
Characterization and Functional Importance of Two Glycoside Hydrolase Family 16 Genes from the Rice White Tip Nematode Aphelenchoides besseyi




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Nematode Preparation
2.2. AbGH16 Gene Discovery and Cloning
2.3. Sequence and Phylogenetic Analysis
2.4. In Situ Hybridization
2.5. Differential Expression in Developmental Stages and Feeding Progress
2.6. Heterologous Expression of AbGH16 Proteins
2.7. Antifungal Activity Assays
2.8. RNA Interference
3. Results
3.1. Identification of the Novel GH16 Gene in the A. besseyi
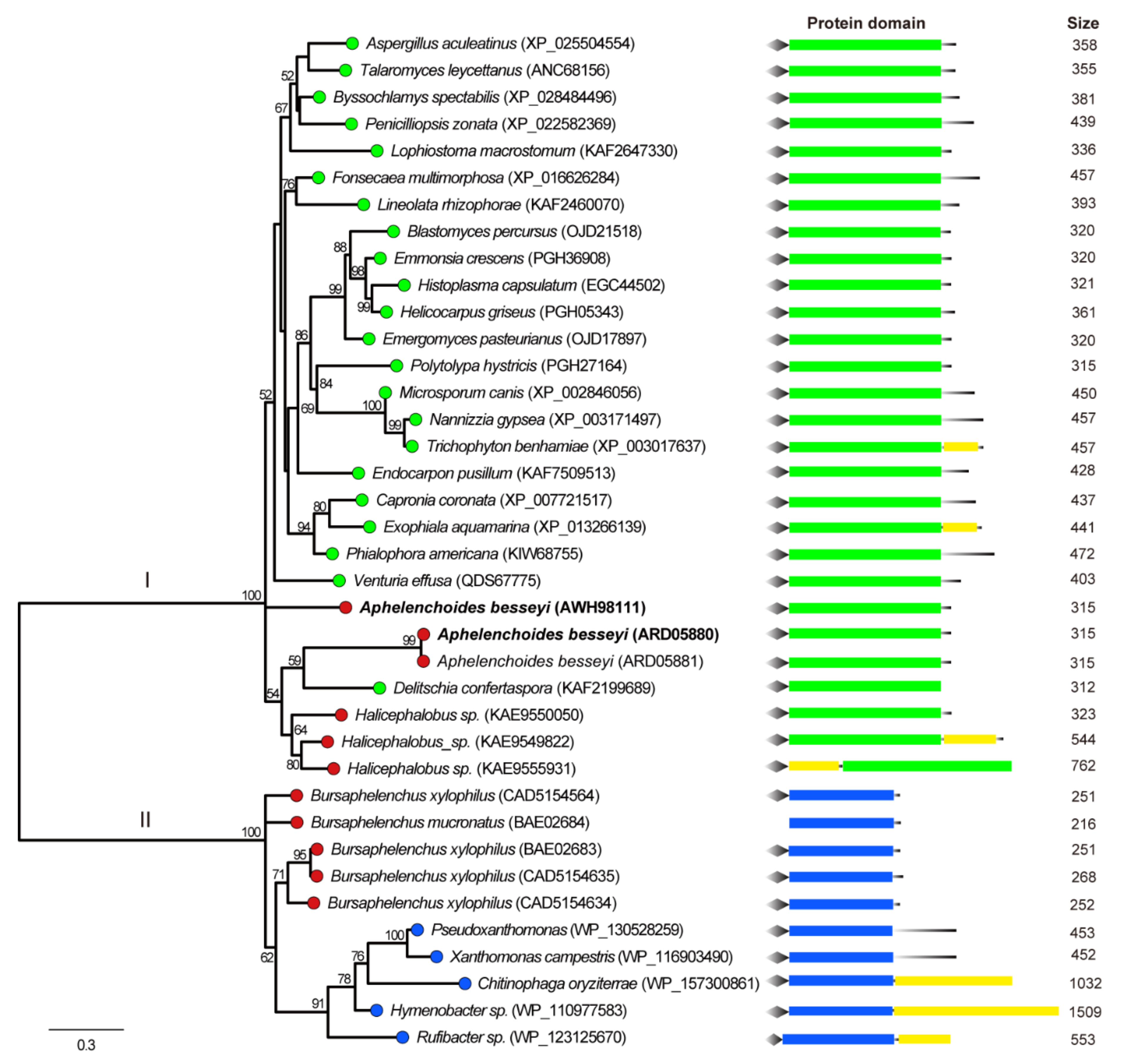
3.2. Phylogenetic and Structural Analysis
3.3. In Situ Hybridization and Expression Analysis
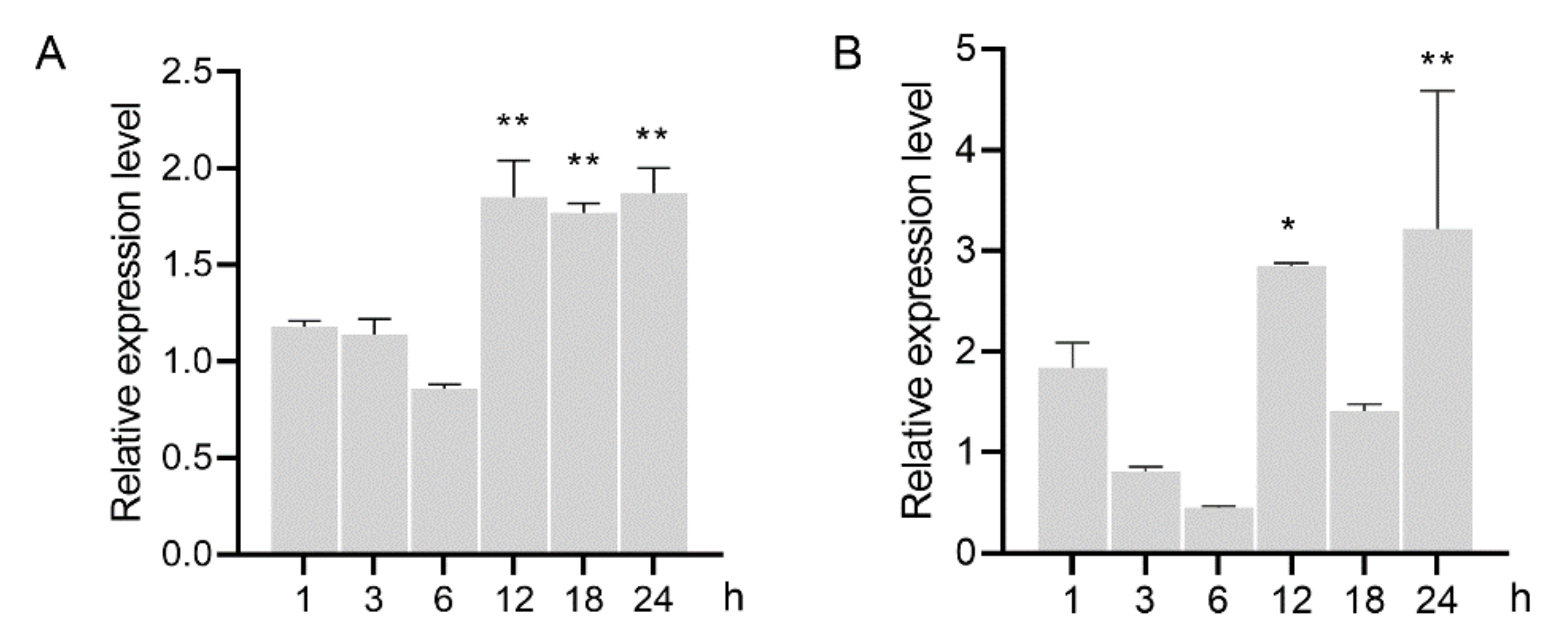
3.4. GH16 Expression was Induced in A. besseyi Feeding on the Fungus
3.5. In Vitro Antifungal Activity of Purified Recombinant AbGH16s
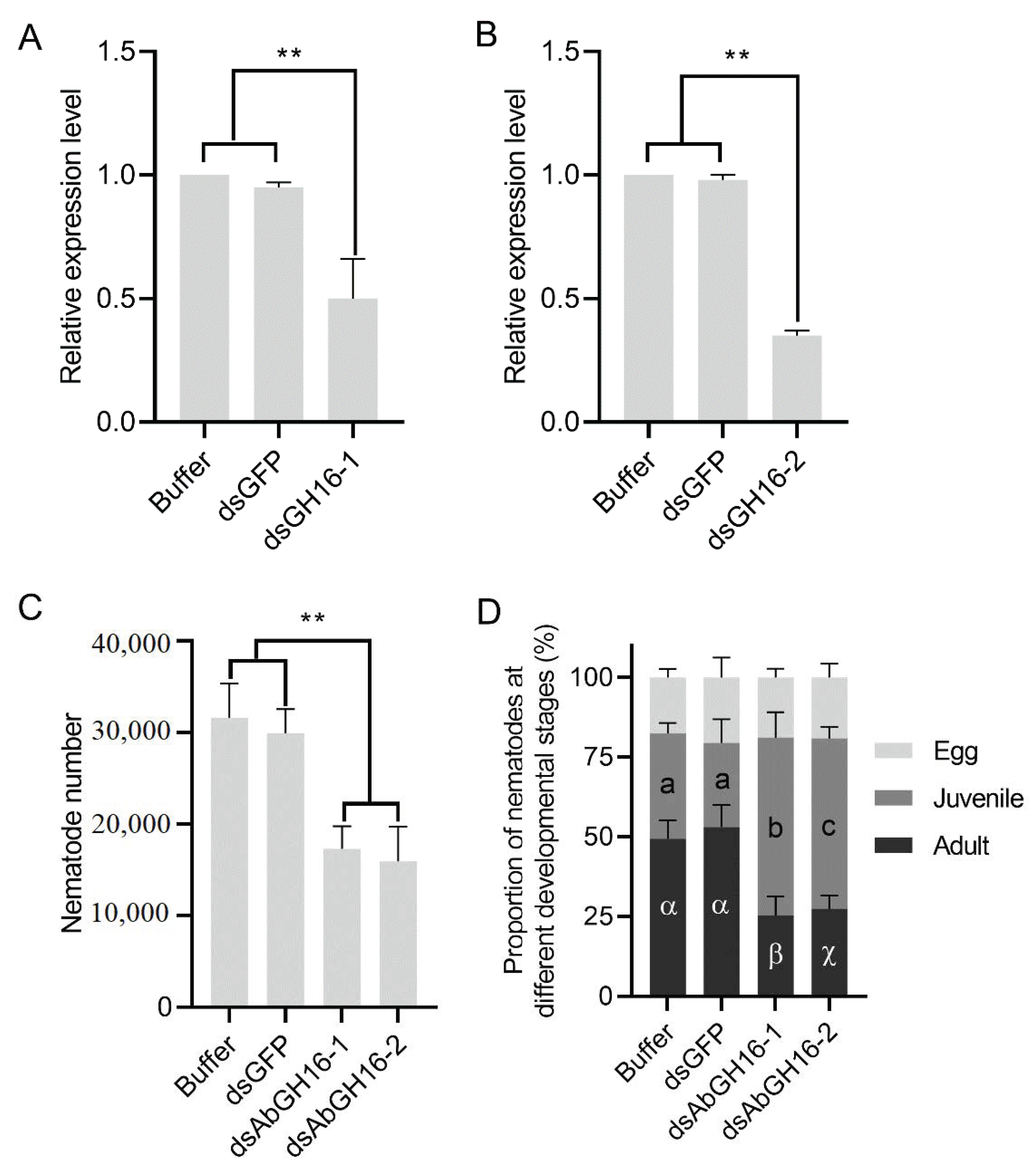
3.6. Decreased Reproduction Ability in the GH16 Knockdown Nematodes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- OEPP/EPPO. Aphelenchoides besseyi. Bull. OEPP/EPPO Bull. 2004, 34, 303–308. [Google Scholar] [CrossRef]
- Lin, M.S.; Ding, X.F.; Wang, Z.M.; Zhou, F.M.; Lin, N. Description of Aphelenchoides besseyi from abnormal rice with ‘Small Grains and Erect Panicles’ symptom in China. Rice Sci. 2004, 12, 289–294. [Google Scholar]
- Jones, O.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-LoPez, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Handoo, Z.A.; Rao, U.; Rao, S.B.; Prasad, J.S. Observations on the foliar nematode, Aphelenchoides besseyi, infecting tuberose and rce in India. J. Nematol. 2012, 44, 391–398. [Google Scholar] [PubMed]
- Wu, G.L.; Kuo, T.H.; Tsay, T.T.; Tsai, I.J.; Chen, P.J. Glycoside hydrolase (GH) 45 and 5 candidate cellulases in Aphelenchoides besseyi isolated from bird’s-nest fern. PLoS ONE 2016, 11, e0158663. [Google Scholar] [CrossRef]
- Sharma, S.B. Plant parasitic nematodes in rice-wheat based cropping systems in South Asia: Present status and future research thrust. J. Crop Prod. 2001, 4, 227–247. [Google Scholar] [CrossRef]
