
Effects of β-Mannanase and Bacteriophage Supplementation on Health and Growth Performance of Holstein Calves

 , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design and Diets
2.3. Enumeration of Intestinal Pathogenic Bacteria
2.4. Hematological Parameters
2.5. Chemical Analysis
2.6. Statistical Analysis
3. Results
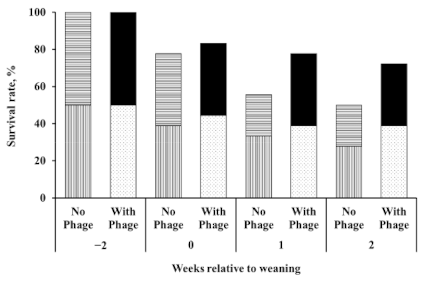
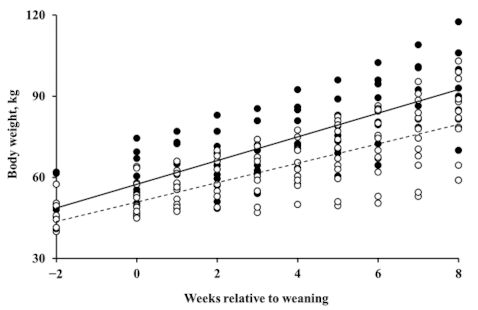
3.1. Effects of Supplementation of Bacteriophage
3.2. Effects of Supplementation of β-Mannanase
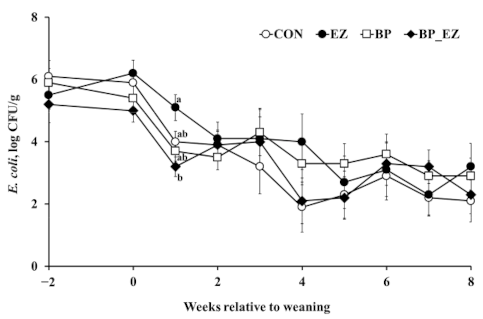
3.3. Intestinal Pathogenic Bacteria and Hematological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Svensson, C.; Linder, A.; Olsson, S.-O. Mortality in Swedish dairy calves and replacement heifers. J. Dairy Sci. 2006, 89, 4769–4777. [Google Scholar] [CrossRef]
- Taylor, M. Protozoal disease in cattle and sheep. In Pract. 2000, 22, 604–617. [Google Scholar] [CrossRef]
- Cho, Y.-I.; Yoon, K.-J. An overview of calf diarrhea-infectious etiology, diagnosis, and intervention. J. Vet. Sci. 2014, 15, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Uetake, K. Newborn calf welfare: A review focusing on mortality rates. Anim. Sci. J. 2013, 84, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Lago, A.; McGuirk, S.M.; Bennett, T.B.; Cook, N.B.; Nordlund, K.V. Calf respiratory disease and pen microenvironments in naturally ventilated calf barns in winter. J. Dairy Sci. 2006, 89, 4014–4025. [Google Scholar] [CrossRef]
- Chattopadhyay, M.K. Use of antibiotics as feed additives: A burning question. Front. Microbiol. 2014, 5, 334–336. [Google Scholar] [CrossRef]
- Gebru, E.; Lee, J.S.; Son, J.C.; Yang, S.Y.; Shin, S.A.; Kim, B.; Kim, M.K.; Park, S.C. Effect of probiotic-, bacteriophage-, or organic acid-supplemented feeds or fermented soybean meal on the growth performance, acute-phase response, and bacterial shedding of grower pigs challenged with serotype Typhimurium. J. Anim. Sci. 2010, 88, 3880–3886. [Google Scholar] [CrossRef]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef]
- Brüssow, H. Phage therapy: The Escherichia coli experience. Microbiology 2005, 151, 2133–2140. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, J.W.; Shin, H.S.; Kim, M.C.; Lee, J.H.; Kim, G.B.; Kil, D.Y. Effect of dietary supplementation of bacteriophage on performance, egg quality and caecal bacterial populations in laying hens. Br. Poult. Sci. 2015, 56, 132–136. [Google Scholar] [CrossRef]
- Kim, J.S.; Hosseindoust, A.; Lee, S.H.; Choi, Y.H.; Kim, M.J.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage cocktail and multi-strain probiotics in the feed for weanling pigs: Effects on intestine morphology and targeted intestinal coliforms and Clostridium. Animal 2017, 11, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Castillo, M.; Martin-Orue, S.; Taylor-Pickard, J.; Perez, J.; Gasa, J. Use of mannanoligosaccharides and zinc chelate as growth promoters and diarrhea preventative in weaning pigs: Effects on microbiota and gut function. J. Anim. Sci. 2008, 86, 94–101. [Google Scholar] [CrossRef]
