
Timing and Weather Offer Alternative Mitigation Strategies for Lowering Bat Mortality at Wind Energy Facilities in Ontario


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials & Methods
2.1. Bat Mortality
2.2. Bat Activity
2.3. Predictions and Covariates in Models
2.4. Statistical Analyses
3. Results
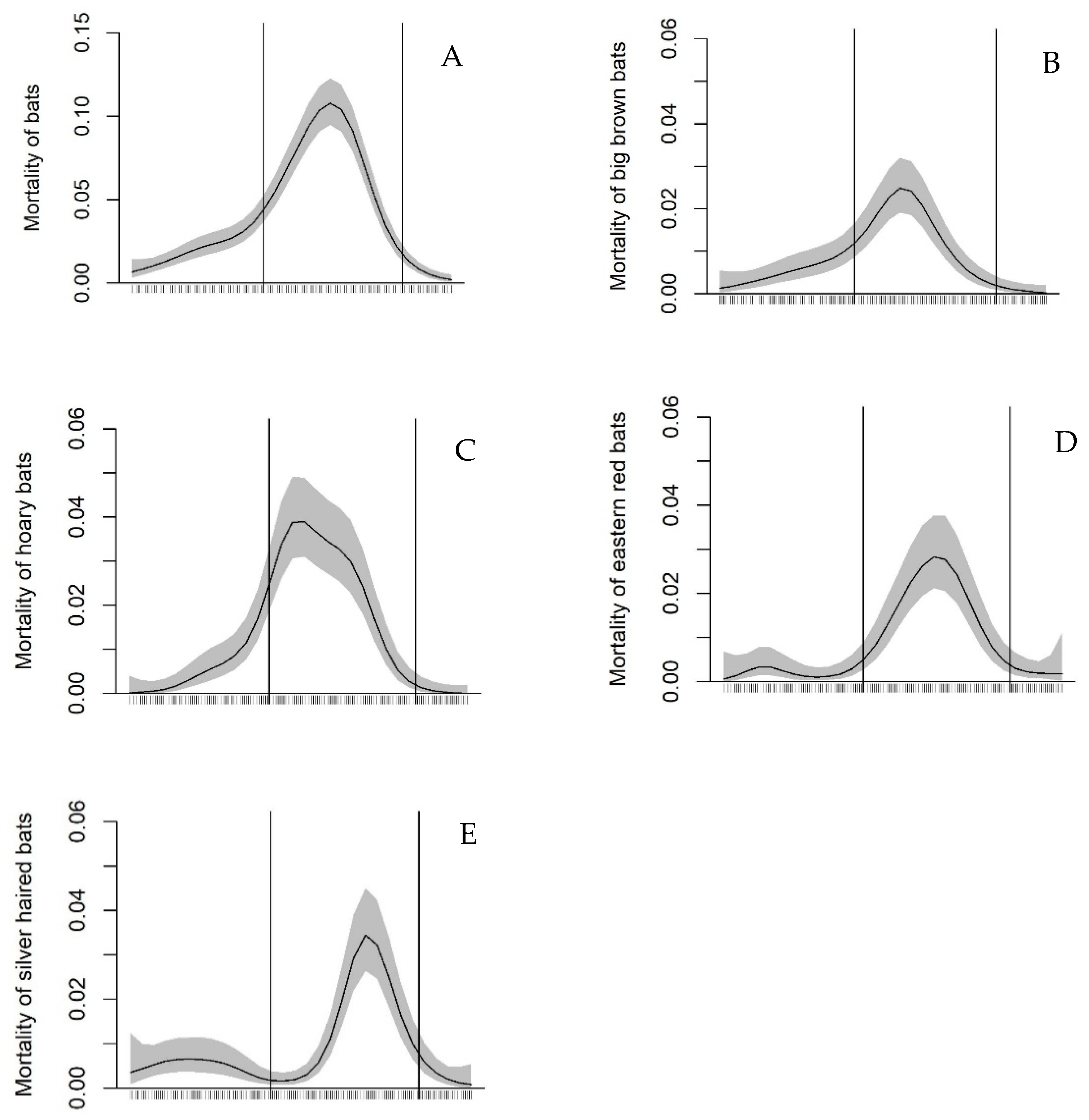
3.1. Relationship of Bat Mortality to Seasonal Time and Weather
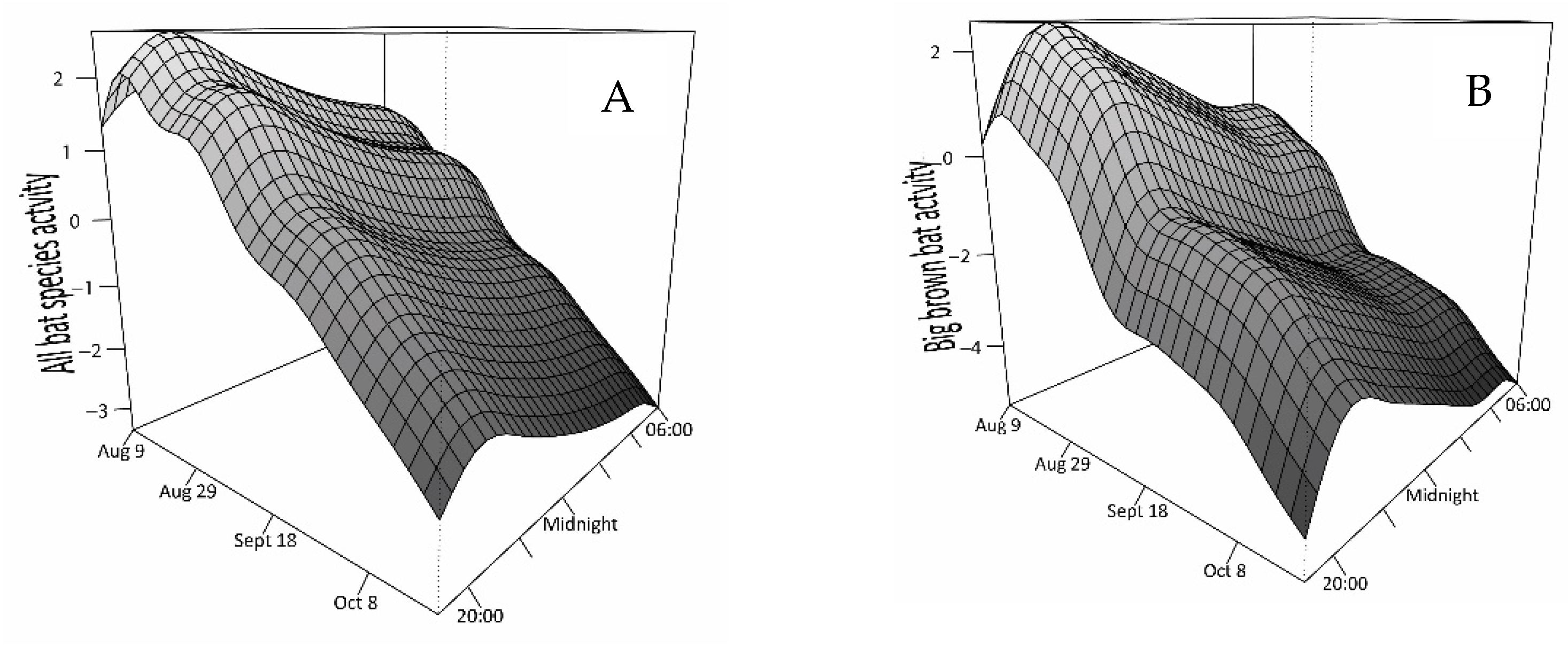
3.2. Relationship of Bat Activity to Time and Weather
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cryan, P.M. Wind turbines as landscape impediments to the migratory connectivity of bats. J. Environ. Law 2011, 41, 355–370. [Google Scholar]
- Arnett, E.B.; Baerwald, E.F. Impacts of wind energy development on bats: Implications for conservation. In Bat Evolution, Ecology, and Conservation; Adams, R.A., Peterson, S.C., Eds.; Springer Science Press: New York, NY, USA, 2013. [Google Scholar]
- Hayes, M.A. Bats killed in large numbers at United States wind energy facilities. BioScience 2013, 63, 975–979. [Google Scholar]
- Zimmerling, J.R.; Francis, C.M. Bat mortality due to wind turbines in Canada. J. Wildl. Manag. 2016, 80, 1360–1369. [Google Scholar] [CrossRef]
- Frick, W.F.; Baerwald, E.F.; Pollock, J.F.; Barclay, R.M.R.; Szymanski, J.A.; Weller, T.J.; Russell, A.L.; Loeb, S.C.; Medellin, R.A.; McGuire, L.P. Fatalities at wind turbines may threaten population viability of a migratory bat. Biol. Conserv. 2017, 209, 172–177. [Google Scholar] [CrossRef]