- Jagadiswari, R.; Kauraw, L.P.; Anand, P. Deterioration of seed quality due to interaction between Aphelenchoides besseyi and Curvularia lunata in rice. Ind. Phytopathol. 1994, 47, 92–93. [Google Scholar]
- Ruess, L.; Zapata, E.J.G.; Dighton, J. Food preferences of a fungal-feeding Aphelenchoides species. Nematology 2000, 2, 223–230. [Google Scholar] [CrossRef]
- Jamali, S.; Pourjam, E.; Alizadeh, A.; Alinia, F. Reproduction of the white tip nematode in different monoxenic cultures. J. Agric. Sci. Technol. 2008, 10, 165–171. [Google Scholar]
- Yoshida, K.; Hasegawa, K.; Mochiji, N.; Miwa, J. Early embryogenesis and anterior-posterior axis formation in the white-tip nematode Aphelenchoides besseyi (Nematoda: Aphelenchoididae). J. Nematol. 2009, 41, 17–22. [Google Scholar] [PubMed]
- Rosso, M.N.; Favery, B.; Piotte, C.; Arthaud, L.; Boer, J.M.D.; Hussey, R.S.; Bakker, J.; Baum, T.J.; Abad, P. Isolation of a cDNA encoding a β-1,4-endoglucanase in the root-knot nematode Meloidogyne incognita and expression analysis during plant parasitism. Mol. Plant Microbe Interact. 1999, 12, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Smant, G.; Stokkermans, J.P.W.G.; Yan, Y.; Boer, J.M.D.; Baum, T.J.; Wang, X.; Hussey, R.S.; Gommers, F.J.; Henrissat, B.; Davis, E.L.; et al. Endogenous cellulases in animals: Isolation of β-1,4-endoglucanase genes from two species of plant-parasitic cyst nematodes. Proc. Natl. Acad. Sci. USA 1998, 95, 4906–4911. [Google Scholar] [CrossRef] [PubMed]
- Popeijus, H.; Overmars, H.; Jones, J.; Blok, V.; Goverse, A.; Helder, J.; Schots, A.; Bakker, J.; Smant, G. Degradation of plant cell walls by a nematode. Nature 2000, 406, 36–37. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, B.; Haegeman, A.; Jacob, J.; Cannoot, B.; Gheysen, G. Arabinogalactan endo-1,4-β-galactosidase: A putative plant cell wall-degrading enzyme of plant-parasitic nematodes. Nematology 2009, 11, 739–747. [Google Scholar] [CrossRef]
