
Spatial Behavior and Habitat Use of Two Sympatric Bat Species



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
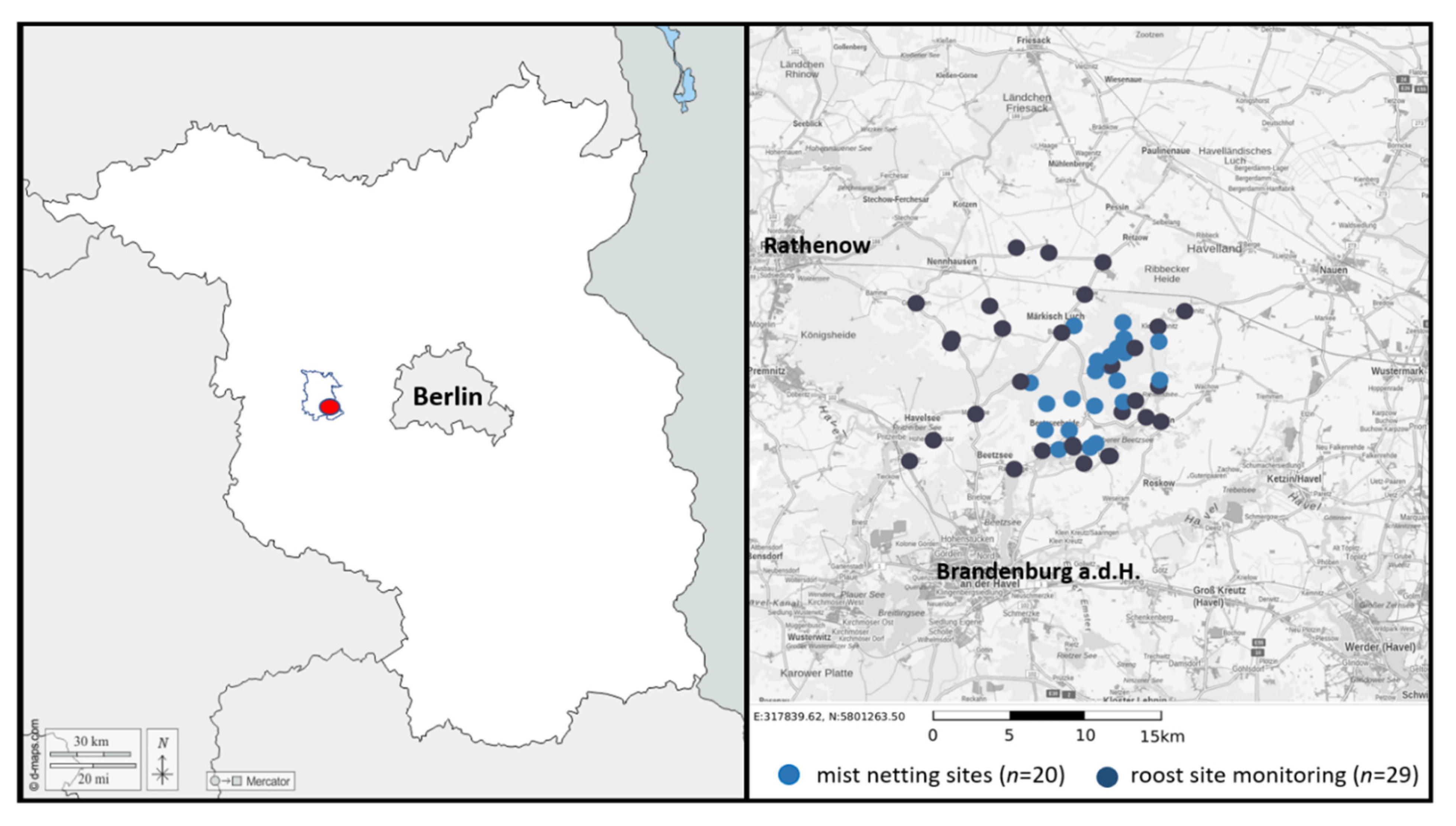
2.1. Study Area
2.2. Data Collection
2.2.1. Species Occurrence and Phenotypic Distinction
