
Identification of Genes Related to Cold Tolerance and Novel Genetic Markers for Molecular Breeding in Taiwan Tilapia (Oreochromis spp.) via Transcriptome Analysis


 ,
,  , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Conditions
2.2. Cooling Test
2.3. Total RNA Extraction
2.4. Transcriptome High-Throughput Next-Generation Sequencing
2.5. Transcript Database Gene Differential Expression
2.6. Reverse Transcription Polymerase Chain Reaction
2.7. Real-Time Quantitative Polymerase Chain Reaction
2.8. Genomic DNA Extraction
2.9. Microsatellite Marker DNA Genotyping
2.10. Single Nucleotide Polymorphism Markers Genotyping
2.11. Statistical Analysis
3. Results
3.1. Phenotypic Differences in Cold-Tolerance as Assessed by the Loss of Balance Behavior in Response to Cooling Stress
3.2. Transcriptome Sequencing Analysis Overview
3.2.1. RNAseq Retrieval, Pre-Processing, Assembly, and Annotation of the Unigenes
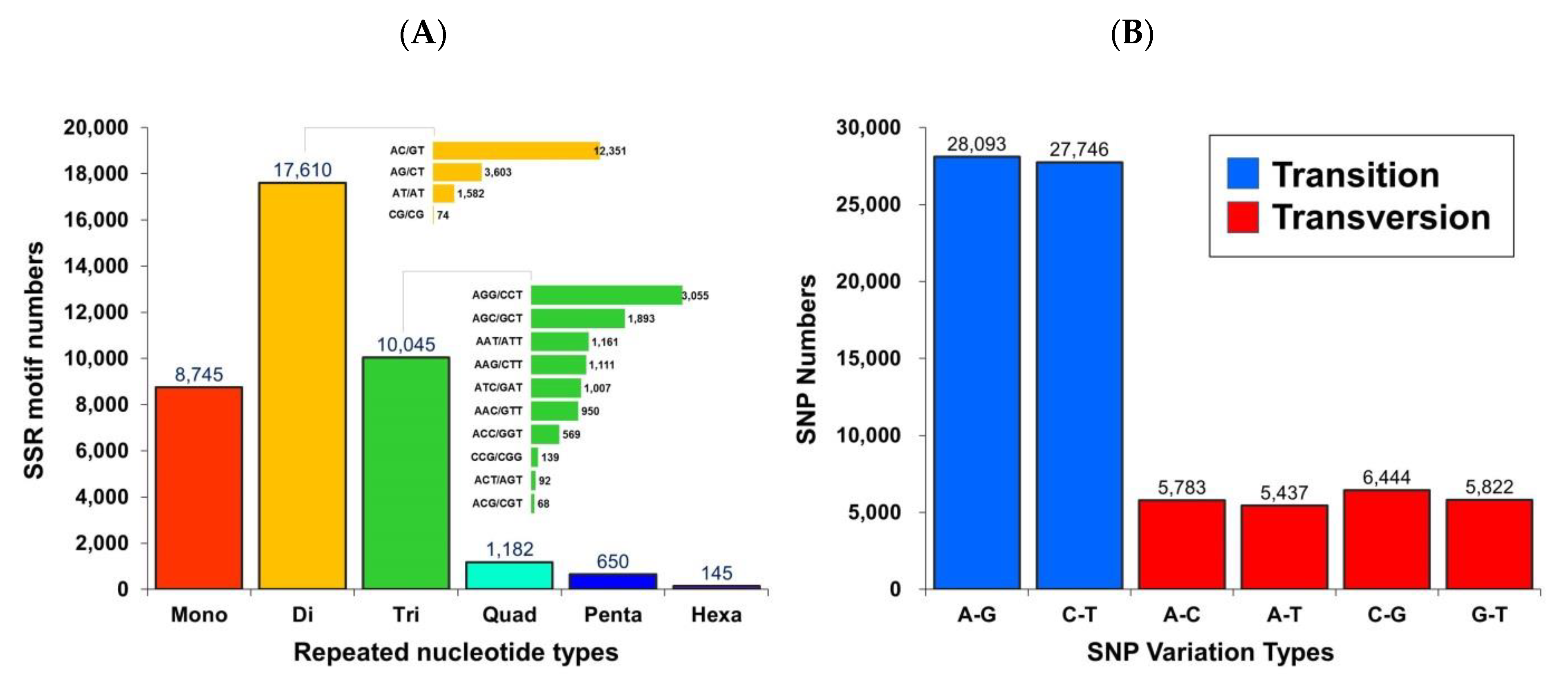
3.2.2. Detection of Microsatellites and Single Nucleotide Polymorphism Markers
3.3. Transcriptome Responses to Temperature Decreases
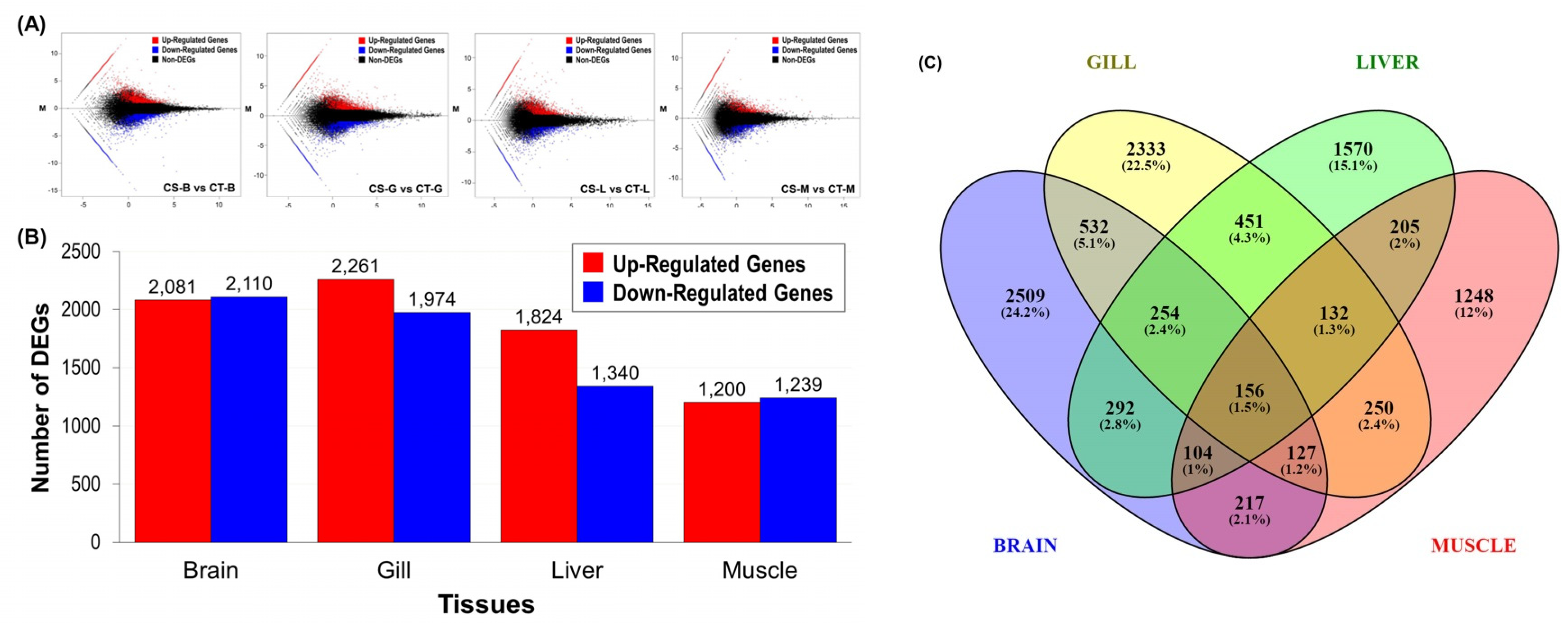
3.3.1. Differential Gene Expression between Cold-Tolerant and Sensitive Fish
3.3.2. Differential Expression of Functional Genes Containing SSRs and SNPs
3.3.3. Validation of the Transcriptome Sequencing Results Using Real-Time qPCR
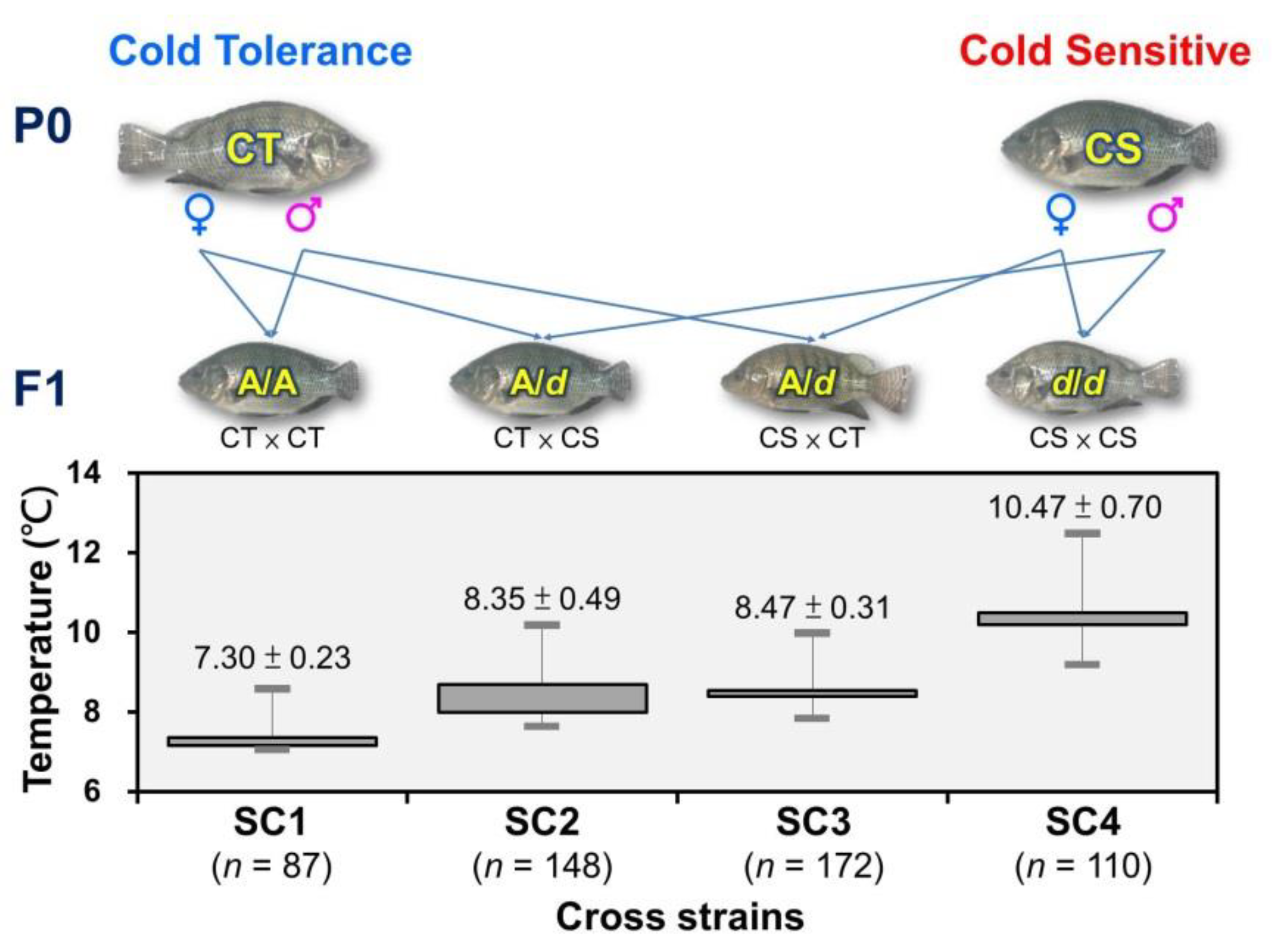
3.4. Correlation between the Genotypes of the Polymorphic RNAseq Markers and Cold-Tolerance with Significant Genetic Variation in Taiwan Tilapia
3.4.1. Identification of Candidate SSR Markers Involved in Cold-Tolerance
3.4.2. Genotype of the Gene-Based SNP Marker That Was Significantly Correlated with Cold-Tolerance
3.5. Verification of the SNP Marker Assisted Selection for Cold-Tolerant Strains in Taiwan Tilapia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agha, S.; Mekkawy, W.; Ibanez-Escriche, N.; Lind, C.E.; Kumar, J.; Mandal, A.; Benzie, J.A.H.; Doeschl-Wilson, A. Breeding for robustness: Investigating the genotype-by-environment interaction and micro-environmental sensitivity of Genetically Improved Farmed Tilapia (Oreochromis niloticus). Anim. Genet. 2018, 49, 421–427. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Cattiaux, J.; Yiou, P.; Vautard, R. Dynamics of future seasonal temperature trends and extremes in Europe: A multi-model analysis from CMIP3. Clim. Dyn. 2012, 38, 1949–1964. [Google Scholar] [CrossRef]
- Plagányi, É. Climate change impacts on fisheries. Science 2019, 363, 930–931. [Google Scholar] [CrossRef] [PubMed]
- Trewavas, E. Genus Oreochromis. In Tilapiine Fishes of the Genera Sarotherodon, Oreochromis, and Danakilia; Natural History British Museum Library: London, UK, 1983; pp. 139–193. [Google Scholar]
- Wohlfarth, G.W.; Hulata, G. Applied Genetics of Tilapias, 2nd ed.; International Center for Living Aquatic Resources Management Studies and Reviews: Manila, Philippines, 1983; pp. 1–26. [Google Scholar]
- Cnaani, A.; Gall, G.; Hulata, G. Cold tolerance of tilapia species and hybrids. Aquac. Int. 2000, 8, 289–298. [Google Scholar] [CrossRef]
- Yu, C.H.; Huang, C.J.; Sung, C.H.; Huang, J.L.; Ho-Hsu, Y.Y. Taiwan tilapia production history, traceability in seafood supply, and transfer pricing in the global market. Adv. Financ. Plan. Forecast. 2015, 6, 31–64. [Google Scholar]
- Nitzan, T.; Kokou, F.; Doron-Faigenboim, A.; Slosman, T.; Biran, J.; Mizrahi, I.; Zak, T.; Benet, A.; Cnaani, A. Transcriptome analysis reveals common and differential response to low temperature exposure between tolerant and sensitive blue tilapia (Oreochromis aureus). Front. Genet. 2019, 10, 100. [Google Scholar] [CrossRef]
- Gewaily, M.S.; Abdo, S.E.; Moustafa, E.M.; AbdEl-Kader, M.F.; Abd El-Razek, I.M.; El-Sharnouby, M.; Alkafafy, M.; Raza, S.H.A.; El Basuini, M.F.; Van Doan, H.; et al. Dietary synbiotics can help relieve the impacts of deltamethrin toxicity of Nile tilapia reared at low temperatures. Animals 2021, 11, 1790. [Google Scholar] [CrossRef] [PubMed]