- Hammerson, G.A.; Kling, M.; Harkness, M.; Ormes, M.; Young, B.E. Strong geographic and temporal patterns in conservation status of North American bats. Biol. Conserv. 2017, 212, 144–152. [Google Scholar] [CrossRef]
- Davy, C.M.; Squires, K.; Zimmerling, J.R. Estimation of spatiotemporal trends in bats abundance from mortality data collected at wind turbines. Conserv. Biol. 2020, 35, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Friedenberg, N.A.; Frick, W.F. Assessing fatality minimization for hoary bats amid continued wind energy development. Biol. Conserv. 2021, 262, 109309. [Google Scholar] [CrossRef]
- Johnson, G.D. A review of bat mortality at wind-energy developments in the United States. Bat Res. News 2005, 46, 45–49. [Google Scholar]
- Arnett, E.B.; Brown, K.; Erickson, W.P.; Fiedler, J.K.; Hamilton, B.L.; Henry, T.L.; Jain, A.; Johnson, G.D.; Kerns, J.; Korford, R.R.; et al. Patterns of fatality of bats at wind energy facilities in North America. J. Wildl. Manag. 2008, 72, 61–78. [Google Scholar] [CrossRef]
- Calvert, A.M.; Bishop, C.A.; Elliot, R.D.; Krebs, E.A.; Kydd, T.M.; Machtans, C.S.; Robertson, G.J. A synthesis of human-related avian mortality in Canada. Avian Conserv. Ecol. 2013, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Zimmerling, J.R.; Pomeroy, A.C.; d’Entremont, M.V.; Francis, C.M. Canadian estimate of bird mortality due to collisions and direct habitat loss associated with wind turbine developments. Avian Conserv. Ecol. 2013, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.S.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mamm. Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Cryan, P.M.; Brown, A.C. Migration of bats past a remote island offers clues toward the problem of bat fatalities at wind turbines. Biol. Conserv. 2007, 139, 1–11. [Google Scholar] [CrossRef]
- Horn, J.W.; Arnett, E.B.; Kunz, T.H. Behavioral responses of bats to operating wind turbines. J. Wildl. Manag. 2008, 72, 123–132. [Google Scholar] [CrossRef]
- Baerwald, E.F.; Edworthy, J.; Holder, M.; Barclay, R.M.R. A large-scale mitigation experiment to reduce bat fatalities at wind energy facilities. J. Wildl. Manag. 2009, 73, 1077–1081. [Google Scholar] [CrossRef]
- Arnett, E.B.; Huso, M.M.P.; Schirmacher, M.R.; Hayes, J.P. Altering turbine speed reduces bat mortality at wind-energy facilities. Front. Ecol. Environ. 2011, 9, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Ontario Ministry of Natural Resources [OMNR]. Bats and Bat Habitats: Guidelines for Wind Power Projects, 2nd ed.; Ontario Ministry of Natural Resources and Forestry: Peterborough, ON, Canada, 2011.
