
Effects of Substitution of Corn with Ground Brown Rice on Growth Performance, Nutrient Digestibility, and Gut Microbiota of Growing-Finishing Pigs


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animals, and Diets
2.2. Sample Collection
2.3. Slaughter and Carcass Evaluation
2.4. Chemical Analysis
2.5. 16S rRNA Gene Sequencing and Analysis of Fecal Microbiota
2.6. Statistical Analysis
3. Results
3.1. Growth Performance, Nutrient Digestibility, and Carcass Characteristics
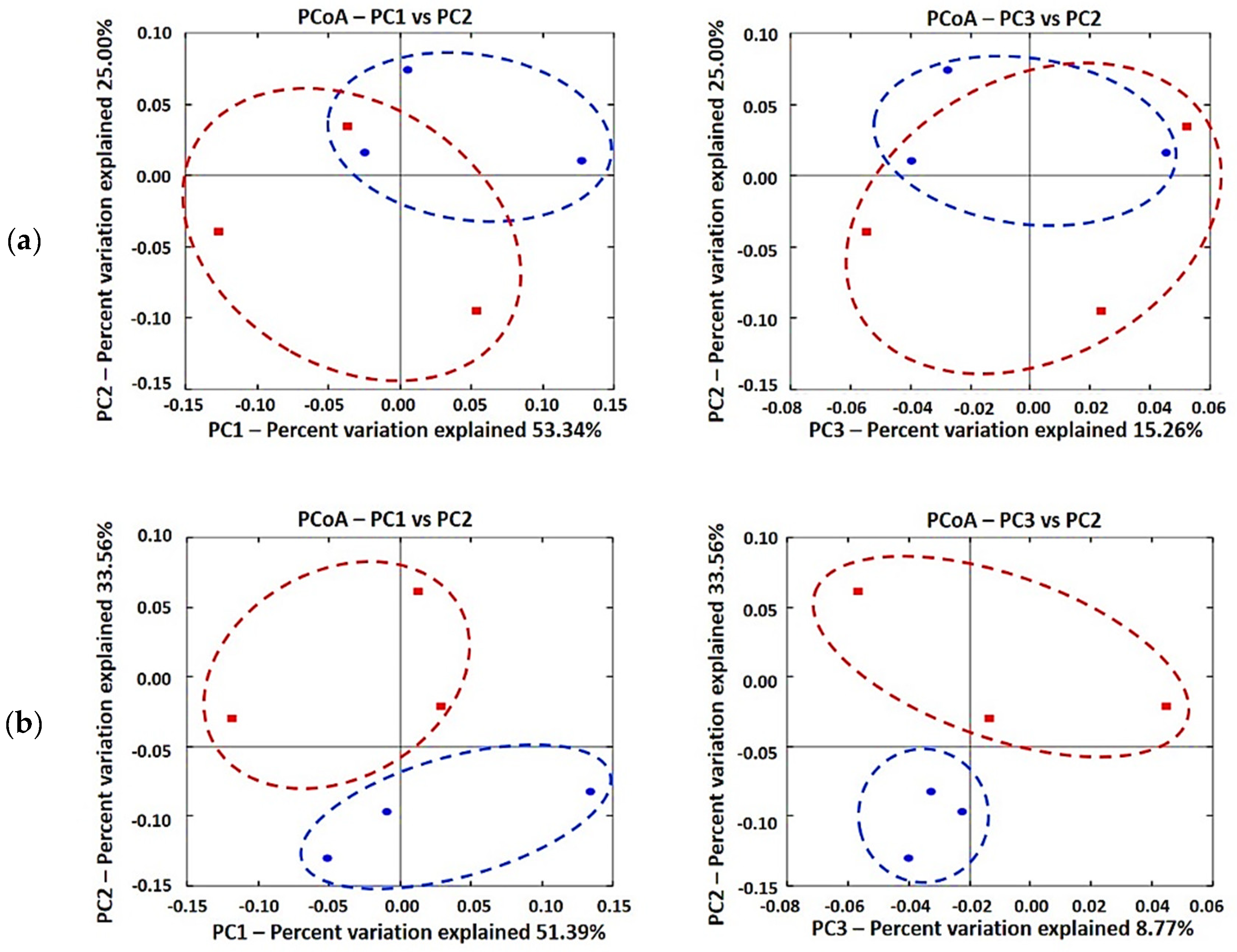
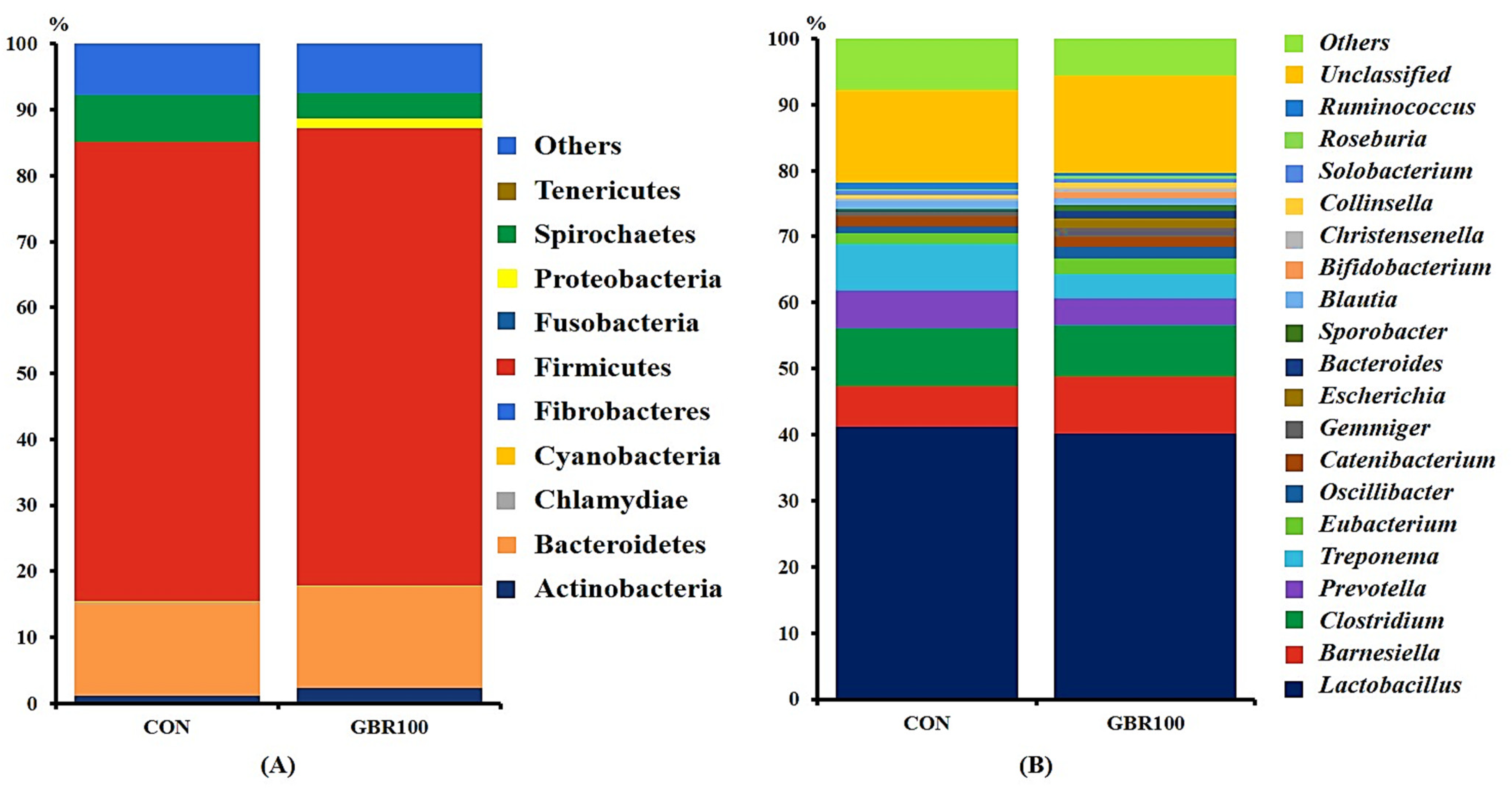
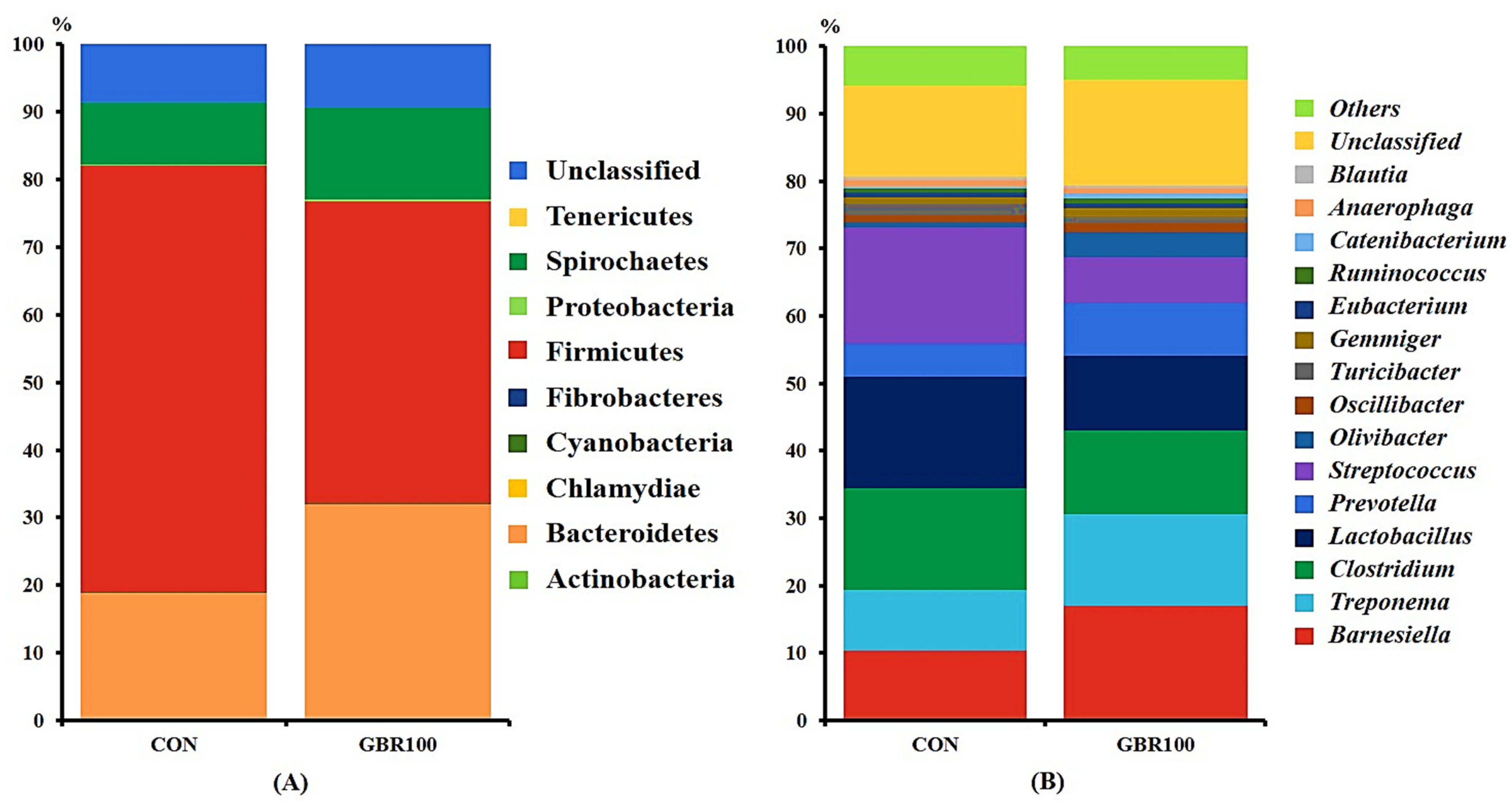
3.2. Diversity of Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nadeem, M.; Anjum, F.M.; Amir, R.M.; Khan, M.R.; Hussain, S.; Javed, M.S. An Overview of Anti-Nutritional Factors in Cereal Grains with Special Reference to Wheat-A Review. Pakistan J. Food Sci. 2010, 20, 54–61. [Google Scholar]
- Hopwood, D.E.; Pethick, D.W.; Pluske, J.R.; Hampson, D.J. Addition of pearl barley to a rice-based diet for newly weaned piglets increases the viscosity of the intestinal contents, reduces starch digestibility and exacerbates post-weaning colibacillosis. Br. J. Nutr. 2004, 92, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Vicente, B.; Valencia, D.G.; Pérez-Serrano, M.; Lázaro, R.; Mateos, G.G. The effects of feeding rice in substitution of corn and the degree of starch gelatinization of rice on the digestibility of dietary components and productive performance of young pigs. J. Anim. Sci. 2008, 86, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.; McCouch, S. The Complex History of the Domestication of Rice. Ann. Bot. 2007, 100, 951–957. [Google Scholar] [CrossRef]
- Bray, C.I. Rice and Rice Byproducts for Fattening Swine. In LSU Agricultural Experiment Station Reports; LSU Digital Commons: Louisiana, LO, USA, 1943; pp. 6–7. [Google Scholar]
- Tester, R.; Karkalas, J.; Qi, X. Starch—composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Ai, Y.; Jane, J.-L. Understanding Starch Structure and Functionality; Woodhead Publishing: Cambridge, UK, 2018; pp. 151–178. [Google Scholar]
- Popova, A.; Mihaylova, D. Antinutrients in Plant-based Foods: A Review. Open Biotechnol. J. 2019, 13, 68–76. [Google Scholar] [CrossRef]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 1–14. [Google Scholar] [CrossRef]
