by a double-label immunofluorescence technique for monoclonal antibodies. J. Histochem. Cytochem. 1986, 34, 703–706. [Google Scholar] [CrossRef] [PubMed]
- Gonkowski, S. Bisphenol A (BPA)-induced changes in the number of serotonin-positive cells in the mucosal layer of porcine small intestine-the preliminary studies. Int. J. Mol. Sci. 2020, 21, 1079. [Google Scholar] [CrossRef]
- Piovezana Gomes, J.V.; Buttow Rigolon, T.C.; Sampaio da Silveira Souza, M.; Alvarez-Leite, J.I.; Della Lucia, C.M.; Stampini Duarte Martino, H.; de Oliveira Barbosa Rosa, C. Antiobesity effects of anthocyanins on mitochondrial biogenesis, inflammation, and oxidative stress: A systematic review. Nutrition 2019, 66, 192–202. [Google Scholar] [CrossRef]
- Damiano, S.; Iovane, V.; Squillacioti, C.; Amenta, M.; Florio, S.; Ciarcia, R. Red orange and lemon extract prevents the renal toxicity induced by ochratoxin A in rats. J. Cell. Physiol. 2020, 235, 5386–5393. [Google Scholar] [CrossRef]
- Mora-Sánchez, B.; Fuertes, H.; Balcázar, J.L.; Perez-Sanchez, T. Effect of a multi-citrus extract-based feed additive on the survival of rainbow trout (Oncorhynchus mykiss) following challenge with Lactococcus garvieae. Acta Vet. Scand. 2020, 62, 38. [Google Scholar] [CrossRef]
- Lauro, M.R.; Crascì, L.; Giannone, V.; Ballistreri, G.; Fabroni, S.; Sansone, F.; Rapisarda, P.; Panico, A.M.; Puglisi, G. An alginate/cyclodextrin spray drying matrix to improve shelf life and antioxidant efficiency of a blood orange by-product extract rich in polyphenols: MMPs inhibition and antiglycation activity in dysmetabolic diseases. Oxid. Med. Cell. Long. 2017, 2017, 2867630. [Google Scholar] [CrossRef] [PubMed]
- Changxing, L.; Chenling, M.; Alagawany, M.; Jianhua, L.; Dongfang, D.; Gaichao, W.; Wenyin, Z.; Syed, S.F.; Arain, M.A.; Saeed, M.; et al. Health benefits and potential applications of anthocyanins in poultry feed industry. World’s Poult. Sci. J. 2018, 74, 251–264. [Google Scholar] [CrossRef]
- Passamonti, S.; Vrhovsek, U.; Vanzo, A.; Mattivi, F. The stomach as a site for anthocyanins absorption from food. FEBS Lett. 2003, 544, 210–213. [Google Scholar] [CrossRef]
- Matuschek, M.C.; Hendriks, W.H.; McGhie, T.K.; Reynolds, G.W. The jejunum is the main site of absorption for anthocyanins in mice. J. Nutr. Biochem. 2006, 17, 31–36. [Google Scholar] [CrossRef]
- Vona-Davis, L.C.; McFadden, D.W. NPY family of hormones: Clinical relevance and potential use in gastrointestinal disease. Curr. Top. Med. Chem. 2007, 7, 1710–1720. [Google Scholar] [CrossRef]
- Cox, H.M. Neuropeptide Y receptors; antisecretory control of intestinal epithelial function. Auton. Neurosci. 2007, 133, 76–85. [Google Scholar] [CrossRef]
- Sheikh, S.P. Neuropeptide Y and peptide YY: Major modulators modulators of gastrointestinal blood flow and function. Am. J. Physiol. 1991, 261, G701–G715. [Google Scholar] [CrossRef] [PubMed]
- Sipos, G.; Altdorfer, K.; Pongor, E.; Chen, L.P.; Fehér, E. Neuroimmune link in the mucosa of chronic gastritis with Helicobacter pylori infection. Dig. Dis. Sci. 2006, 51, 1810–1817. [Google Scholar] [CrossRef]
- Palus, K.; Calka, J. Neurochemical plasticity of the coeliac-superior mesenteric ganglion complex neurons projecting to the prepyloric area of the porcine stomach following hyperacidity. Neural Plast. 2016, 2016, 8596214. [Google Scholar] [CrossRef] [PubMed]