- Arnett, E.B.; Johnson, G.D.; Erickson, W.P.; Hein, C.D. A synthesis of operational mitigation studies to reduce bat fatalities at wind energy facilities in North America. In A Report Submitted to the National Renewable Energy Laboratory; Bat Conservation International: Austin, TX, USA, 2013. [Google Scholar]
- Whitby, M.D.; Schirmacher, M.R.; Frick, W.F. The State of the Science on Operational Minimization to Reduce Bat Fatality at Wind Energy Facilities. In A Report Submitted to National Renewables Energy Lab; Bat Conservation International: Austin, TX, USA, 2021. [Google Scholar]
- Hein, C.D.; Prichard, A.; Mabee, T.; Schirmacher, M.R. Effectiveness of an Operational Mitigation Experiment to Reduce Bat Fatalities at the Pinnacle Wind Farm, Mineral County, West Virginia, 2012. In An Annual Report Submitted to Edison Mission Energy and the Bats and Wind Energy Cooperative; Bat Conservation International: Austin, TX, USA, 2013. [Google Scholar]
- Martin, C.M.; Arnett, E.B.; Stevens, R.D.; Wallace, M.C. Reducing bat fatalities at wind facilities while improving the economic efficiency of operational mitigation. J. Mammal. 2017, 98, 378–385. [Google Scholar] [CrossRef]
- Adams, E.M.; Gulka, J.; Williams, K.A. A review of the effectiveness of operational curtailment for reducing bat fatalities at terrestrial wind farms in North America. PLoS ONE 2021, 16, e0256382. [Google Scholar] [CrossRef]
- North American Clean Energy. Available online: http://www.nacleanenergy.com/articles/30944/bat-smart-curtailment-suitability-assessment-and-modeling (accessed on 1 January 2018).
- Hayes, M.A.; Hooten, L.A.; Gilland, K.L.; Grandgent, C.; Smith, R.L.; Lindsay, S.R.; Collins, J.D.; Schumacher, S.M.; Rabie, P.A.; Gruver, J.C.; et al. A smart curtailment approach for reducing bat fatalities and curtailment time at wind energy facilities. Ecol. Appl. 2019, 29, e01881. [Google Scholar] [CrossRef]
- Young, D.P., Jr.; Nomani, S.; Tidhar, W.L.; Bay, K. NedPower Mount Storm Wind Energy Facility post-construction avian and bat monitoring: July-October 2010. In Report Prepared for NedPower Mount Story; Western EcoSystems Technology, Inc.: Cheyenne, WY, USA, 2011. [Google Scholar]
- Weller, T.J.; Baldwin, J.A. Using echolocation monitoring to model bat occupancy and inform mitigations at wind energy facilities. J. Wildl. Manag. 2012, 76, 619–631. [Google Scholar] [CrossRef]
- Farnsworth, A.; Horton, K.; Heist, K.; Bridge, E.; Diehl, R.; Frick, W.; Kelly, J.; Stepanian, P. The Role of Regional-Scale Weather Variables in Predicting Bat Mortality and Bat Vocalizations: Potential for Use in the Development of Smart Curtailment Algorithms; AWWI Technical Report: Washington, DC, USA, 2021. [Google Scholar]
- Canada Wind Energy Association Homepage. Available online: https://canwea.ca/ (accessed on 1 November 2021).