- Cho, M.K.; Kim, S.H.; Kang, M.Y. Application of Rice Polishing By-Products to Processed Rice Food—Antioxidative Effect of Black Rice Bran Pigment Fraction on Rice Embryo Lipid Oxidation. J. East Asian Soc. Diet. Life. 2008, 18, 361–367. [Google Scholar]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Jansman, A.J.M. Health and functions of the gastrointestinal tract in pigs: Effects of functional ingredients and feed and ingredient processing. J. Anim. Sci. 2016, 94, 12–21. [Google Scholar] [CrossRef]
- Li, X.L.; Yuan, S.L.; Piao, X.S.; Lai, C.H.; Zang, J.J.; Ding, Y.H.; Han, L.J.; Han, I.K. The Nutritional Value of Brown Rice and Maize for Growing Pigs. Asian-Australasian J. Anim. Sci. 2006, 19, 892–897. [Google Scholar] [CrossRef]
- Li, D.; Zhang, D.F.; Piao, X.S.; Han, I.K.; Yang, C.J.; Li, J.B.; Lee, J.H. Effects of Replacing Corn with Chinese Brown Rice on Growth Performance and Apparent Fecal Digestibility of Nutrients in Weanling Pigs. Asian-Australasian J. Anim. Sci. 2002, 15, 1191–1197. [Google Scholar] [CrossRef]
- Zhang, D.; Li, D.; Piao, X.S.; Han, I.K.; Yang, C.J.; Shin, I.S.; Dai, J.G.; Li, J.B. Effects of Replacing Corn with Brown Rice or Brown Rice with Enzyme on Growth Performance and Nutrient Digestibility in Growing Pigs. Asian-Australasian J. Anim. Sci. 2002, 15, 1334–1340. [Google Scholar] [CrossRef]
- Cromwell, G.L.; Henry, B.J.; Scott, A.L.; Gerngross, M.F.; Dusek, D.L.; Fletcher, D.W. Glufosinate herbicide-tolerant (LibertyLink) rice vs. conventional rice in diets for growing-finishing swine1,2. J. Anim. Sci. 2005, 83, 1068–1074. [Google Scholar] [CrossRef]
- Molina-Poveda, C. Nutrient requirements. Aquafeed Form. 2016, 75–216. [Google Scholar] [CrossRef]
- NPPC—National Pork Producers Council. Procedures to Evaluate Market Hogs, 3rd ed.; Pork Producers Council: Des Moines, IA, USA, 1991. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Williams, C.H.; David, D.J.; Iismaa, O. The determination of chromic oxide in faeces samples by atomic absorption spectrophotometry. J. Agric. Sci. 1962, 59, 381–385. [Google Scholar] [CrossRef]
- Stein, H.H.; Sève, B.; Fuller, M.F.; Moughan, P.J.; De Lange, C. Invited review: Amino acid bioavailability and digestibility in pig feed ingredients: Terminology and application. J. Anim. Sci. 2007, 85, 172–180. [Google Scholar] [CrossRef]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Hong, S.H.; Cho, J.H.; Kim, B.-R.; Shin, J.; Lee, J.H.; Na Kang, B.; Kim, Y.; Wattanaphansak, S.; Isaacson, R.E.; et al. The dynamics of the piglet gut microbiome during the weaning transition in association with health and nutrition. J. Anim. Sci. Biotechnol. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.-L.; Zheng, J.-B.; Chen, F.-S. Relationships between degree of milling and loss of Vitamin B, minerals, and change in amino acid composition of brown rice. LWT 2017, 82, 429–436. [Google Scholar] [CrossRef]