- Hume, M.E. Food safety symposium: Potential impact of reduced antibiotic use and the roles of prebiotics, probiotics, and other alternatives in antibiotic-free broiler production. Poult. Sci. 2011, 90, 2663–2669. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.E.; Geronian, K.; Knox, A.; McNab, J.; McCartney, E. A dose-response study with the feed enzyme beta-mannanase in broilers provided with corn-soybean meal based diets in the absence of antibiotic growth promoters. Poult. Sci. 2004, 83, 1992–1996. [Google Scholar] [CrossRef]
- Cho, J.H.; Kim, I.H. Effects of beta-mannanase supplementation in combination with low and high energy dense diets for growing and finishing broilers. Livest. Sci. 2013, 154, 137–143. [Google Scholar] [CrossRef]
- Yoon, S.Y.; Yang, Y.X.; Shinde, P.L.; Choi, J.Y.; Kim, J.S.; Kim, Y.W.; Yun, K.; Jo, J.K.; Lee, J.H.; Ohh, S.J.; et al. Effects of mannanase and distillers dried grain with solubles on growth performance, nutrient digestibility, and carcass characteristics of grower-finisher pigs. J. Anim. Sci. 2010, 88, 181–191. [Google Scholar] [CrossRef]
- Kim, J.; Ingale, S.; Lee, S.; Kim, K.; Lee, J.; Chae, B. Effects of energy levels of diet and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites in growing pigs. Anim. Feed Sci. Technol. 2013, 186, 64–70. [Google Scholar] [CrossRef]
- Lee, J.-J.; Seo, J.; Jung, J.K.; Lee, J.; Lee, J.-H.; Seo, S. Effects of β-mannanase supplementation on growth performance, nutrient digestibility, and nitrogen utilization of Korean native goat (Capra hircus coreanae). Livest. Sci. 2014, 169, 83–87. [Google Scholar] [CrossRef]
- Tewoldebrhan, T.; Appuhamy, J.; Lee, J.-J.; Niu, M.; Seo, S.; Jeong, S.; Kebreab, E. Exogenous β-mannanase improves feed conversion efficiency and reduces somatic cell count in dairy cattle. J. Dairy Sci. 2017, 100, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-Y.; Lee, S.-M.; Kim, J.-H.; Ki, K.-S.; Kim, H.-S.; Kam, D.-K.; Lee, J.-H.; Lee, J.-J.; Bae, G.-S.; Seo, S.-W. Effect of β-mannanase (CTCZYME®) on the growth of young calf. Korean J. Agric. Sci. 2010, 37, 239–243. [Google Scholar] [CrossRef]
- Wang, T.; Lee, K.-H.; Jung, U.-S.; Jin, Y.-C.; Lee, S.-B.; Lee, J.-S.; Hwang, J.-H.; Lim, J.-N.; Kim, M.-J.; Choi, S.-H. Responses of blood metabolites and proteins to different vitamin A levels in Korean native steers. Pak. Vet. J. 2014, 34, 527–531. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Jeon, S.; Ha, J.K. Guidelines for experimental design and statistical analyses in animal studies submitted for publication in the Asian-Australasian Journal of Animal Sciences. Asian-Australas. J. Anim. Sci. 2018, 31, 1381–1386. [Google Scholar] [CrossRef] [PubMed]
- Torsein, M.; Lindberg, A.; Sandgren, C.H.; Waller, K.P.; Törnquist, M.; Svensson, C. Risk factors for calf mortality in large Swedish dairy herds. Prev. Vet. Med. 2011, 99, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Wymann, M.N.; Bonfoh, B.; Schelling, E.; Bengaly, S.; Tembely, S.; Tanner, M.; Zinsstag, J. Calf mortality rate and causes of death under different herd management systems in peri-urban Bamako, Mali. Livest. Sci. 2006, 100, 169–178. [Google Scholar] [CrossRef]