- McCauley, H. Enteroendocrine regulation of nutrient absorption. J. Nutr. 2020, 150, 10–21. [Google Scholar] [CrossRef]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The diverse metabolic roles of peripheral serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging roles for serotonin in regulating metabolism: New implications for an ancient molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Chappell, A.E.; Buret, A.G.; Barrett, K.E.; Dong, H. 5-Hydroxytryptamine contributes significantly to a reflex pathway by which the duodenal mucosa protects itself from gastric acid injury. FASEB J. 2006, 20, 2486–2495. [Google Scholar] [CrossRef] [PubMed]
- Diwakarla, S.; Fothergill, L.J.; Fakhry, J.; Callaghan, B.; Furness, J.B. Heterogeneity of enterochromaffin cells within the gastrointestinal tract. Neurogastroenterol. Motil. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Gribble, F.M.; Reimann, F. Enteroendocrine cells: Chemosensors in the intestinal epithelium. Ann. Rev. Physiol. 2016, 78, 277–299. [Google Scholar] [CrossRef] [PubMed]
- Almaça, J.; Molina, J.; Menegaz, D.; Pronin, A.N.; Tamayo, A.; Slepak, V.; Berggren, P.O.; Caicedo, A. Human beta cells produce and release serotonin to inhibit glucagon secretion from alpha cells. Cell. Rep. 2016, 17, 3281–3291. [Google Scholar] [CrossRef]
- Moon, J.H.; Kim, Y.G.; Kim, K.; Osonoi, S.; Wang, S.; Saunders, D.C.; Wang, J.; Yang, K.; Kim, H.; Lee, J.; et al. Serotonin regulates adult β-cell mass by stimulating perinatal β-cell proliferation. Diabetes 2020, 69, 205–214. [Google Scholar] [CrossRef]
- El-Merahbi, R.; Löffler, M.; Mayer, A.; Sumara, G. The roles of peripheral serotonin in metabolic homeostasis. FEBS Lett. 2015, 589, 1728–1734. [Google Scholar] [CrossRef]
- Wyler, S.C.; Lord, C.C.; Lee, S.; Elmquist, J.K.; Liu, C. Serotonergic control of metabolic homeostasis. Front. Cell. Neurosci. 2017, 11, 277. [Google Scholar] [CrossRef]
- Myojin, T.; Kitamura, N.; Hondo, E.; Baltazar, E.T.; Pearson, G.T.; Yamada, J. Immunohistochemical localization of neuropeptides in bovine pancreas. Anat. Histol. Embryol. 2000, 29, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Arciszewski, M.B.; Zacharko-Siembida, A. A co-localization study on the ovine pancreas innervation. Ann. Anat. 2007, 189, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Myrsen-Axcrona, U.; Ekblad, E.; Sundler, F. Developmental expression of NPY, PYY and PP in the rat pancreas and their coexistence with islet hormones. Regul. Pept. 1997, 68, 165–175. [Google Scholar] [CrossRef]
- Jackerott, M.; Oster, A.; Larsson, L. PYY in developing murine islet cells: Comparisons to development of islet hormones, NPY, and BrdU incorporation. J. Histochem. Cytochem. 1996, 44, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Stein, G.H.; Pan, N.; Goebbels, S.; Hornberg, H.; Nave, K.-A.; Herrera, P.; White, P.; Kaestner, K.H.; Sussel, L.; et al. Pancreatic beta cells require NeuroD to achieve and maintain functional maturity. Cell. Metab. 2010, 11, 298–310. [Google Scholar]