- Bird, A.R.; Hayakawa, T.; Marsono, Y.; Gooden, J.M.; Record, I.R.; Correll, R.L.; Topping, D.L. Coarse Brown Rice Increases Fecal and Large Bowel Short-Chain Fatty Acids and Starch but Lowers Calcium in the Large Bowel of Pigs. J. Nutr. 2000, 130, 1780–1787. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Yoshikawa, T.; Murakami, A. Effects of Brown Rice Particle Size on Growth Performance and Energy Digestibility in Finishing Pigs. Nihon Yoton Gakkaishi 2016, 53, 143–146. [Google Scholar] [CrossRef]
- Gao, G.H.; Dong, T.X. Pilot Study on Replacing Corn with Brown Rice in Duhu Pigs. Hubei Agric. Sci. China. 1993, 11, 25–26. [Google Scholar]
- Xiang, Y.; Tang, Q.; Huang, Y. The Relativity of Rice Grain Quality Characteristics I. Relations between Eating Quality and Other Grain Quality Characteristics of Indica Non-Waxy Rice. J. Hunan Agric. Coll. 1990, 16, 325–330. [Google Scholar]
- He, R.G.; Wang, Y.L.; Ma, L.B.; Li, M.; Zhang, S.X. Nutritional Value of Early Long-Grain Brown Rice in Hubei Province: 2. Effect of Substitution of Brown Rice for Maize as Energy Feedstuff on the Growth and Meat Quality of Growing- Finishing Pigs. J. Chin. Cereal. Oils Assoc. 2000, 15, 50–53. [Google Scholar]
- He, R.G.; Ma, Y.L.; Wang, Y.Q.; Zhao, J.Y.; Wang, H.X. Study of the Brown Rice Nutritional Value by the Pig’s Digestion and Metabolism Trial. J. Huazhong Agric. Univ. 1994, 13, 268–273. [Google Scholar]
- Li, T.; Huang, R.; Wu, G.; Lin, Y.; Jiang, Z.; Kong, X.; Chu, W.; Zhang, Y.; Kang, P.; Hou, Z.; et al. Growth performance and nitrogen metabolism in weaned pigs fed diets containing different sources of starch. Livest. Sci. 2007, 109, 73–76. [Google Scholar] [CrossRef]
- MacGregor, A.W.; Ballance, D.L. Hydrolysis of Large and Small Starch Granules from Normal and Waxy Barley Cultivas by Alpha-Amylases from Barley Malt. Cereal Chem. 1980, 57, 397–402. [Google Scholar]
- Tester, R.; Qi, X.; Karkalas, J. Hydrolysis of native starches with amylases. Anim. Feed. Sci. Technol. 2006, 130, 39–54. [Google Scholar] [CrossRef]
- Puncha-Arnon, S.; Pathipanawat, W.; Puttanlek, C.; Rungsardthong, V.; Uttapap, D. Effects of relative granule size and gelatinization temperature on paste and gel properties of starch blends. Food Res. Int. 2008, 41, 552–561. [Google Scholar] [CrossRef]
- Casas, G.; Almeida, J.; Stein, H. Amino acid digestibility in rice co-products fed to growing pigs. Anim. Feed. Sci. Technol. 2015, 207, 150–158. [Google Scholar] [CrossRef]