2.2.2. Phenology, Activity and Emergence Behavior
2.2.3. Reconstruction of the Prey Spectrum
2.2.4. Habitat Use and Foraging Behavior
3. Results
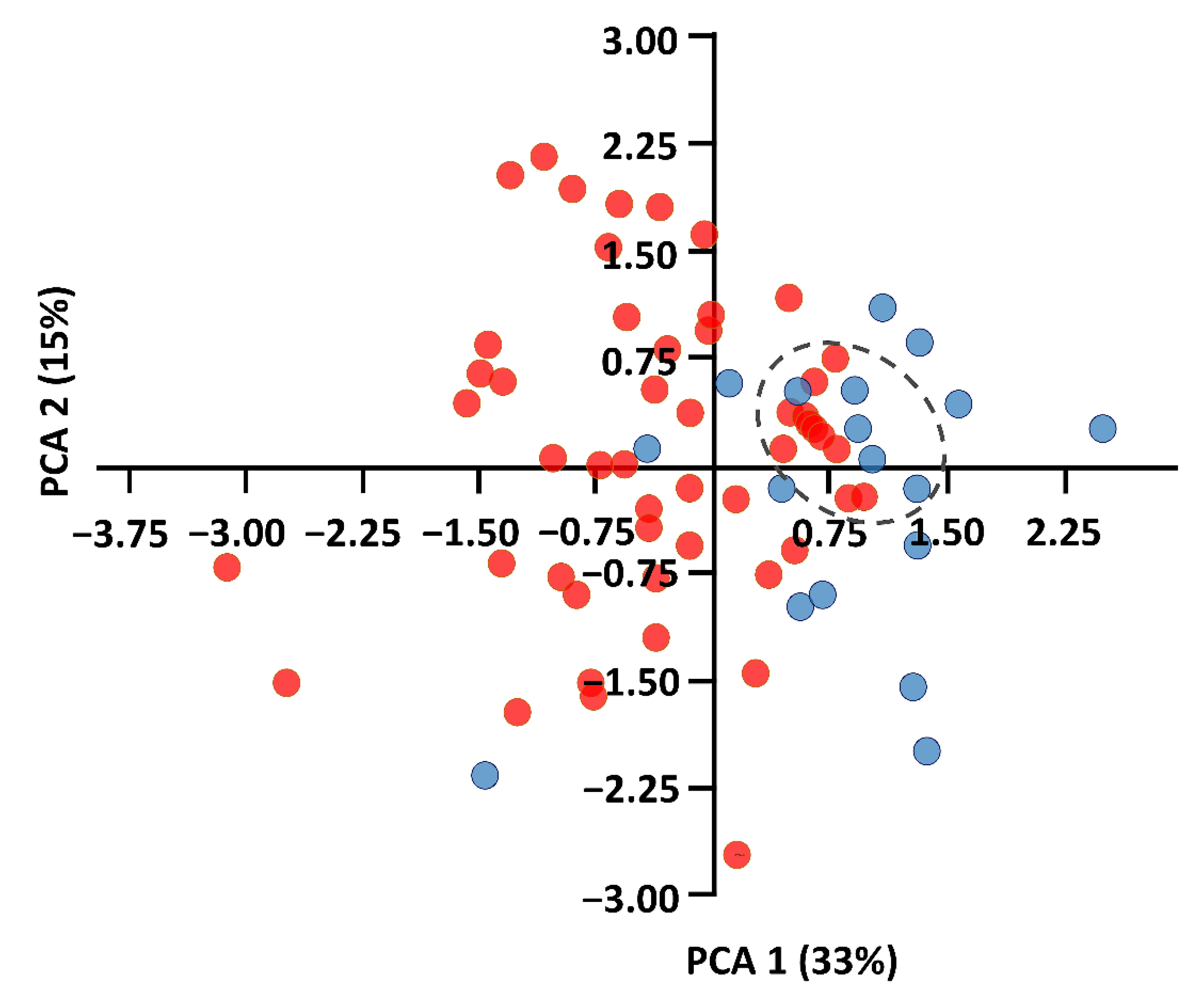
3.1. Comparison of External Characteristics