- Schulte, P.M.; Healy, T.M.; Fangue, N.A. Thermal performance curves, phenotypic plasticity, and the time scales of temperature exposure. Integr. Comp. Biol. 2011, 51, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Sinyakov, M.S.; Haimovich, A.; Avtalion, R.R. Acute stress promotes post-injury brain regeneration in fish. Brain Res. 2017, 1676, 28–37. [Google Scholar] [CrossRef]
- Cowan, M.; Azpeleta, C.; López-Olmeda, J.F. Rhythms in the endocrine system of fish: A review. J. Comp. Physiol. B 2017, 187, 1057–1089. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.D.; Moriyama, S.; Björnsson, B.T. Low temperature limits photoperiod control of smolting in Atlantic salmon through endocrine mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, 1352–1361. [Google Scholar] [CrossRef]
- Miao, B.B.; Niu, S.F.; Wu, R.X.; Liang, Z.B.; Tang, B.G.; Zhai, Y.; Xu, X.Q. Gene expression profile and co-expression network of pearl gentian grouper under cold stress by integrating Illumina and PacBio sequences. Animals 2021, 11, 1745. [Google Scholar] [CrossRef] [PubMed]
- Sonna, L.A.; Fujita, J.; Gaffin, S.L.; Lilly, C.M. Invited review: Effects of heat and cold stress on mammalian gene expression. J. Appl. Physiol. 2002, 92, 1725–1742. [Google Scholar] [CrossRef] [PubMed]
- Curzon, A.Y.; Shirak, A.; Zak, T.; Dor, L.; Benet-Perlberg, A.; Naor, A.; Low-Tanne, S.I.; Sharkawi, H.; Ron, M.; Seroussi, E. All-male production by marker-assisted selection for sex determining loci of admixed Oreochromis niloticus and Oreochromis aureus stocks. Anim. Genet. 2021, 52, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Cnaani, A.; Hallerman, E.M.; Ron, M.; Weller, J.I.; Indelman, M.; Kashi, Y.; Gall, G.A.; Hulata, G. Detection of a chromosomal region with two quantitative trait loci, affecting cold tolerance and fish size, in an F2 tilapia hybrid. Aquaculture 2003, 223, 117–128. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, Z.; Lu, M.; Gao, F.; Ke, X.; Huang, Z. Screening and identification of microsatellite markers associated with cold tolerance in Nile tilapia Oreochromis niloticus. Genet. Mol. Res. 2015, 14, 10308–10314. [Google Scholar] [CrossRef]
- Ju, Z.; Dunham, R.; Liu, Z. Differential gene expression in the brain of channel catfish (Ictalurus punctatus) in response to cold acclimation. Mol. Genet. Genom. 2002, 268, 87–95. [Google Scholar] [CrossRef]
- Long, Y.; Song, G.; Yan, J.; He, X.; Li, Q.; Cui, Z. Transcriptomic characterization of cold acclimation in larval zebrafish. BMC Genom. 2013, 14, 612. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Chang, Y.; He, X.; Tang, R. Transcriptome analysis to identify cold-responsive genes in amur carp (Cyprinus carpio haematopterus). PLoS ONE 2015, 10, e0130526. [Google Scholar] [CrossRef] [PubMed]
- Hung, I.C.; Hsiao, Y.C.; Sun, H.S.; Chen, T.M.; Lee, S.J. MicroRNAs regulate gene plasticity during cold shock in zebrafish larvae. BMC Genom. 2016, 17, 922. [Google Scholar] [CrossRef] [PubMed]
- Qiang, J.; Cui, Y.T.; Tao, F.Y.; Bao, W.J.; He, J.; Li, X.H.; Xu, P.; Sun, L.Y. Physiological response and microRNA expression profiles in head kidney of genetically improved farmed tilapia (GIFT, Oreochromis niloticus) exposed to acute cold stress. Sci. Rep. 2018, 8, 172. [Google Scholar] [CrossRef]
- Chen, R.H.; Chang, Y.C.; Chang, K.C.; Liu, F.G. Comparison of growth performance of the hybridization and inbred lines in tilapia-development of fast-growing strain. J. Taiwan Fish. Res. 2008, 16, 41–47. [Google Scholar]
- Huang, C.W.; Li, Y.H.; Hu, S.Y.; Chi, J.R.; Liao, C.H.; Lin, G.H.; Lin, C.C.; Gong, H.Y.; Chen, R.H.; Chang, S.J.; et al. Differential expression patterns of growth-related microRNAs in the skeletal muscle of Nile tilapia (Oreochromis niloticus). J. Anim. Sci. 2012, 90, 4266–4279. [Google Scholar] [CrossRef]
- Kõressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating masking of template sequence with primer design software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.; Boyle, T.J.; Ye, Z.; Xiyan, J.M. PopGene32, Microsoft Window-Based Freeware for Population Genetic Analysis, version 1.32; Molecular Biology and Biotechnology Centre, University of Alberta: Edmonton, AB, Canada, 2000. [Google Scholar]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Pan, G.; Yang, J. Analysis of microsatellite DNA markers reveals no genetic differentiation between wild and hatchery populations of Pacific Threadfin in Hawaii. Int. J. Biol. Sci. 2010, 6, 827. [Google Scholar] [CrossRef]