- Casas, G.A.; Stein, H.H. Effects of microbial xylanase on digestibility of dry matter, organic matter, neutral detergent fiber, and energy and the concentrations of digestible and metabolizable energy in rice coproducts fed to weanling pigs1. J. Anim. Sci. 2016, 94, 1933–1939. [Google Scholar] [CrossRef]
- Snow, P.; O’Dea, K. Factors affecting the rate of hydrolysis of starch in food. Am. J. Clin. Nutr. 1981, 34, 2721–2727. [Google Scholar] [CrossRef]
- Piao, X.S.; Li, D.; Han, I.K.; Chen, Y.; Lee, J.H.; Wang, D.Y.; Li, J.B.; Zhang, D.F. Evaluation of Chinese Brown Rice as an Alternative Energy Source in Pig Diets. Asian-Australasian J. Anim. Sci. 2002, 15, 89–93. [Google Scholar] [CrossRef]
- Yuichiro, W.; Hideaki, O.; Daisuke, U.; Hirofumi, K.; Hideo, M.; Sadayuki, A.; Shigeki, N.; Mitsuhito, M.; Hiroaki, I. Effects of Graded Replacement of Corn by Brown Rice in Diet Containing Barley and Tea Manufacturing Residual on Growth Performance and Meat Quality of Finishing Pigs in Summer. Jpn. J. Swine Sci. 2013, 50, 147–156. [Google Scholar]
- Katsumata, M.; Akane, A.; Aiko, I.; Hiroyuki, K. Effects of Replacement of All of Corn Contained in Feed with Brown Rice and Feeding Brown Rice Together with Sweet Potato on Growth Performance and Quality of Pork of Fattening Pigs. Jpn. J. Swine Sci. 2015, 52, 17–28. [Google Scholar] [CrossRef]
- Isaacson, R.; Kim, H.B. The intestinal microbiome of the pig. Anim. Health Res. Rev. 2012, 13, 100–109. [Google Scholar] [CrossRef]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Leser, T.D.; Lindecrona, R.H.; Jensen, T.K.; Jensen, B.B.; Møller, K. Changes in Bacterial Community Structure in the Colon of Pigs Fed Different Experimental Diets and after Infection with Brachyspira hyodysenteriae. Appl. Environ. Microbiol. 2000, 66, 3290–3296. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.T.S.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of dietary protein on microbiota composition and activity in the gastrointestinal tract of piglets in relation to gut health: A review. Animals 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van De Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Fouhse, J.; Zijlstra, R.; Willing, B. The role of gut microbiota in the health and disease of pigs. Anim. Front. 2016, 6, 30–36. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef]
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The Impact of Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 1–18. [Google Scholar] [CrossRef]