- Wells, S.J.; Dargatz, D.A.; Ott, S.L. Factors associated with mortality to 21 days of life in dairy heifers in the United States. Prev. Vet. Med. 1996, 29, 9–19. [Google Scholar] [CrossRef]
- Godden, S. Colostrum management for dairy calves. Vet. Clin. N. Am. Food Anim. 2008, 24, 19–39. [Google Scholar] [CrossRef]
- Beam, A.; Lombard, J.; Kopral, C.; Garber, L.; Winter, A.; Hicks, J.; Schlater, J. Prevalence of failure of passive transfer of immunity in newborn heifer calves and associated management practices on US dairy operations. J. Dairy Sci. 2009, 92, 3973–3980. [Google Scholar] [CrossRef]
- Waldner, C.L.; Rosengren, L.B. Factors associated with serum immunoglobulin levels in beef calves from Alberta and Saskatchewan and association between passive transfer and health outcomes. Can. Vet. J. 2009, 50, 275–281. [Google Scholar] [CrossRef][Green Version]
- Dean-Nystrom, E.A.; Bosworth, B.T.; Cray, W.; Moon, H.W. Pathogenicity of Escherichia coli O157: H7 in the intestines of neonatal calves. Infect. Immun. 1997, 65, 1842–1848. [Google Scholar] [CrossRef]
- Callaway, T.R.; Edrington, T.S.; Brabban, A.D.; Keen, J.E.; Anderson, R.C.; Rossman, M.L.; Engler, M.J.; Genovese, K.J.; Gwartney, B.L.; Reagan, J.O.; et al. Fecal prevalence of Escherichia coli O157, Salmonella, Listeria, and Bacteriophage Infecting E. coli O157:H7 in feedlot cattle in the Southern Plains region of the United States. Foodborne Pathog. Dis. 2006, 3, 234–244. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Ross, R.P.; Coffey, A. Bacteriophage and their lysins for elimination of infectious bacteria. FEMS Microbiol. Rev. 2009, 33, 801–819. [Google Scholar] [CrossRef]
- Lim, T.-H.; Kim, M.-S.; Lee, D.-H.; Lee, Y.-N.; Park, J.-K.; Youn, H.-N.; Lee, H.-J.; Yang, S.-Y.; Cho, Y.-W.; Lee, J.-B.; et al. Use of bacteriophage for biological control of Salmonella Enteritidis infection in chicken. Res. Vet. Sci. 2012, 93, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Callaway, T.R.; Edrington, T.S.; Brabban, A.D.; Anderson, R.C.; Rossman, M.L.; Engler, M.J.; Carr, M.A.; Genovese, K.J.; Keen, J.E.; Looper, M.L.; et al. Bacteriophage isolated from feedlot cattle can reduce Escherichia coli O157:H7 populations in ruminant gastrointestinal tracts. Foodborne Pathog. Dis. 2008, 5, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Lau, G.; Sieo, C.; Tan, W.; Hair-Bejo, M.; Jalila, A.; Ho, Y. Efficacy of a bacteriophage isolated from chickens as a therapeutic agent for colibacillosis in broiler chickens. Poult. Sci. 2010, 89, 2589–2596. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.Y.; Baek, H.Y.; Kim, I.H. Effects of bacteriophage supplementation on egg performance, egg quality, excreta microflora, and moisture content in laying hens. Asian-Australas. J. Anim. Sci. 2012, 25, 1015–1020. [Google Scholar] [CrossRef]
- Wang, J.P.; Yan, L.; Lee, J.H.; Kim, I.H. Evaluation of bacteriophage supplementation on growth performance, blood characteristics, relative organ weight, breast muscle characteristics and excreta microbial shedding in broilers. Asian-Australas. J. Anim. Sci. 2013, 26, 573–578. [Google Scholar] [CrossRef]