- Khaw, H.L.; Ponzoni, R.W.; Danting, M.J.C. Estimation of genetic change in the GIFT strain of Nile tilapia (Oreochromis niloticus) by comparing contemporary progeny produced by males born in 1991 or in 2003. Aquaculture 2008, 275, 64–69. [Google Scholar] [CrossRef]
- He, J.; Qiang, J.; Yang, H.; Xu, P.; Zhu, Z.; Yang, R. Changes in the fatty acid composition and regulation of antioxidant enzymes and physiology of juvenile genetically improved farmed tilapia Oreochromis niloticus (L.), subjected to short-term low temperature stress. J. Therm. Biol. 2015, 53, 90–99. [Google Scholar] [CrossRef]
- Sifa, L.; Chenhong, L.; Dey, M.; Gagalac, F.; Dunham, R. Cold tolerance of three strains of Nile tilapia, Oreochromis niloticus, in China. Aquaculture 2002, 213, 123–129. [Google Scholar] [CrossRef]
- Agnèse, J.F.; Adépo-Gourène, B.; Abban, E.K.; Fermon, Y. Genetic differentiation among natural populations of the Nile tilapia Oreochromis niloticus (Teleostei, Cichlidae). Heredity 1997, 79, 88–96. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Carroll, S.B. Evo-devo and an expanding evolutionary synthesis: A genetic theory of morphological evolution. Cell 2008, 134, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, H.E.; Coyne, J.A. The locus of evolution: Evo devo and the genetics of adaptation. Evolution 2007, 61, 995–1016. [Google Scholar] [CrossRef] [PubMed]
- Romero, I.G.; Ruvinsky, I.; Gilad, Y. Comparative studies of gene expression and the evolution of gene regulation. Nat. Rev. Genet. 2012, 13, 505–516. [Google Scholar] [CrossRef]
- Khater, A.A.; Smitherman, R.O. Cold tolerance and growth of three strains of Oreochromis niloticus. In Proceedings of the 2nd International Symposium on Tilapia in Aquaculture, ICLARM Conference Proceedings, Bangkok, Thailand, 16–20 March 1987; Pullin, R.S.V., Bhukaswan, T., Tonguthai, K., Maclean, J.L., Eds.; Department of Fisheries: Metro Manila, Philippines, 1988; pp. 215–218. [Google Scholar]
- Cossins, A.R.; Crawford, D.L. Fish as models for environmental genomics. Nat. Rev. Genet. 2005, 6, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Qiang, J.; He, J.; Gabriel, N.N.; Xu, P. Changes in the physiological parameters, fatty acid metabolism, and SCD activity and expression in juvenile GIFT tilapia (Oreochromis niloticus) reared at three different temperatures. Fish. Physiol. Biochem. 2015, 41, 937–950. [Google Scholar] [CrossRef] [PubMed]
- King, M.; Sardella, B. The effects of acclimation temperature, salinity, and behavior on the thermal tolerance of Mozambique tilapia (Oreochromis mossambicus). J. Exp. Zool. A Ecol. Integr. Physiol. 2017, 327, 417–422. [Google Scholar] [CrossRef]
- Kokou, F.; Sasson, G.; Nitzan, T.; Doron-Faigenboim, A.; Harpaz, S.; Cnaani, A.; Mizrahi, I. Host genetic selection for cold tolerance shapes microbiome composition and modulates its response to temperature. Elife 2018, 7, e36398. [Google Scholar] [CrossRef]
- Long, Y.; Liu, R.; Song, G.; Li, Q.; Cui, Z. Establishment and characterization of a cold-sensitive neural cell line from the brain of tilapia (Oreochromis niloticus). J. Fish. Biol. 2021, 98, 842–854. [Google Scholar] [CrossRef]
- Hu, P.; Liu, M.; Liu, Y.; Wang, J.; Zhang, D.; Niu, H.; Jiang, S.; Wang, J.; Zhang, D.; Han, B. Transcriptome comparison reveals a genetic network regulating the lower temperature limit in fish. Sci. Rep. 2016, 6, 28952. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, R.E.; El-Houseiny, W.; Behairy, A.; Abo-Elmaaty, A.; Al-Sagheer, A.A. The palliative role of Eruca sativa leaves dietary supplementation against oxidative stress, immunosuppression, and growth retardation in temperature-stressed Oreochromis niloticus. J. Therm. Biol. 2019, 84, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Lima de Almeida, C.A.; Lima de Almeida, C.K.; de Fátima Ferreira Martins, E.; Gomes, Â.M.; da Anunciação Pimentel, L.; Pereira, R.T.; Fortes-Silva, R. Effect of the dietary linoleic/alpha-linolenic ratio (n6/n3) on histopathological alterations caused by suboptimal temperature in tilapia (Oreochromis niloticus). J. Therm. Biol. 2019, 85, 102386. [Google Scholar] [CrossRef]