- McCormack, U.M.; Curiao, T.; Buzoianu, S.G.; Prieto, M.L.; Ryan, T.; Varley, P.; Crispie, F.; Magowan, E.; Metzler-Zebeli, B.U.; Berry, D.; et al. Exploring a Possible Link between the Intestinal Microbiota and Feed Efficiency in Pigs. Appl. Environ. Microbiol. 2017, 83, e00380-17. [Google Scholar] [CrossRef]
- Han, G.G.; Lee, J.-Y.; Jin, G.-D.; Park, J.; Choi, Y.H.; Kang, S.-K.; Chae, B.J.; Kim, E.B.; Choi, Y.J. Tracing of the fecal microbiota of commercial pigs at five growth stages from birth to shipment. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Tan, Z.; Wang, Y.; Yang, T.; Ao, H.; Chen, S.; Xing, K.; Zhang, F.; Zhao, X.; Liu, J.; Wang, C. Differences in gut microbiota composition in finishing Landrace pigs with low and high feed conversion ratios. Antonie van Leeuwenhoek 2018, 111, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Quan, J.; Wu, Z.; Ye, Y.; Peng, L.; Wu, J.; Ruan, D.; Qiu, Y.; Ding, R.; Wang, X.; Zheng, E.; et al. Metagenomic Characterization of Intestinal Regions in Pigs With Contrasting Feed Efficiency. Front. Microbiol. 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Bian, G.; Su, Y.; Zhu, W.-Y. Comparison of Faecal Microbial Community of Lantang, Bama, Erhualian, Meishan, Xiaomeishan, Duroc, Landrace, and Yorkshire Sows. Asian-Australasian J. Anim. Sci. 2014, 27, 898–906. [Google Scholar] [CrossRef]
- Mueller, S.; Saunier, K.; Hanisch, C.; Norin, E.; Alm, L.; Midtvedt, T.; Cresci, A.; Silvi, S.; Orpianesi, C.; Verdenelli, M.C.; et al. Differences in Fecal Microbiota in Different European Study Populations in Relation to Age, Gender, and Country: A Cross-Sectional Study. Appl. Environ. Microbiol. 2006, 72, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Le Sciellour, M.; Labussière, E.; Zemb, O.; Renaudeau, D. Effect of dietary fiber content on nutrient digestibility and fecal microbiota composition in growing-finishing pigs. PLoS ONE 2018, 13, e0206159. [Google Scholar] [CrossRef]
- Yang, H.; Xiang, Y.; Robinson, K.; Wang, J.; Zhang, G.; Zhao, J.; Xiao, Y. Gut Microbiota Is a Major Contributor to Adiposity in Pigs. Front. Microbiol. 2018, 9, 3045. [Google Scholar] [CrossRef]
- Park, S.-J.; Kim, J.; Lee, J.-S.; Rhee, S.-K.; Kim, H. Characterization of the fecal microbiome in different swine groups by high-throughput sequencing. Anaerobe 2014, 28, 157–162. [Google Scholar] [CrossRef]
- Knecht, D.; Cholewińska, P.; Jankowska-Mąkosa, A.; Czyż, K. Development of Swine’s Digestive Tract Microbiota and Its Relation to Production Indices—A Review. Animals 2020, 10, 527. [Google Scholar] [CrossRef]