- Qin, L.; Kudla, U.; Roze, E.H.; Goverse, A.; Popeijus, H.; Nieuwland, J.; Overmars, H.; Jones, J.T.; Schots, A.; Smant, G.; et al. Plant degradation: A nematode expansin acting on plants. Nature 2004, 427, 30. [Google Scholar] [CrossRef]
- Liu, J.; Peng, H.; Cui, J.; Huang, W.; Kong, L.; Clarke, J.L.; Jian, H.; Wang, G.L.; Peng, D. Molecular characterization of a novel effector expansin-like protein from Heterodera avenae that induces cell death in Nicotiana benthamiana. Sci. Rep. 2016, 6, 35677. [Google Scholar] [CrossRef]
- Peng, H.; Peng, D.; Long, H.; He, W.; Qiao, F.; Wang, G.; Huang, W. Characterisation and functional importance of β-1,4-endoglucanases from the potato rot nematode, Ditylenchus destructor. Nematology 2014, 16, 505–517. [Google Scholar] [CrossRef]
- Kikuchi, T.; Cotton, J.A.; Dalzell, J.J.; Hasegawa, K.; Kanzaki, N.; McVeigh, P.; Takanashi, T.; Tsai, I.J.; Assefa, S.A.; Cock, P.J.; et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011, 7, e1002219. [Google Scholar] [CrossRef]
- Ju, Y.; Wang, X.; Guan, T.; Peng, D.; Li, H. Versatile glycoside hydrolase family 18 chitinases for fungi ingestion and reproduction in the pinewood nematode Bursaphelenchus xylophilus. Int. J. Parasitol. 2016, 46, 819–828. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Hirooka, Y.; Tsai, I.J.; Masuya, H.; Hino, A.; Kanzaki, N.; Jones, J.T.; Kikuchi, T. Distribution and evolution of glycoside hydrolase family 45 cellulases in nematodes and fungi. BMC Evol. Biol. 2014, 14, 69. [Google Scholar] [CrossRef]
- Kikuchi, T.; Cock, P.J.A.; Helder, J.; Jones, J.T. Characterisation of the transcriptome of Aphelenchoides besseyi and identification of a GHF 45 cellulase. Nematology 2014, 16, 99–107. [Google Scholar] [CrossRef]
- Wang, F.; Li, D.; Wang, Z.; Dong, A.; Liu, L.; Wang, B.; Chen, Q.; Liu, X. Transcriptomic analysis of the rice white tip Nematode, Aphelenchoides besseyi (Nematoda: Aphelenchoididae). PLoS ONE 2014, 9, e91591. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Xu, C.L.; Yang, S.H.; Li, J.Y.; Wang, H.L.; Zhang, Z.X.; Chen, C.; Xie, H. Life-stage specific transcriptomes of a migratory endoparasitic plant nematode, Radopholus similis elucidate a different parasitic and life strategy of plant parasitic nematodes. Sci. Rep. 2019, 9, 6277. [Google Scholar] [CrossRef] [PubMed]