- Zhou, T.; Gui, L.; Liu, M.; Li, W.; Hu, P.; Duarte, D.F.C.; Niu, H.; Chen, L. Transcriptomic responses to low temperature stress in the Nile tilapia, Oreochromis niloticus. Fish. Shellfish Immunol. 2019, 84, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; You, F.; Wang, Q.; Weng, S.; Liu, H.; Wang, L.; Zhang, P.J.; Tan, X. Transcriptional responses of olive flounder (Paralichthys olivaceus) to low temperature. PLoS ONE 2014, 9, e108582. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.C.; Kang, C.K.; Tang, C.H.; Lee, T.H. Transcriptomic analysis of metabolic pathways in milkfish that respond to salinity and temperature changes. PLoS ONE 2015, 10, e0134959. [Google Scholar]
- Liu, S.; Vallejo, R.L.; Palti, Y.; Gao, G.; Marancik, D.P.; Hernandez, A.G.; Wiens, G.D. Identification of single nucleotide polymorphism markers associated with bacterial cold water disease resistance and spleen size in rainbow trout. Front. Genet. 2015, 6, 298. [Google Scholar] [CrossRef]
- Gracey, A.Y.; Fraser, E.J.; Li, W.; Fang, Y.; Taylor, R.R.; Rogers, J.; Brass, A.; Cossins, A.R. Coping with cold: An integrative, multitissue analysis of the transcriptome of a poikilothermic vertebrate. Proc. Natl. Acad. Sci. USA 2004, 101, 16970–16975. [Google Scholar] [CrossRef] [PubMed]
- Assefa, A.T.; Vandesompele, J.; Thas, O. On the utility of RNA sample pooling to optimize cost and statistical power in RNA sequencing experiments. BMC Genom. 2020, 21, 312. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, C.; Mu, J.; Zhang, H.; Yao, W.; Ding, X.; Ding, J.; Chang, Y. All-in-one sequencing: An improved library preparation method for cost-effective and high-throughput next-generation sequencing. Plant. Methods 2020, 16, 74. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Furuse, M. Tight Junction Structure and Function Revisited. Trends Cell Biol. 2020, 30, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A.; Orian, A.; Schwartz, A.L. Ubiquitin-mediated proteolysis: Biological regulation via destruction. Bioessays 2000, 22, 442–451. [Google Scholar] [CrossRef]
- Seger, R.; Krebs, E.G. The MAPK signaling cascade. FASEB J. 1995, 9, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Lind, C.E.; Kilian, A.; Benzie, J.A.H. Development of Diversity Arrays Technology markers as a tool for rapid genomic assessment in Nile tilapia, Oreochromis niloticus. Anim. Genet. 2017, 48, 362–364. [Google Scholar] [CrossRef]
- Landegren, U.; Nilsson, M.; Kwok, P.Y. Reading bits of genetic information: Methods for single-nucleotide polymorphism analysis. Genome Res. 1998, 8, 769–776. [Google Scholar] [CrossRef]
- Houston, R.D.; Taggart, J.B.; Cézard, T.; Bekaert, M.; Lowe, N.R.; Downing, A.; Talbot, R.; Bishop, S.C.; Archibald, A.L.; Bron, J.E. Development and validation of a high density SNP genotyping array for Atlantic salmon (Salmo salar). BMC Genom. 2014, 15, 90. [Google Scholar] [CrossRef] [PubMed]
- Van Bers, N.; Crooijmans, R.; Groenen, M.; Dibbits, B.; Komen, J. SNP marker detection and genotyping in tilapia. Mol. Ecol. Resour. 2012, 12, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Xu, J.; Ji, P.; Zhao, Z.; Zhang, Y.; Feng, J.; Wang, J.; Li, J.; Zhang, X.; Zhao, L.; Liu, G. Genome-wide SNP discovery from transcriptome of four common carp strains. PLoS ONE 2012, 7, e48140. [Google Scholar] [CrossRef]
- Xia, J.H.; Wan, Z.Y.; Ng, Z.L.; Wang, L.; Fu, G.H.; Lin, G.; Liu, F.; Yue, G.H. Genome-wide discovery and in silico mapping of gene-associated SNPs in Nile tilapia. Aquaculture 2014, 432, 67–73. [Google Scholar] [CrossRef]
- Peñaloza, C.; Robledo, D.; Barría, A.; Trịnh, T.Q.; Mahmuddin, M.; Wiener, P.; Benzie, J.A.H.; Houston, R.D. Development and validation of an open access SNP array for Nile tilapia (Oreochromis niloticus). G3 2020, 10, 2777–2785. [Google Scholar] [CrossRef] [PubMed]