- Iino, T.; Mori, K.; Tanaka, K.; Suzuki, K.-I.; Harayama, S. Oscillibacter valericigenes gen. nov., sp. nov., a valerate-producing anaerobic bacterium isolated from the alimentary canal of a Japanese corbicula clam. Int. J. Syst. Evol. Microbiol. 2007, 57, 1840–1845. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Na Kang, B.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Ramayo-Caldas, Y.; Mach, N.; Lepage, P.; Levenez, F.; Denis, C.; Lemonnier, G.; Leplat, J.-J.; Billon, Y.; Berri, M.; Doré, J.; et al. Phylogenetic network analysis applied to pig gut microbiota identifies an ecosystem structure linked with growth traits. ISME J. 2016, 10, 2973–2977. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Li, P.; Hao, S.; Zhang, Y.; Kim, S.W.; Li, H.; Ma, X.; Gao, S.; He, L.; Wu, W.; et al. Dynamic Distribution of the Gut Microbiota and the Relationship with Apparent Crude Fiber Digestibility and Growth Stages in Pigs. Sci. Rep. 2015, 5, 9938. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Kong, F.; Xiang, Y.; Zhou, W.; Wang, J.; Yang, H.; Zhang, G.; Zhao, J. Comparative biogeography of the gut microbiome between Jinhua and Landrace pigs. Sci. Rep. 2018, 8, 5985. [Google Scholar] [CrossRef] [PubMed]
- Scheppach, W.; Weiler, F. The butyrate story: Old wine in new bottles? Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Montagne, L.; Pluske, J.; Hampson, D. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed. Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- May, T.; Mackie, R.I.; Fahey, G.C.; Cremin, J.C.; Garleb, K.A. Effect of Fiber Source on Short-Chain Fatty Acid Production and on the Growth and Toxin Production by Clostridium difficile. Scand. J. Gastroenterol. 1994, 29, 916–922. [Google Scholar] [CrossRef]
- Wang, X.; Gibson, G.R. Effects of thein vitrofermentation of oligofructose and inulin by bacteria growing in the human large intestine. J. Appl. Bacteriol. 1993, 75, 373–380. [Google Scholar] [CrossRef]
- Vigors, S.; O’Doherty, J.V.; Kelly, A.K.; O’Shea, C.J.; Sweeney, T. The Effect of Divergence in Feed Efficiency on the Intestinal Microbiota and the Intestinal Immune Response in Both Unchallenged and Lipopolysaccharide Challenged Ileal and Colonic Explants. PLoS ONE 2016, 11, e0148145. [Google Scholar] [CrossRef]
- Flint, H.J.; Bayer, E.A. Plant Cell Wall Breakdown by Anaerobic Microorganisms from the Mammalian Digestive Tract. Ann. N. Y. Acad. Sci. 2008, 1125, 280–288. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items 1 | Growing Period | Finishing Period | ||||||
|---|---|---|---|---|---|---|---|---|
| CON | GBR50 | GBR75 | GBR100 | CON | GBR50 | GBR75 | GBR100 | |
| Ingredient (%) | ||||||||
| Corn | 63.59 | 31.80 | 15.90 | - | 70.80 | 35.40 | 17.70 | - |