- Viborg, A.H.; Terrapon, N.; Lombard, V.; Michel, G.; Czjzek, M.; Henrissat, B.; Brumer, H. A subfamily roadmap of the evolutionarily diverse glycoside hydrolase family 16 (GH16). J. Biol. Chem. 2019, 294, 15973–15986. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
- Mao, S.; Gao, P.; Lu, P.; Lu, Z.; Zhang, F.; Zhao, C.; Bie, X. Purification, characterization, and heterologous expression of a thermostable β-1,3-1,4-glucanase from Bacillus altitudinis YC-9. Appl. Biochem. Biotechnol. 2013, 169, 960–975. [Google Scholar] [CrossRef]
- Li, J.; Liu, W.; Luo, L.; Dong, D.; Liu, T.; Zhang, T.; Lu, C.; Liu, D.; Zhang, D.; Wu, H. Expression of Paenibacillus polymyxa β-1,3-1,4-glucanase in Streptomyces lydicus A01 improves its biocontrol effect against Botrytis cinerea. Biol. Control 2015, 90, 141–147. [Google Scholar] [CrossRef]
- Anderson, V.A.; Haley, S.D.; Peairs, F.B.; van Eck, L.; Leach, J.E.; Lapitan, N.L. Virus-induced gene silencing suggests (1,3;1,4)-beta-glucanase is a susceptibility factor in the compatible Russian wheat aphid-wheat interaction. Mol. Plant Microbe Interact. 2014, 27, 913–922. [Google Scholar] [CrossRef]
- Linton, S.M. Review: The structure and function of cellulase (endo-beta-1,4-glucanase) and hemicellulase (beta-1,3-glucanase and endo-beta-1,4-mannase) enzymes in invertebrates that consume materials ranging from microbes, algae to leaf litter. Comp. Biochem. Physiol. B 2020, 240, 110354. [Google Scholar] [CrossRef]
- Waldron, R.; McGowan, J.; Gordon, N.; Mitchell, E.B.; Fitzpatrick, D.A.; Doyle, S. Characterisation of three novel beta-1,3 glucanases from the medically important house dust mite Dermatophagoides pteronyssinus (airmid). Insect Biochem. Mol. Biol. 2019, 115, 103242. [Google Scholar] [CrossRef]