- Yáñez, J.M.; Joshi, R.; Yoshida, G.M. Genomics to accelerate genetic improvement in tilapia. Anim. Genet. 2020, 51, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Yáñez, J.M.; Yoshida, G.; Barria, A.; Palma-Véjares, R.; Travisany, D.; Díaz, D.; Cáceres, G.; Cádiz, M.I.; López, M.E.; Lhorente, J.P.; et al. High-throughput single nucleotide polymorphism (SNP) discovery and validation through whole-genome resequencing in Nile tilapia (Oreochromis niloticus). Mar. Biotechnol. 2020, 22, 109–117. [Google Scholar] [CrossRef]
- Majeed, S.R.; Vasudevan, L.; Chen, C.Y.; Luo, Y.; Torres, J.A.; Evans, T.M.; Sharkey, A.; Foraker, A.B.; Wong, N.M.; Esk, C. Clathrin light chains are required for the gyrating-clathrin recycling pathway and thereby promote cell migration. Nat. Commun. 2014, 5, 3891. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V.; Korbelová, J.; Poupardin, R.; Moos, M.; Šimek, P. Arginine and proline applied as food additives stimulate high freeze tolerance in larvae of Drosophila melanogaster. J. Exp. Biol. 2016, 219, 2358–2367. [Google Scholar] [CrossRef]
- Fan, L.; Wang, L.; Wang, Z. Proteomic characterization of the hepatopancreas in the Pacific white shrimp Litopenaeus vannamei under cold stress: Revealing the organism homeostasis mechanism. Fish. Shellfish Immunol. 2019, 92, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Ghany, H.M.; El-Sayed, A.M.; Ezzat, A.A.; Essa, M.A.; Helal, A.M. Dietary lipid sources affect cold tolerance of Nile tilapia (Oreochromis niloticus). J. Therm. Biol. 2019, 79, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Y.; Liu, N.; Chen, J.; Guo, L.; Dai, Z.; Wang, C.; Wu, Z.; Wu, G. Dietary L-arginine supplementation reduces lipid accretion by regulating fatty acid metabolism in Nile tilapia (Oreochromis niloticus). J. Anim. Sci. Biotechnol. 2020, 11, 82. [Google Scholar] [CrossRef]
- Tian, J.; Wen, H.; Lu, X.; Liu, W.; Wu, F.; Yang, C.G.; Jiang, M.; Yu, L.J. Dietary phosphatidylcholine impacts on growth performance and lipid metabolism in adult Genetically Improved Farmed Tilapia (GIFT) strain of Nile tilapia Oreochromis niloticus. Br. J. Nutr. 2018, 119, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Chi, J.R.; Huang, C.W.; Wu, J.L.; Hu, S.Y. Prolactin I microsatellite as genetic markers for characterization of five Oreochromis tilapia species and two Oreochromis Niloticus strains. J. Aquac. Res. Dev. 2014, 5, 251. [Google Scholar] [CrossRef]
- Lu, J.K.; Chou, H.Y.; Gong, H.Y.; Chiou, P.P.; Huang, C.W.; Chen, L.L.; Wu, J.L. The Marine aquaculture biotechnology status and its sustainability in Taiwan. Curr. Biotechnol. 2015, 4, 311–318. [Google Scholar] [CrossRef][Green Version]
- Herkenhoff, M.E.; Ribeiro, A.O.; Costa, J.M.; Oliveira, A.C.; Dias, M.A.D.; Reis Neto, R.V.; Hilsdorf, A.W.S.; Pinhal, D. Expression profiles of growth-related genes in two Nile tilapia strains and their crossbred provide insights into introgressive breeding effects. Anim. Genet. 2020, 51, 611–616. [Google Scholar] [CrossRef]
- Wang, J.H.; Lee, S.K.; Lai, Y.C.; Lin, C.C.; Wang, T.Y.; Lin, Y.R.; Hsu, T.H.; Huang, C.W.; Chiang, C.P. Anomalous Behaviors Detection for Underwater Fish Using AI Techniques. IEEE Access 2020, 8, 1–11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Clean Reads 2 | Clean Bases 3 | Q20 (%) 4 | Q30 (%) 5 | GC (%) 6 | RL (bp) 7 |
|---|---|---|---|---|---|---|
| CT-B | 29,473,122 | 4,420,968,300 | 98.88 | 96.28 | 49.36 | 150 |
| CT-G | 29,944,552 | 4,491,682,800 | 98.69 | 95.63 | 51.22 | 150 |
| CT-L | 29,536,538 | 4,430,480,700 | 98.76 | 95.83 | 50.54 | 150 |
| CT-M | 29,268,400 | 4,390,260,000 | 98.9 | 96.27 | 52.25 | 150 |
| CS-B | 29,547,582 | 4,432,137,300 | 98.72 | 95.85 | 49.57 | 150 |
| CS-G | 28,697,764 | 4,304,664,600 | 98.8 | 96.04 | 49.92 | 150 |
| CS-L | 29,117,880 | 4,367,682,000 | 99.05 | 96.72 | 50.07 | 150 |
| CS-M | 29,179,716 | 4,376,957,400 | 98.81 | 96.02 | 52.11 | 150 |
| Transcriptome Dataset | Unigene Number | Percentage (%) |
|---|---|---|
| NR 1 | 74,656 | 58.26 |
| NT 2 | 97,575 | 76.14 |
| Swiss-Prot 3 | 61,098 | 47.68 |
| COG 4 | 26,342 | 20.56 |