- Teitelman, G.; Alpert, S.; Polak, J.M.; Martinez, A.; Hanahan, D. Precursor cells of mouse endocrine pancreas coexpress insulin, glucagon and the neuronal proteins tyrosine hydroxylase and neuropeptide Y, but not pancreatic polypeptide. Development 1993, 118, 1031–1039. [Google Scholar]
- Myrsen, U.; Sundler, F. Neuropeptide Y is expressed in islet somatostatin cells of the hamster pancreas: A combined immunocytochemical and in situ hybridization study. Regul. Pept. 1995, 57, 65–76. [Google Scholar] [CrossRef]
- Whim, M.D. Pancreatic beta cells synthesize neuropeptide Y and can rapidly release peptide co-transmitters. PLoS ONE 2011, 6, e19478. [Google Scholar] [CrossRef] [PubMed]
- Machida, Y.; Bruinsma, C.; Hallinger, D.R.; Roper, S.M.; Garcia, E.; Trevino, M.B.; Nadler, J.; Ahima, R.; Imai, Y. Pancreatic islet Neuropeptide Y overexpression has minimal effect on islet morphology and β-cell adaptation to high-fat diet. Endocrinolgy 2014, 155, 4634–4640. [Google Scholar] [CrossRef] [PubMed]
- Opara, E.C.; Burch, W.M.; Taylor, I.L.; Akawi, O.E. Pancreatic hormone response to Neuropeptide Y (NPY) perifusion in vitro. Regul. Peptides 1991, 34, 225–233. [Google Scholar] [CrossRef]
- McGhie, T.K.; Ainge, G.D.; Barnett, L.E.; Cooney, J.M.; Jensen, D.J. Anthocyanin glycosides from berry fruit are absorbed and excreted unmetabolized by both humans and rats. J. Agric. Food Chem. 2003, 51, 4539–4548. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasam, B.; Vareed, S.K.; Olson, L.K.; Nair, M.G. Insulin secretion by bioactive anthocyanins and anthocyanidins present in fruits. J. Agric. Food Chem. 2005, 53, 28–31. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class of Compounds | Relative Composition (%) |
|---|---|
| Total flavanones | 15.91 ± 0.01 |
| Total Anthocyanins (as cyanidin 3-glucoside equivalents) | 2.66 ± 0.01 |
| Total Hydroxycinnamic acids | 1.77 ± 0.02 |
| Ascorbic acid | 2.40 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Felice, E.; Giaquinto, D.; Damiano, S.; Salzano, A.; Fabroni, S.; Ciarcia, R.; Scocco, P.; de Girolamo, P.; D’Angelo, L. Distinct Pattern of NPY in Gastro–Entero–Pancreatic System of Goat Kids Fed with a New Standardized Red Orange and Lemon Extract (RLE). Animals 2021, 11, 449. https://doi.org/10.3390/ani11020449
De Felice E, Giaquinto D, Damiano S, Salzano A, Fabroni S, Ciarcia R, Scocco P, de Girolamo P, D’Angelo L. Distinct Pattern of NPY in Gastro–Entero–Pancreatic System of Goat Kids Fed with a New Standardized Red Orange and Lemon Extract (RLE). Animals. 2021; 11(2):449. https://doi.org/10.3390/ani11020449
Chicago/Turabian StyleDe Felice, Elena, Daniela Giaquinto, Sara Damiano, Angela Salzano, Simona Fabroni, Roberto Ciarcia, Paola Scocco, Paolo de Girolamo, and Livia D’Angelo. 2021. "Distinct Pattern of NPY in Gastro–Entero–Pancreatic System of Goat Kids Fed with a New Standardized Red Orange and Lemon Extract (RLE)" Animals 11, no. 2: 449. https://doi.org/10.3390/ani11020449
APA StyleDe Felice, E., Giaquinto, D., Damiano, S., Salzano, A., Fabroni, S., Ciarcia, R., Scocco, P., de Girolamo, P., & D’Angelo, L. (2021). Distinct Pattern of NPY in Gastro–Entero–Pancreatic System of Goat Kids Fed with a New Standardized Red Orange and Lemon Extract (RLE). Animals, 11(2), 449. https://doi.org/10.3390/ani11020449