- Souza, R.S.; Gama, M.d.V.F.; Schama, R.; Lima, J.B.P.; Diaz-Albiter, H.M.; Genta, F.A. Biochemical and functional characterization of glycoside hydrolase family 16 genes in Aedes aegypti larvae: Identification of the major digestive β-1,3-glucanase. Front. Physiol. 2019, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, C.; Jiang, Y.; Huang, Q.; Shen, Y.; Ni, J. A Novel Digestive GH16 beta-1,3(4)-Glucanase from the fungus-growing termite Macrotermes barneyi. Appl. Biochem. Biotechnol. 2020, 192, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Shibuya, H.; Jones, J.T. Molecular and biochemical characterization of an endo-β-1,3-glucanase from the pinewood nematode Bursaphelenchus xylophilus acquired by horizontal gene transfer from bacteria. Biochem. J. 2005, 389, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Wei, L.; Lin, M.; Zhou, Y. Assessment of rice cultivars in China for field resistance to Aphelenchoides besseyi. J. Integr. Agric. 2014, 13, 2221–2228. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- de Boer, J.M.; Yan, Y.; Smant, G.; Davis, E.L.; Baum, T.J. In-situ hybridization to messenger RNA in Heterodera glycines. J. Nematol. 1998, 30, 309–312. [Google Scholar]
- Michel, G.; Chantalat, L.; Duee, E.; Barbeyron, T.; Henrissat, B.; Kloareg, B.; Dideberg, O. The κ-carrageenase of P. carrageenovora features a tunnel-shaped active site: A novel insight in the evolution of Clan-B glycoside hydrolases. Structure 2001, 9, 513–525. [Google Scholar] [CrossRef]
- Ragsdale, E.J.; Koutsovoulos, G.; Biddle, J.F. A draft genome for a species of Halicephalobus (Panagrolaimidae). J. Nematol. 2019, 51, 1–4. [Google Scholar] [CrossRef]
- Henrissat, B.; Bairoch, A. New families in the classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1993, 293, 781–788. [Google Scholar] [CrossRef]
- Danchin, E.G.; Rosso, M.N.; Vieira, P.; de Almeida-Engler, J.; Coutinho, P.M.; Henrissat, B.; Abad, P. Multiple lateral gene transfers and duplications have promoted plant parasitism ability in nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 17651–17656. [Google Scholar] [CrossRef]