| CO 5 | 7154 | 5.58 |
| KEGG 6 | 59,699 | 46.59 |
| Overall (total annotation) 7 | 100,108 | 78.12 |
| Total | 128,147 | 100 |
| No. | Unigene ID | SSR | Length | Position | LG 1 | Location | Gene Annotation 2 |
|---|---|---|---|---|---|---|---|
| 1 | CL138_1 | (A)n | 2492 | 1902 | LG7 | 3′-UTR | CTD small phosphatase-like protein 2-A |
| 2 | CL1487_25 | (GT)n | 8664 | 1862 | LG16 | 5′-UTR | Nuclear pore complex protein Nup98-Nup96 isoform X6 |
| 3 | CL1876_16 | (TTC)n | 7751 | 7251 | LG4 | 3′-UTR | Myosin-10 isoform X3 |
| 4 | CL5902_1 | (G)n | 2572 | 569 | LG12 | 3′-UTR | Ubiquitin-conjugating enzyme E2 G1 |
| 5 | CL9541_2 | (CAG)n | 5825 | 2164 | LG17 | Exon | AT-rich interactive domain-containing protein 2 |
| 6 | CL10781_10 | (A)n | 9102 | 8403 | LG8 | 3′-UTR | Fatty acid synthase isoform X1 |
| 7 | CL279_7 | (GCA)n | 4772 | 2266 | LG23 | Exon | R3H domain-containing protein 1 |
| 8 | CL2262_1 | (T)n | 3105 | 3083 | LG2 | 3′-UTR | TNFAIP3-interacting protein 1 isoform X1 |
| 9 | Unigene7071 | (GT)n | 2729 | 1722 | LG14 | Intron | Protein IWS1 homolog isoform X3 |
| 10 | Unigene196 | (CA)n | 5125 | 5073 | LG14 | 3′-UTR | Serine/threonine-protein kinase SIK3 isoform X3 |
| 11 | CL2061_8 | (TC)n | 1393 | 102 | LG7 | 5′-UTR | Glucose-6-phosphate isomerase-like |
| 12 | CL9318_1 | (ATT)n | 706 | 530 | LG12 | 3′-UTR | Bifunctional methylenetetrahydrofolate dehydrogenase/cyclohydrolase, mitochondrial-like |
| 13 | CL241_10 | (GAG)n | 5371 | 3079 | LG8 | Exon | Rho GTPase-activating protein 17 isoform X1 |
| SNP | Unigene ID | Length | LG 1 | Allele | Location | Change 2 | Gene Annotation 3 |
|---|---|---|---|---|---|---|---|
| 1 | CL4970 | 3236 | 2 | T/C | UTR | - | pre-mRNA-splicing factor RBM22 |
| 2 | CL6296_2 | 3447 | 23 | d/A | UTR | - | lipoamide acyltransferase component of branched-chain alpha-keto acid dehydrogenase complex, mitochondrial |
| 3 | CL2980_2 | 1540 | 3 | G/C | Exon | P/P | NADH dehydrogenase |
| 4 | CL6447_4 | 3573 | ND | C/T | Intron | - | WD repeat domain phosphoinositide-interacting protein 2 isoform X1 |
| 5 | CL4350_5 | 3286 | 23 | G/A | UTR | - | protein Jade-3 |
| 6 | CL4943_1 | 2166 | 2 | A/G | UTR | - | NEDD4 family-interacting protein 1 isoform X2 |
| 7 | CL179_6 | 5803 | 5 | G/A | Exon | V/I | period circadian protein homolog 3-like isoform X4 |
| 8 | CL4788_1 | 1945 | 4 | d/G | UTR | - | hsc70-interacting protein isoform X2 |
| 9 | CL4788_1 | 1945 | 4 | d/A | UTR | - | |
| 10 | CL5212_1 | 1774 | 2 | d/A | UTR | - | clathrin light chain B-like isoform X4 |
| 11 | CL11264_1 | 2075 | 2 | G/A | UTR | - | cyclin-G1-like |
| 12 | CL11264_1 | 2075 | 2 | C/T | UTR | - | |
| 13 | CL11264_1 | 2075 | 2 | C/A | UTR | - | |
| 14 | CL11264_1 | 2075 | 2 | T/C | UTR | - | |
| 15 | CL11264_1 | 2075 | 2 | G/A | Exon | I/V | |
| 16 | CL3637_1 | 10,003 | 23 | T/A | UTR | - | protein PRRC2C |
| 17 | CL715_12 | 5227 | 13 | d/A | Exon | S/K | adipocyte plasma membrane-associated protein isoform X2 |
| 18 | CL715_12 | 5227 | 13 | d/C | UTR | - | |
| 19 | CL715_12 | 5227 | 13 | d/C | UTR | - | |
| 20 | CL715_12 | 5227 | 13 | d/G | UTR | - | |
| 21 | CL8349_2 | 3478 | 7 | T/C | UTR | - | ATP-dependent Clp protease ATP-binding subunit clpX-like, mitochondrial isoform X1 |
| 22 | CL4663_1 | 3831 | ND | A/T | UTR | - | bone morphogenetic protein receptor type-1A |
| 23 | CL11264_2 | 2074 | 2 | A/G | UTR | - | cyclin-G1-like |
| 24 | CL11264_2 | 2074 | 2 | C/T | UTR | - | |
| 25 | CL11264_2 | 2074 | 2 | G/T | UTR | - | |
| 26 | CL4356_2 | 1809 | ND | d/G | UTR | - | bone morphogenetic protein receptor type-1A |
| 27 | CL1248_2 | 3831 | 2 | C/T | Intron | - | dynactin subunit 4 isoform X2 |
| 28 | CL2394_12 | 8460 | 13 | G/A | Exon | R/H | spectrin beta chain, non-erythrocytic 1 |
| 29 | CL2394_12 | 8460 | 13 | C/T | Exon | V/I | |
| 30 | CL4788_2 | 2157 | 4 | A/G | Exon | A/A | hsc70-interacting protein isoform X2 |
| 31 | CL144_1 | 1772 | 13 | d/G | UTR | - | E3 ubiquitin-protein ligase UBR2 isoform X3 |
| 32 | CL4659_4 | 3929 | 13 | d/T | Exon | F/F | echinoderm microtubule-associated protein-like 4 isoform X3 |