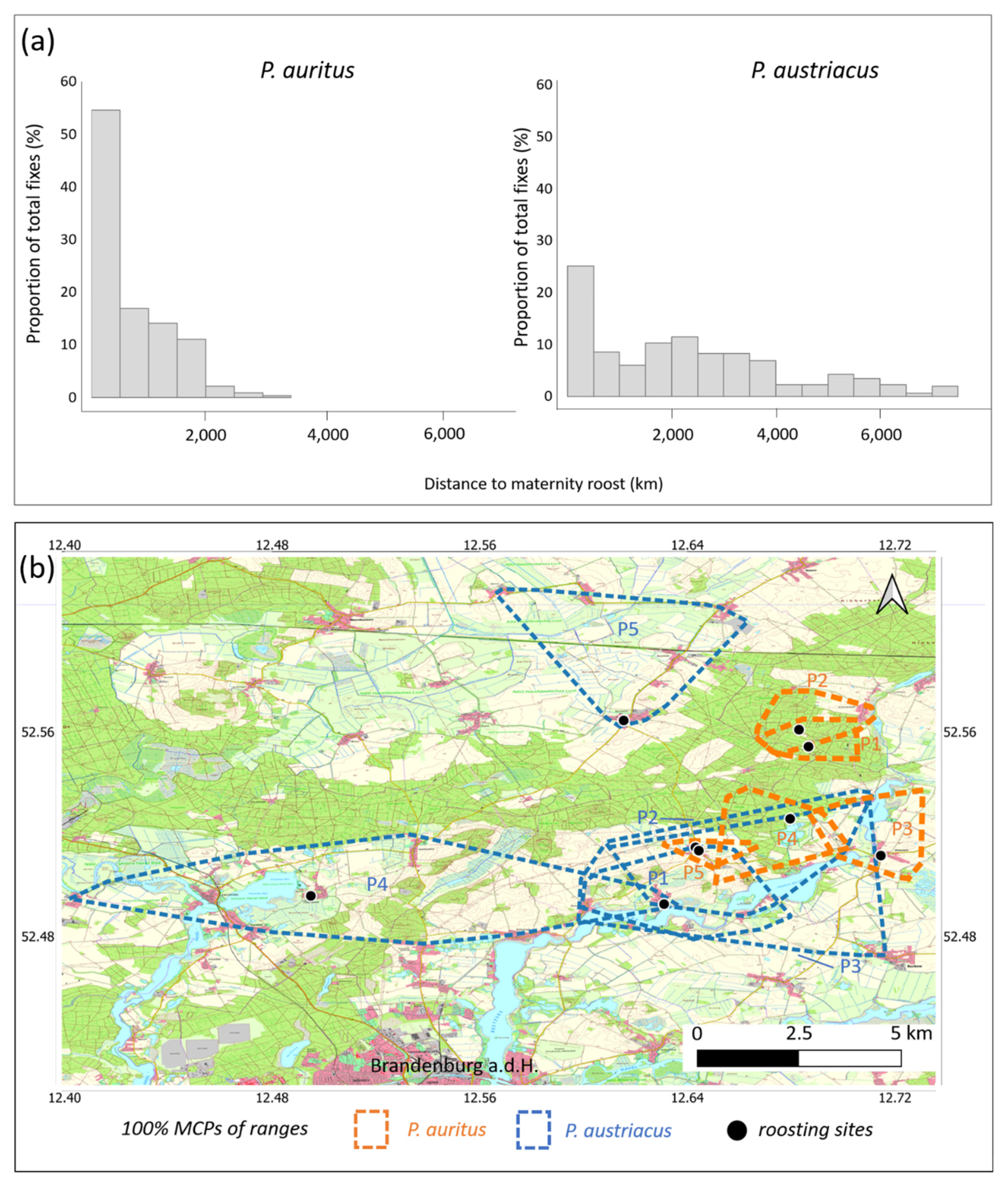
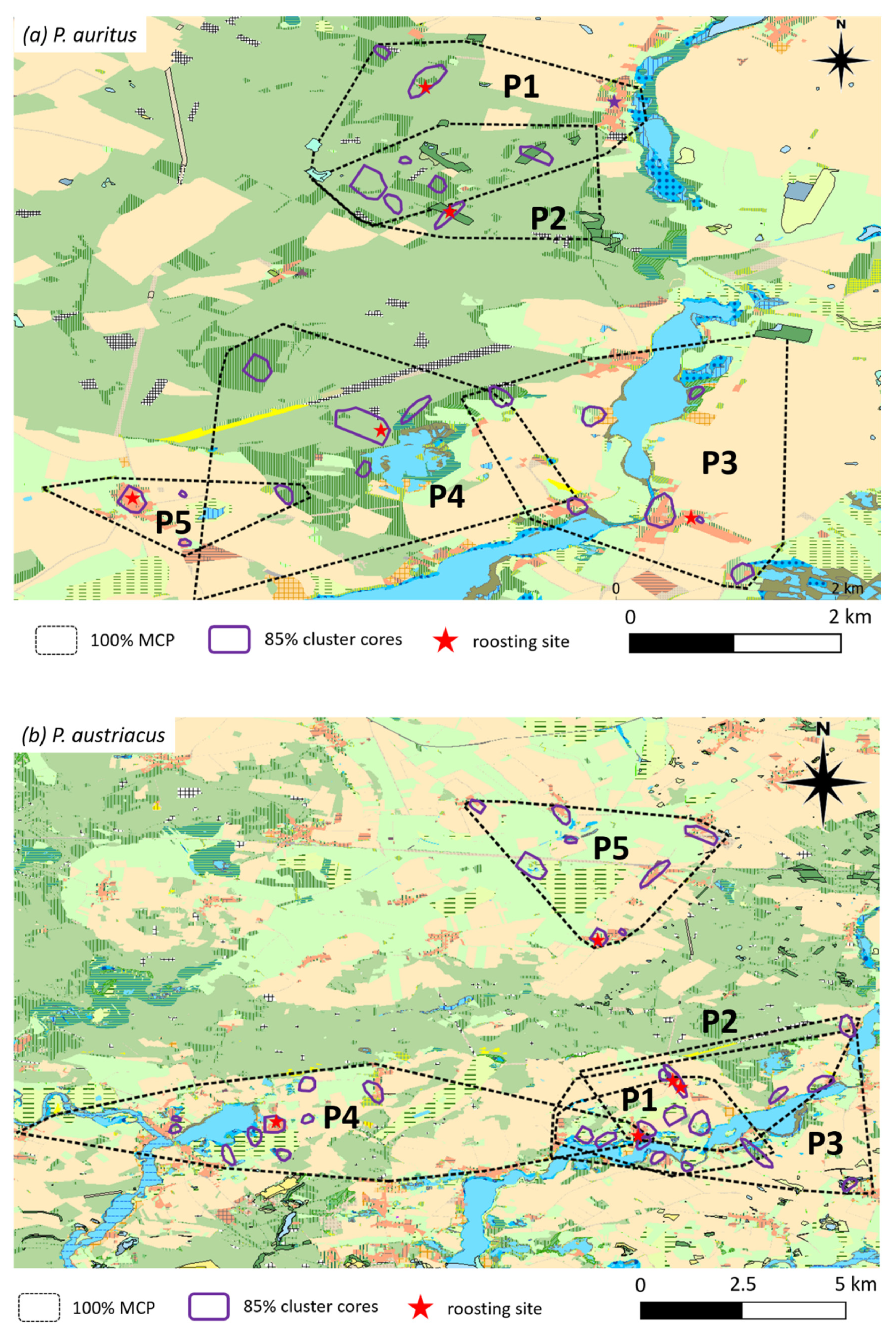
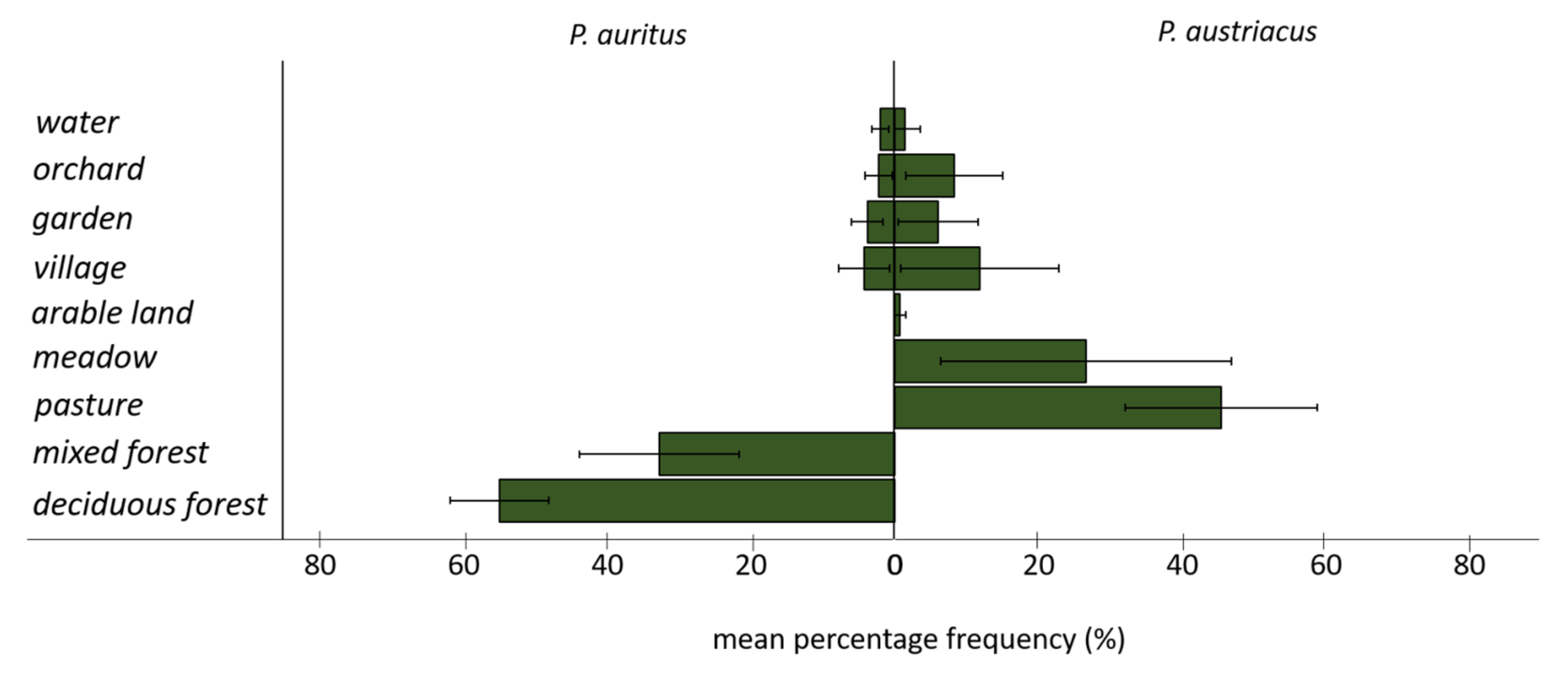