- Wieczorek, K.; Elashry, A.; Quentin, M.; Grundler, F.M.; Favery, B.; Seifert, G.J.; Bohlmann, H. A distinct role of pectate lyases in the formation of feeding structures induced by cyst and root-knot nematodes. Mol. Plant Microbe Interact. 2014, 27, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Danchin, E.G.J.; Perfus-Barbeoch, L.; Rancurel, C.; Thorpe, P.; Da Rocha, M.; Bajew, S.; Neilson, R.; Guzeeva, E.S.; Da Silva, C.; Guy, J.; et al. The transcriptomes of Xiphinema index and Longidorus elongatus suggest independent acquisition of some plant parasitism genes by horizontal gene transfer in early-branching nematodes. Genes 2017, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The role of the fungal cell wall in the infection of plants. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Jijakli, M.H.; Lepoivre, P. Characterization of an exo-β-1,3-glucanase produced by Pichia anomala strain k, antagonist of Botrytis cinerea on apples. Phytopathology 1998, 88, 335–343. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, H.; Zhou, D.; Daly, P.; Wang, X.; Wei, L. Characterization and Functional Importance of Two Glycoside Hydrolase Family 16 Genes from the Rice White Tip Nematode Aphelenchoides besseyi. Animals 2021, 11, 374. https://doi.org/10.3390/ani11020374
Feng H, Zhou D, Daly P, Wang X, Wei L. Characterization and Functional Importance of Two Glycoside Hydrolase Family 16 Genes from the Rice White Tip Nematode Aphelenchoides besseyi. Animals. 2021; 11(2):374. https://doi.org/10.3390/ani11020374
Chicago/Turabian StyleFeng, Hui, Dongmei Zhou, Paul Daly, Xiaoyu Wang, and Lihui Wei. 2021. "Characterization and Functional Importance of Two Glycoside Hydrolase Family 16 Genes from the Rice White Tip Nematode Aphelenchoides besseyi" Animals 11, no. 2: 374. https://doi.org/10.3390/ani11020374
APA StyleFeng, H., Zhou, D., Daly, P., Wang, X., & Wei, L. (2021). Characterization and Functional Importance of Two Glycoside Hydrolase Family 16 Genes from the Rice White Tip Nematode Aphelenchoides besseyi. Animals, 11(2), 374. https://doi.org/10.3390/ani11020374