- Kim, K.H.; Ingale, S.L.; Kim, J.S.; Lee, S.H.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage and probiotics both enhance the performance of growing pigs but bacteriophage are more effective. Anim. Feed Sci. Technol. 2014, 196, 88–95. [Google Scholar] [CrossRef]
- Hosseindoust, A.R.; Lee, S.H.; Kim, J.S.; Choi, Y.H.; Noh, H.S.; Lee, J.H.; Jha, P.K.; Kwon, I.K.; Chae, B.J. Dietary bacteriophages as an alternative for zinc oxide or organic acids to control diarrhoea and improve the performance of weanling piglets. Vet. Med. 2017, 62, 53–61. [Google Scholar] [CrossRef]
- Jones, M.L.; Allison, R.W. Evaluation of the ruminant complete blood cell count. Vet. Clin. N. Am. Food Anim. 2007, 23, 377–402. [Google Scholar] [CrossRef]
- Jo, N.C.; Jung, J.; Kim, J.N.; Lee, J.; Jeong, S.Y.; Kim, W.; Sung, H.G.; Seo, S. Effect of vaccination against foot-and-mouth disease on growth performance of Korean native goat (Capra hircus coreanae). J. Anim. Sci. 2014, 92, 2578–2586. [Google Scholar] [CrossRef] [PubMed]
- Sarıkaya, B.; Azkur, A.K.; Gazyagci, S. Inactivated bovine viral diarrhea virus vaccine trigger leucopenia and lymphopenia on calves. Acta Sci. Vet. 2011, 39, 994–999. [Google Scholar]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; Chen, Y.; Li, Z.; Cao, Y. Effects of β-mannanase expressed by Pichia pastoris in corn–soybean meal diets on broiler performance, nutrient digestibility, energy utilization and immunoglobulin levels. Anim. Feed Sci. Technol. 2010, 159, 59–67. [Google Scholar] [CrossRef]
- Lv, J.; Chen, Y.; Guo, X.; Piao, X.; Cao, Y.; Dong, B. Effects of supplementation of β-mannanase in corn-soybean meal diets on performance and nutrient digestibility in growing pigs. Asian-Australas. J. Anim. Sci. 2013, 26, 579. [Google Scholar] [CrossRef]
- Kim, S.C.; Kim, J.W.; Kim, J.U.; Kim, I.H. Effects of dietary supplementation of bacteriophage on growth performance, nutrient digestibility, blood profiles, carcass characteristics and fecal microflora in broilers. Korean J. Poult. Sci. 2013, 40, 75–81. [Google Scholar] [CrossRef][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Experimental Diets | ||||||
|---|---|---|---|---|---|---|
| Item 1 | CON 2 | EZ | BP | BP_EZ | Timothy | Milk Replacer |
| DM (g/kg as fed) | 889 | 889 | 888 | 886 | 928 | 952 |
| CP | 216 | 219 | 200 | 226 | 60 | 211 |
| EE | 67 | 64 | 58 | 61 | 12 | 140 |
| Ash | 56 | 65 | 59 | 62 | 37 | 104 |
| aNDF | 308 | 327 | 362 | 377 | 713 | - |
| ADF | 223 | 245 | 276 | 293 | 470 | - |
| p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Treatment 1 | Contrast | |||||||||
| Item 2 | n | CON | EZ | BP | BP_EZ | SEM 3 | Overall | EZ | BP | Interaction |
| BW, kg | ||||||||||
| Initial BW | 36 | 44 | 47 | 44 | 45 | 2.7 | 0.84 | 0.40 | 0.77 | 0.80 |
| BW at weaning | 29 | 52 | 55 | 50 | 56 | 3.4 | 0.56 | 0.18 | 0.81 | 0.61 |
| Final BW | 22 | 87 | 99 | 77 | 89 | 7.3 | 0.16 | 0.08 | 0.17 | 0.99 |
| ADG, g | 22 | 569 | 638 | 451 | 587 | 73.7 | 0.21 | 0.13 | 0.21 | 0.61 |
| Intake, g/d | ||||||||||
| DMI | 22 | 1612 | 1834 | 1402 | 1624 | 149.6 | 0.18 | 0.11 | 0.14 | 1.00 |