- Britzke, E.R.; Gillam, E.H.; Murray, K.L. Current state of understanding of ultrasonic detectors for the study of bat ecology. Acta. Theriol. 2013, 58, 109–117. [Google Scholar] [CrossRef]
- Miller, B.W. A method for determining relative activity of free flying bats using a new activity index for acoustic monitoring. Acta. Chiropt. 2001, 3, 93–105. [Google Scholar]
- Kerns, J.; Erickson, W.P.; Arnett, E.B. Bat and bird fatality at wind energy facilities in Pennsylvania and West Virginia. In Relationships between Bats and Wind Turbines in Pennsylvania and West Virginia: An Assessment of Bat Fatality Search Protocols, Patterns of Fatality, and Behavioral Interactions with Wind Turbines; Arnett, E.B., Ed.; A Final Report to the Bats and Wind Energy Cooperative; Bat Conservation International: Austin, TX, USA, 2005; pp. 24–95. [Google Scholar]
- Baerwald, E.F.; Barclay, R.M.R. Patterns of activity and fatality of migratory bats at a wind energy facility in Alberta, Canada. J. Wildl. Manag. 2011, 75, 1103–1114. [Google Scholar] [CrossRef]
- McGuire, L.P.; Guglielmo, C.G.; Mackenzie, S.A.; Taylor, P.D. Migratory stopover in the long-distance migrant silver-haired bat, Lacionycteris noctivagans. J. Anim. Ecol. 2012, 81, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Good, R.E.; Merrill, A.; Simon, S.; Murray, K.L.; Bay, K. Bat Monitoring Studies at the Fowler Ridge Wind Farm, Benton County, Indiana; Final Report: 1 April–31 October, 2011; Prepared for Fowler Ridge Wind Farm, Fowler, Indiana; Western EcoSystems Technology, Inc.: Bloomington, IN, USA, 2012. [Google Scholar]
- Environment and Climate Change Canada Historical Climate Data. Available online: http://climate.weather.gc.ca/ (accessed on 1 January 2017).
- Bañuelos-Ruedas, F.; Angeles-Camacho, C.; Rios-Marcuello, S. Analysis and validation of the methodology used in the extrapolation of wind speed data at different heights. Renew. Sustain. Energy Rev. 2010, 14, 2383–2391. [Google Scholar] [CrossRef]
- Austin, R.H.; Phillips, B.F.; Webb, D.J. A method for calculating moonlight illuminance at the Earth’s surface. J. Appl. Ecol. 1976, 13, 741–748. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, L.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Pedersen, E.J.; Miller, D.L.; Simpson, G.L.; Ross, N. Hierarchical generalized additive models in ecology: An introduction with mgcv. PeerJ 2019, 7, e6876. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Vaida, F.; Blanchard, S. Conditional Akaike information for mixed-effects models. Biometrika 2005, 92, 351–370. [Google Scholar] [CrossRef]
- Arnold, T.W. Uninformative parameters and model selection using Akaike’s Information Criterion. J. Wildl. Manag. 2010, 74, 1175–1178. [Google Scholar] [CrossRef]
- Jameson, J.W.; Willis, C.K.R. Bat mortality at a wind power facility in central Canada. Northwest Nat. 2012, 93, 194–202. [Google Scholar] [CrossRef]
- Smith, A.D.; McWilliams, S.R. Bat activity during autumn relates to atmospheric conditions: Implications for coastal wind energy development. J. Mammal. 2016, 97, 1565–1577. [Google Scholar] [CrossRef]
- Muthersbaugh, M.S.; Ford, W.M.; Powers, K.E.; Silvis, A. Activity patterns of bats during the fall and spring along ridgelines in the central Appalachians. J. Fish Wildl. Manag. 2019, 10, 180–195. [Google Scholar] [CrossRef] [Green Version]