| Ground brown rice | - | 31.80 | 47.69 | 63.59 | - | 35.40 | 53.10 | 70.80 |
| Soybean meal (44%) | 32.30 | 32.30 | 32.30 | 32.3 | 25.00 | 25.00 | 25.00 | 25.00 |
| Soybean oil | 1.50 | 1.50 | 1.50 | 1.50 | 2.00 | 2.00 | 2.00 | 2.00 |
| Limestone | 0.90 | 0.90 | 0.90 | 0.90 | 1.00 | 1.00 | 1.00 | 1.00 |
| Dicalcium phosphate | 1.00 | 1.00 | 1.00 | 1.00 | - | - | - | - |
| Iodized salt | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| Vitamin-Mineral premix 2 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 |
| L-lysine-HCl | 0.10 | 0.10 | 0.10 | 0.10 | - | - | - | - |
| DL-methionine | 0.01 | 0.01 | 0.01 | 0.01 | - | - | - | - |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Analyzed energy and nutrients | ||||||||
| DM, % | 96.36 | 96.47 | 96.62 | 96.38 | 96.72 | 96.41 | 96.56 | 96.52 |
| CP, % | 20.41 | 20.01 | 20.64 | 19.43 | 17.59 | 17.05 | 18.54 | 16.78 |
| GE, kcal/kg | 4436 | 4351 | 4446 | 4325 | 4325 | 4486 | 4275 | 4316 |
| Calculated energy and nutrients | ||||||||
| ME, kcal/kg | 3368 | 3438 | 3473 | 3508 | 3399 | 3477 | 3516 | 3555 |
| CP, % | 19.52 | 19.69 | 19.77 | 19.85 | 16.81 | 16.99 | 17.09 | 17.18 |
| Calcium, % | 0.68 | 0.68 | 0.68 | 0.68 | 0.59 | 0.59 | 0.59 | 0.59 |
| Phosphorous, % | 0.56 | 0.58 | 0.59 | 0.6 | 0.47 | 0.49 | 0.5 | 0.51 |
| Total lysine, % | 1.13 | 1.15 | 1.16 | 1.17 | 0.87 | 0.89 | 0.9 | 0.92 |
| Total methionine, % | 0.32 | 0.32 | 0.33 | 0.33 | 0.28 | 0.28 | 0.29 | 0.29 |
| Total methionine + cysteine, % | 0.66 | 0.66 | 0.65 | 0.65 | 0.58 | 0.58 | 0.58 | 0.57 |
| Total threonine, % | 0.75 | 0.76 | 0.76 | 0.77 | 0.64 | 0.65 | 0.65 | 0.66 |
| Total tryptophan, % | 0.23 | 0.27 | 0.29 | 0.31 | 0.19 | 0.23 | 0.25 | 0.27 |
| Items 2 | CON | GBR50 | GBR75 | GBR100 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Growing period (6 weeks) | ||||||
| Initial BW, kg | 23.79 | 24.07 | 24.08 | 23.98 | 1.490 | 0.982 |
| Final BW, kg | 63.15 | 65.06 | 64.20 | 63.54 | 1.875 | 0.946 |
| ADG, g/d | 937 | 976 | 955 | 941 | 21.35 | 0.702 |
| ADFI, g/d | 2073 | 2116 | 2084 | 2043 | 78.45 | 0.812 |
| G:F, g/g | 0.452 | 0.461 | 0.458 | 0.461 | 0.007 | 0.438 |
| Finishing period (6 weeks) | ||||||
| Initial BW, kg | 64.38 | 66.73 | 64.19 | 65.67 | 2.040 | 0.797 |
| Final BW, kg | 103.61 | 105.57 | 102.73 | 105.07 | 2.160 | 0.779 |
| ADG, g/d | 934 | 925 | 918 | 938 | 21.53 | 0.905 |
| ADFI, g/d | 3190 | 3168 | 3062 | 3185 | 96.39 | 0.761 |
| G:F, g/g | 0.293 | 0.292 | 0.300 | 0.295 | 0.008 | 0.916 |
| Items 2 | CON | GBR50 | GBR75 | GBR100 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Growing pig | ||||||