| Concentrate DMI | 22 | 802 | 993 | 759 | 921 | 71.0 | 0.06 | 0.01 | 0.46 | 0.82 |
| Forage DMI | 22 | 700 | 715 | 538 | 590 | 92.2 | 0.34 | 0.68 | 0.07 | 0.82 |
| Feed Efficiency 4 | 22 | 0.36 | 0.32 | 0.35 | 0.36 | 0.032 | 0.60 | 0.40 | 0.59 | 0.30 |
| p-Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments 1 | Contrast | ||||||||
| Item 2 | CON | EZ | BP | BP_EZ | SEM 3 | Overall | EZ | BP | Interaction |
| Leukocytes | |||||||||
| WBC, 103/μL | 7.6 | 8.7 | 7.7 | 9.2 | 0.53 | 0.66 | 0.75 | 0.76 | 0.24 |
| Lymphocytes, 103/μL | 3.2 | 3.1 | 3.2 | 2.9 | 0.16 | 0.07 | 0.66 | 0.94 | <0.01 |
| Monocytes, 103/μL | 0.75 | 0.93 | 0.74 | 0.76 | 0.111 | 0.28 | 0.22 | 0.26 | 0.29 |
| Neutrophils, 103/μL | 2.3 | 2.2 | 2.3 | 2.4 | 0.12 | 0.18 | 0.68 | 0.71 | 0.03 |
| Eosinophils, 103/μL | 0.44 | 0.47 | 0.46 | 0.57 | 0.068 | 0.27 | 0.19 | 0.64 | 0.16 |
| Basophils, 103/μL | 0.02 | 0.03 | 0.03 | 0.02 | 0.004 | 0.15 | 0.40 | 0.04 | 0.56 |
| N:L | 0.81 | 0.93 | 0.81 | 0.91 | 0.13 | 0.08 | 0.48 | 0.46 | 0.02 |
| Erythrocytes | |||||||||
| RBC, 106/μL | 7.1 | 7.3 | 7.5 | 6.8 | 0.31 | 0.53 | 0.66 | 0.31 | 0.35 |
| MCV, fL | 35.6 | 34.0 | 35.5 | 34.2 | 0.71 | 0.23 | 0.26 | 0.56 | 0.10 |
| HCT, % | 25.1 | 25.3 | 26.3 | 23.4 | 1.16 | 0.58 | 0.96 | 0.45 | 0.25 |
| MCH, pg | 12.0 | 11.8 | 12.0 | 11.9 | 0.16 | 0.85 | 0.49 | 0.57 | 1.00 |
| MCHC, g/dL | 34.2 | 34.8 | 34.5 | 35.7 | 0.51 | 0.78 | 0.88 | 0.72 | 0.34 |
| Hgb, g/dL | 9.3 | 8.5 | 8.8 | 8.2 | 0.59 | 0.41 | 0.30 | 0.59 | 0.22 |
| RDW, % | 29.0 | 19.3 | 20.5 | 21.3 | 4.58 | 1.00 | 0.87 | 0.98 | 0.94 |
| Thrombocytes | |||||||||
| PLT, 106/μL | 2.3 | 2.5 | 2.5 | 2.4 | 0.24 | 0.61 | 0.41 | 0.36 | 0.61 |
| MPV, fl | 16.2 | 16.8 | 16.7 | 16.7 | 0.37 | 0.67 | 0.83 | 0.48 | 0.33 |
| Total protein, mg/dL | 6.1 | 5.9 | 6.1 | 6.3 | 0.18 | 0.84 | 0.47 | 0.85 | 0.62 |
| BUN, mg/dL | 11.5 | 9.5 | 12.0 | 12.1 | 0.70 | 0.56 | 0.65 | 0.29 | 0.40 |
| Glucose, mg/dL | 68.7 | 74.3 | 64.1 | 63.1 | 3.51 | 0.72 | 0.29 | 0.63 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Jo, N.; Lee, J.-J.; Lee, J.-H.; Kam, D.-K.; Seo, J.; Kebreab, E.; Seo, S. Effects of β-Mannanase and Bacteriophage Supplementation on Health and Growth Performance of Holstein Calves. Animals 2021, 11, 372. https://doi.org/10.3390/ani11020372
Jeong S, Jo N, Lee J-J, Lee J-H, Kam D-K, Seo J, Kebreab E, Seo S. Effects of β-Mannanase and Bacteriophage Supplementation on Health and Growth Performance of Holstein Calves. Animals. 2021; 11(2):372. https://doi.org/10.3390/ani11020372
Chicago/Turabian StyleJeong, Sinyong, Namchul Jo, Jung-Jin Lee, Jae-Hwan Lee, Dong-Keun Kam, Jakyeom Seo, Ermias Kebreab, and Seongwon Seo. 2021. "Effects of β-Mannanase and Bacteriophage Supplementation on Health and Growth Performance of Holstein Calves" Animals 11, no. 2: 372. https://doi.org/10.3390/ani11020372
APA StyleJeong, S., Jo, N., Lee, J.-J., Lee, J.-H., Kam, D.-K., Seo, J., Kebreab, E., & Seo, S. (2021). Effects of β-Mannanase and Bacteriophage Supplementation on Health and Growth Performance of Holstein Calves. Animals, 11(2), 372. https://doi.org/10.3390/ani11020372