- Cryan, P.M.; Veilleux, J.P. Migration and the use of autumn, winter, and spring roosts by tree bats. In Bats in Forests: Conservation and Management; Lacki, M.J., Ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 153–175. [Google Scholar]
- Voigt, C.C.; Schneeberge, K.; Voigt-Heucke, S.L.; Lewanzik, D. Rain increases the energy cost of bat flight. Biol. Lett. 2011, 7, 793–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burles, D.W.; Brigham, R.M.; Ring, R.A.; Reimchen, T.E. Influence of weather on two insectivorous bats in a temperate Pacific Northwest rainforest. Can. J. Zool. 2009, 87, 132–138. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Bat Species | Variable | Estimate | SE | p |
|---|---|---|---|---|
| All combined | Year | −0.20 | 0.06 | <0.001 |
| Temperature | 0.16 | 0.05 | <0.01 | |
| Rain | −0.10 | 0.06 | 0.08 | |
| Strong southerly wind | −0.12 | 0.06 | 0.04 | |
| Hoary bat | Year | −0.32 | 0.10 | <0.001 |
| Temperature | 0.31 | 0.09 | <0.01 | |
| Wind speed | −0.43 | 0.12 | <0.001 | |
| Big brown bat | Year | −0.35 | 0.10 | <0.01 |
| Temperature | 0.19 | 0.11 | 0.09 | |
| Rain | −0.27 | 0.15 | 0.09 | |
| Pressure | 0.32 | 0.17 | 0.07 | |
| Wind speed | −0.13 | 0.14 | 0.39 | |
| Eastern red bat | Year | −0.07 | 0.12 | 0.67 |
| Rain | −0.25 | 0.14 | 0.08 |
| Bat Species | Variable | Estimate | SE | p |
|---|---|---|---|---|
| All combined | Temperature | 0.32 | 0.03 | <0.001 |
| Rain | −0.77 | 0.09 | <0.001 | |
| Wind speed | −0.34 | 0.03 | <0.001 | |
| Pressure | −0.05 | 0.04 | 0.13 | |
| Hoary bat | Temperature | 0.29 | 0.06 | <0.001 |
| Rain | −0.37 | 0.18 | 0.05 | |
| Wind speed | −0.18 | 0.06 | <0.01 | |
| Moon | −0.05 | 0.05 | 0.36 | |
| Big brown bat | Temperature | 0.90 | 0.05 | <0.001 |
| Rain | −0.61 | 0.13 | <0.001 | |
| Wind speed | −0.33 | 0.04 | <0.001 | |
| Moon | −0.19 | 0.04 | <0.001 | |
| Eastern red bat | Temperature | 0.11 | 0.05 | 0.03 |
| Rain | −0.70 | 0.17 | <0.001 | |
| Wind speed | −0.35 | 0.06 | <0.001 | |
| Moon | −0.18 | 0.05 | <0.001 | |
| Silver-haired bat | Temperature | −0.06 | 0.04 | 0.12 |
| Rain | −0.80 | 0.14 | <0.001 | |
| Wind speed | −0.39 | 0.04 | <0.001 | |
| Moon | 0.03 | 0.04 | 0.43 | |
| Pressure | −0.18 | 0.05 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Squires, K.A.; Thurber, B.G.; Zimmerling, J.R.; Francis, C.M. Timing and Weather Offer Alternative Mitigation Strategies for Lowering Bat Mortality at Wind Energy Facilities in Ontario. Animals 2021, 11, 3503. https://doi.org/10.3390/ani11123503
Squires KA, Thurber BG, Zimmerling JR, Francis CM. Timing and Weather Offer Alternative Mitigation Strategies for Lowering Bat Mortality at Wind Energy Facilities in Ontario. Animals. 2021; 11(12):3503. https://doi.org/10.3390/ani11123503
Chicago/Turabian StyleSquires, Kelly A., Bethany G. Thurber, J. Ryan Zimmerling, and Charles M. Francis. 2021. "Timing and Weather Offer Alternative Mitigation Strategies for Lowering Bat Mortality at Wind Energy Facilities in Ontario" Animals 11, no. 12: 3503. https://doi.org/10.3390/ani11123503
APA StyleSquires, K. A., Thurber, B. G., Zimmerling, J. R., & Francis, C. M. (2021). Timing and Weather Offer Alternative Mitigation Strategies for Lowering Bat Mortality at Wind Energy Facilities in Ontario. Animals, 11(12), 3503. https://doi.org/10.3390/ani11123503