| DM, % | 87.88 | 89.43 | 89.05 | 90.06 | 1.571 | 0.446 |
| Energy, % | 85.89 | 85.90 | 86.56 | 87.19 | 2.134 | 0.635 |
| CP, % | 86.44 | 85.77 | 87.63 | 87.19 | 2.179 | 0.274 |
| Finishing pig | ||||||
| DM, % | 79.77 | 80.80 | 80.73 | 82.58 | 3.110 | 0.897 |
| Energy, % | 77.98 | 79.75 | 78.32 | 80.95 | 3.400 | 0.882 |
| CP, % | 73.47 | 70.98 | 77.78 | 77.31 | 3.940 | 0.519 |
| Items 2 | CON | GBR50 | GBR75 | GBR100 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Live weight, kg | 111.5 | 114.12 | 114.22 | 114.14 | 1.008 | 0.188 |
| HCW, kg | 87.74 | 89.78 | 89.87 | 89.8 | 0.52 | 0.289 |
| Dressing percentage, % | 78.69 | 78.68 | 78.69 | 78.67 | 1.16 | 0.914 |
| Back fat thickness, mm | 20.23 | 21.35 | 19.65 | 21.49 | 0.740 | 0.869 |
| Diversity Index 2 | CON | GBR100 | p-Value |
|---|---|---|---|
| Growing pigs | |||
| Average no. of sequence reads per sample | 181,852 ± 19,196 | 186,395 ± 16,116 | 0.769 |
| Observed OTUs | 259.00 ± 13.75 | 243.67 ± 3.79 | 0.136 |
| Chao1 | 276.83 ± 18.03 | 267.06 ± 11.10 | 0.469 |
| Shannon | 4.60 ± 0.44 | 4.62 ± 0.54 | 0.954 |
| Simpson | 0.89 ± 0.03 | 0.89 ± 0.03 | 0.966 |
| Finishing pigs | |||
| Average no. of sequence reads per sample | 217,143 ± 73,819 | 203,195 ± 46,636 | 0.796 |
| Observed OTUs | 330.33 ± 18.15 | 332.67 ± 17.62 | 0.881 |
| Chao1 | 351.12 ± 23.77 | 351.76 ± 18.60 | 0.972 |
| Shannon | 4.99 ± 0.32 | 5.15 ± 0.26 | 0.524 |
| Simpson | 0.92 ± 0.01 | 0.93 ± 0.01 | 0.380 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Cho, J.H.; Kim, Y.; Kim, H.B.; Song, M. Effects of Substitution of Corn with Ground Brown Rice on Growth Performance, Nutrient Digestibility, and Gut Microbiota of Growing-Finishing Pigs. Animals 2021, 11, 375. https://doi.org/10.3390/ani11020375
Kim S, Cho JH, Kim Y, Kim HB, Song M. Effects of Substitution of Corn with Ground Brown Rice on Growth Performance, Nutrient Digestibility, and Gut Microbiota of Growing-Finishing Pigs. Animals. 2021; 11(2):375. https://doi.org/10.3390/ani11020375
Chicago/Turabian StyleKim, Sheena, Jin Ho Cho, Younghoon Kim, Hyeun Bum Kim, and Minho Song. 2021. "Effects of Substitution of Corn with Ground Brown Rice on Growth Performance, Nutrient Digestibility, and Gut Microbiota of Growing-Finishing Pigs" Animals 11, no. 2: 375. https://doi.org/10.3390/ani11020375
APA StyleKim, S., Cho, J. H., Kim, Y., Kim, H. B., & Song, M. (2021). Effects of Substitution of Corn with Ground Brown Rice on Growth Performance, Nutrient Digestibility, and Gut Microbiota of Growing-Finishing Pigs. Animals, 11(2), 375. https://doi.org/10.3390/ani11020375