| 33 | CL179_13 | 1906 | 5 | d/T | UTR | - | period circadian protein homolog 3-like isoform X4 |
| 34 | CL569_1 | 3529 | 13 | d/A | Exon | P/P | cullin-9 isoform X1 |
| 35 | CL2216_1 | 1929 | 8 | T/C | Exon | S/T | dynamin-binding protein isoform X3 |
| 36 | CL4659_13 | 4077 | 13 | d/T | Exon | D/D | echinoderm microtubule-associated protein-like 4 isoform X3 |
| 37 | CL4659_13 | 4077 | 13 | d/A | Exon | T/N |
| Locus 1 | Ho | He | PIC | FIS | p Value 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RF | YH | NT | RF | YH | NT | RF | YH | NT | RF | YH | NT | ||
| UNH916 | 0.722 | 0.480 | 0.532 | 0.775 | 0.485 | 0.395 | 0.769 | 0.483 | 0.390 | 0.061 | 0.004 | −0.362 | 0.345 |
| UNH999 | 0.778 | 0.686 | 0.447 | 0.767 | 0.671 | 0.395 | 0.761 | 0.668 | 0.390 | −0.022 | −0.028 | −0.144 | 0.103 |
| CL1487_25 | 0.472 | 0.726 | 0.000 | 0.624 | 0.676 | 0.000 | 0.620 | 0.672 | 0.000 | 0.239 | −0.079 | — | 0.537 |
| CL1876_16 | 0.181 | 0.000 | 0.000 | 0.289 | 0.000 | 0.000 | 0.287 | 0.000 | 0.000 | 0.371 | — | — | 0.004 ** |
| CL5902_1 | 0.903 | 0.441 | 0.000 | 0.753 | 0.658 | 0.000 | 0.747 | 0.655 | 0.000 | −0.208 | 0.326 | — | 0.456 |
| CL9541_2 | 0.472 | 0.598 | 0.447 | 0.523 | 0.472 | 0.351 | 0.520 | 0.470 | 0.347 | 0.091 | −0.273 | −0.288 | 0.217 |
| CL10781_10 | 0.278 | 0.559 | 0.489 | 0.412 | 0.592 | 0.599 | 0.410 | 0.589 | 0.593 | 0.322 | 0.051 | 0.175 | 0.178 |
| CL279_7 | 0.000 | 0.000 | 0.000 | 0.130 | 0.000 | 0.000 | 0.129 | 0.000 | 0.000 | 1.000 | — | — | 0.017 * |
| CL2262_1 | 0.722 | 0.784 | 0.894 | 0.798 | 0.728 | 0.678 | 0.793 | 0.725 | 0.671 | 0.089 | −0.082 | −0.332 | 0.257 |
| Unigene7071 | 0.694 | 0.196 | 0.575 | 0.749 | 0.201 | 0.431 | 0.744 | 0.200 | 0.427 | 0.066 | 0.021 | −0.346 | 0.011 * |
| Unigene196 | 0.750 | 0.686 | 0.553 | 0.776 | 0.710 | 0.454 | 0.771 | 0.706 | 0.449 | 0.027 | 0.028 | −0.232 | 0.098 |
| CL9318_1 | 0.667 | 0.284 | 0.000 | 0.759 | 0.283 | 0.000 | 0.754 | 0.282 | 0.000 | 0.116 | −0.010 | — | 0.010 * |
| CL241_10 | 0.319 | 0.382 | 0.575 | 0.383 | 0.367 | 0.505 | 0.381 | 0.365 | 0.499 | 0.161 | −0.048 | −0.151 | 0.255 |
| Mean | 0.535 | 0.448 | 0.347 | 0.595 | 0.449 | 0.293 | 0.591 | 0.447 | 0.290 | 0.178 | −0.008 | −0.210 | |
| SD | 0.272 | 0.264 | 0.306 | 0.225 | 0.260 | 0.256 | 0.223 | 0.259 | 0.253 | 0.277 | 0.135 | 0.165 | |
| Unigene | SNP | High Mass Allele Calls | Heterozygous Calls | Low Mass Allele Calls | Homozygous freq | Heterozygous freq | HW ChiSquare | HW p-Value | Trait p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RF | YH | NT | RF | YH | NT | RF | YH | NT | RF | YH | NT | RF | YH | NT | |||||
| CL4970 | C/T | 0 | 0 | 0 | 0 | 2 | 0 | 72 | 94 | 22 | 1.000 | 0.979 | 1.000 | 0.000 | 0.021 | 0.000 | 0.01 | 0.940 | 0.879 |
| CL6296_2 | A/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL2980_2 | C/G | 13 | 13 | 0 | 31 | 35 | 0 | 27 | 45 | 22 | 0.563 | 0.624 | 1.000 | 0.437 | 0.376 | 0.000 | 6.08 | 0.010 | 0.148 |
| CL6447_4 | C/T | 25 | 49 | 0 | 28 | 36 | 22 | 15 | 10 | 0 | 0.588 | 0.621 | 0.000 | 0.412 | 0.379 | 1.000 | 0.00 | 1.000 | 0.056 |
| CL4350_5 | A/G | 11 | 0 | 0 | 23 | 0 | 13 | 38 | 96 | 9 | 0.681 | 1.000 | 0.409 | 0.319 | 0.000 | 0.591 | 13.60 | 0.000 | 0.589 |
| CL4943_1 | A/G | 37 | 73 | 22 | 28 | 20 | 0 | 2 | 3 | 0 | 0.582 | 0.792 | 1.000 | 0.418 | 0.208 | 0.000 | 0.06 | 0.800 | 0.919 |
| CL179_6 | G/A | 58 | 89 | 22 | 13 | 7 | 0 | 1 | 0 | 0 | 0.819 | 0.927 | 1.000 | 0.181 | 0.073 | 0.000 | 0.23 | 0.630 | 0.591 |
| CL4788_1_1 | G/d | 16 | 10 | 0 | 29 | 37 | 22 | 27 | 48 | 0 | 0.597 | 0.611 | 0.000 | 0.403 | 0.389 | 1.000 | 0.00 | 0.980 | 0.094 |
| CL4788_1_2 | A/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL5212_1 | A/d | 11 | 0 | 0 | 30 | 13 | 0 | 31 | 83 | 22 | 0.583 | 0.865 | 1.000 | 0.417 | 0.135 | 0.000 | 7.75 | 0.010 | 0.001 ** |
| CL11264_1_1 | G/A | 38 | 95 | 22 | 28 | 1 | 0 | 6 | 0 | 0 | 0.611 | 0.990 | 1.000 | 0.389 | 0.010 | 0.000 | 8.15 | 0.000 | 0.378 |
| CL11264_1_2 | C/T | 38 | 95 | 22 | 27 | 1 | 0 | 7 | 0 | 0 | 0.625 | 0.990 | 1.000 | 0.375 | 0.010 | 0.000 | 11.92 | 0.000 | 0.602 |
| CL11264_1_3 | C/A | 37 | 95 | 22 | 27 | 1 | 0 | 7 | 0 | 0 | 0.620 | 0.990 | 1.000 | 0.380 | 0.010 | 0.000 | 11.81 | 0.000 | 0.599 |
| CL11264_1_4 | T/C | 1 | 0 | 0 | 25 | 1 | 0 | 46 | 95 | 22 | 0.653 | 0.990 | 1.000 | 0.347 | 0.010 | 0.000 | 0.00 | 0.970 | 0.650 |
| CL11264_1_5 | G/A | 7 | 0 | 0 | 26 | 1 | 0 | 38 | 95 | 22 | 0.634 | 0.990 | 1.000 | 0.366 | 0.010 | 0.000 | 12.91 | 0.000 | 0.490 |
| CL3637_1 | T/A | 38 | 96 | 22 | 25 | 0 | 0 | 9 | 0 | 0 | 0.653 | 1.000 | 1.000 | 0.347 | 0.000 | 0.000 | 22.54 | 0.000 | 0.473 |
| CL715_12_1 | A/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL715_12_2 | C/d | 28 | 7 | 0 | 35 | 43 | 22 | 9 | 46 | 0 | 0.514 | 0.552 | 0.000 | 0.486 | 0.448 | 1.000 | 242.63 | 0.000 | 0.099 |
| CL715_12_3 | C/d | 9 | 0 | 0 | 24 | 0 | 22 | 38 | 96 | 0 | 0.662 | 1.000 | 0.000 | 0.338 | 0.000 | 1.000 | 200.20 | 0.000 | 0.488 |
| CL715_12_4 | G/d | 0 | 0 | 0 | 0 | 0 | 0 | 67 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL8349_2 | C/T | 0 | 28 | 12 | 6 | 50 | 10 | 66 | 18 | 0 | 0.917 | 0.479 | 0.545 | 0.083 | 0.521 | 0.455 | 13.43 | 0.000 | 0.366 |
| CL4663_1 | T/A | 4 | 0 | 0 | 21 | 0 | 22 | 38 | 95 | 0 | 0.667 | 1.000 | 0.000 | 0.333 | 0.000 | 1.000 | 0.06 | 0.810 | 0.769 |
| CL11264_2_1 | G/A | 2 | 0 | 0 | 27 | 1 | 0 | 43 | 95 | 22 | 0.625 | 0.990 | 1.000 | 0.375 | 0.010 | 0.000 | 0.38 | 0.540 | 0.650 |
| CL11264_2_2 | C/T | 46 | 95 | 22 | 25 | 1 | 0 | 1 | 0 | 0 | 0.653 | 0.990 | 1.000 | 0.347 | 0.010 | 0.000 | 0.00 | 0.970 | 0.650 |
| CL11264_2_3 | G/T | 41 | 95 | 22 | 25 | 1 | 0 | 1 | 0 | 0 | 0.627 | 0.990 | 1.000 | 0.373 | 0.010 | 0.000 | 0.00 | 0.950 | 0.650 |
| CL4356_2 | G/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL1248_2 | T/C | 4 | 0 | 0 | 27 | 3 | 0 | 40 | 93 | 22 | 0.620 | 0.969 | 1.000 | 0.380 | 0.031 | 0.000 | 2.83 | 0.090 | 0.581 |
| CL2394_12_1 | T/C | 4 | 0 | 0 | 24 | 9 | 22 | 44 | 87 | 0 | 0.667 | 0.906 | 0.000 | 0.333 | 0.094 | 1.000 | 0.41 | 0.520 | 0.888 |
| CL2394_12_2 | A/G | 4 | 0 | 0 | 24 | 0 | 22 | 44 | 96 | 0 | 0.667 | 1.000 | 0.000 | 0.333 | 0.000 | 1.000 | 0.01 | 0.920 | 0.888 |
| CL4788_2 | G/A | 23 | 80 | 14 | 35 | 15 | 8 | 14 | 1 | 0 | 0.514 | 0.844 | 0.636 | 0.486 | 0.156 | 0.364 | 3.85 | 0.050 | 0.104 |
| CL144_1 | G/d | 0 | 0 | 0 | 0 | 0 | 0 | 70 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL4659_4 | T/d | 4 | 0 | 0 | 23 | 0 | 22 | 44 | 96 | 0 | 0.676 | 1.000 | 0.000 | 0.324 | 0.000 | 1.000 | 199.71 | 0.000 | 0.926 |
| CL179_13 | T/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL569_1 | A/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL2216_1 | T/C | 70 | 96 | 12 | 1 | 0 | 10 | 0 | 0 | 0 | 0.986 | 1.000 | 0.545 | 0.014 | 0.000 | 0.455 | 0.17 | 0.680 | 0.478 |
| CL4659_13_1 | T/d | 0 | 0 | 0 | 0 | 0 | 0 | 72 | 96 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
| CL4659_13_2 | A/d | 0 | 0 | 0 | 0 | 0 | 0 | 66 | 95 | 22 | 1.000 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, P.-Y.; Li, J.-X.; Hsu, T.-H.; Gong, H.-Y.; Lin, C.-Y.; Wang, J.-H.; Huang, C.-W. Identification of Genes Related to Cold Tolerance and Novel Genetic Markers for Molecular Breeding in Taiwan Tilapia (Oreochromis spp.) via Transcriptome Analysis. Animals 2021, 11, 3538. https://doi.org/10.3390/ani11123538
Chu P-Y, Li J-X, Hsu T-H, Gong H-Y, Lin C-Y, Wang J-H, Huang C-W. Identification of Genes Related to Cold Tolerance and Novel Genetic Markers for Molecular Breeding in Taiwan Tilapia (Oreochromis spp.) via Transcriptome Analysis. Animals. 2021; 11(12):3538. https://doi.org/10.3390/ani11123538
Chicago/Turabian StyleChu, Pei-Yun, Jia-Xian Li, Te-Hua Hsu, Hong-Yi Gong, Chung-Yen Lin, Jung-Hua Wang, and Chang-Wen Huang. 2021. "Identification of Genes Related to Cold Tolerance and Novel Genetic Markers for Molecular Breeding in Taiwan Tilapia (Oreochromis spp.) via Transcriptome Analysis" Animals 11, no. 12: 3538. https://doi.org/10.3390/ani11123538
APA StyleChu, P.-Y., Li, J.-X., Hsu, T.-H., Gong, H.-Y., Lin, C.-Y., Wang, J.-H., & Huang, C.-W. (2021). Identification of Genes Related to Cold Tolerance and Novel Genetic Markers for Molecular Breeding in Taiwan Tilapia (Oreochromis spp.) via Transcriptome Analysis. Animals, 11(12), 3538. https://doi.org/10.3390/ani11123538