3.2. Spatial Behavior and Habitat Use
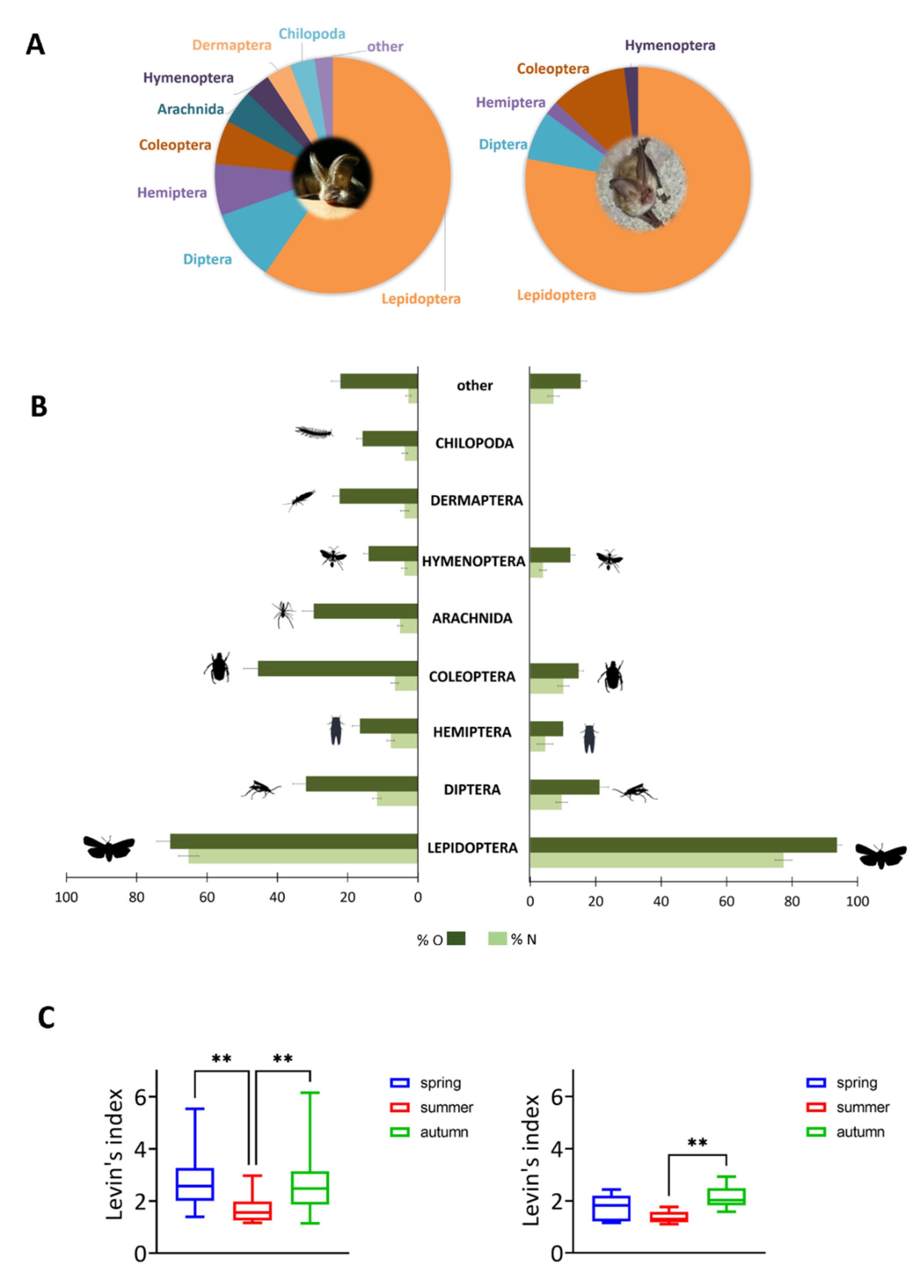
3.3. Reconstruction of the Prey Spectrum
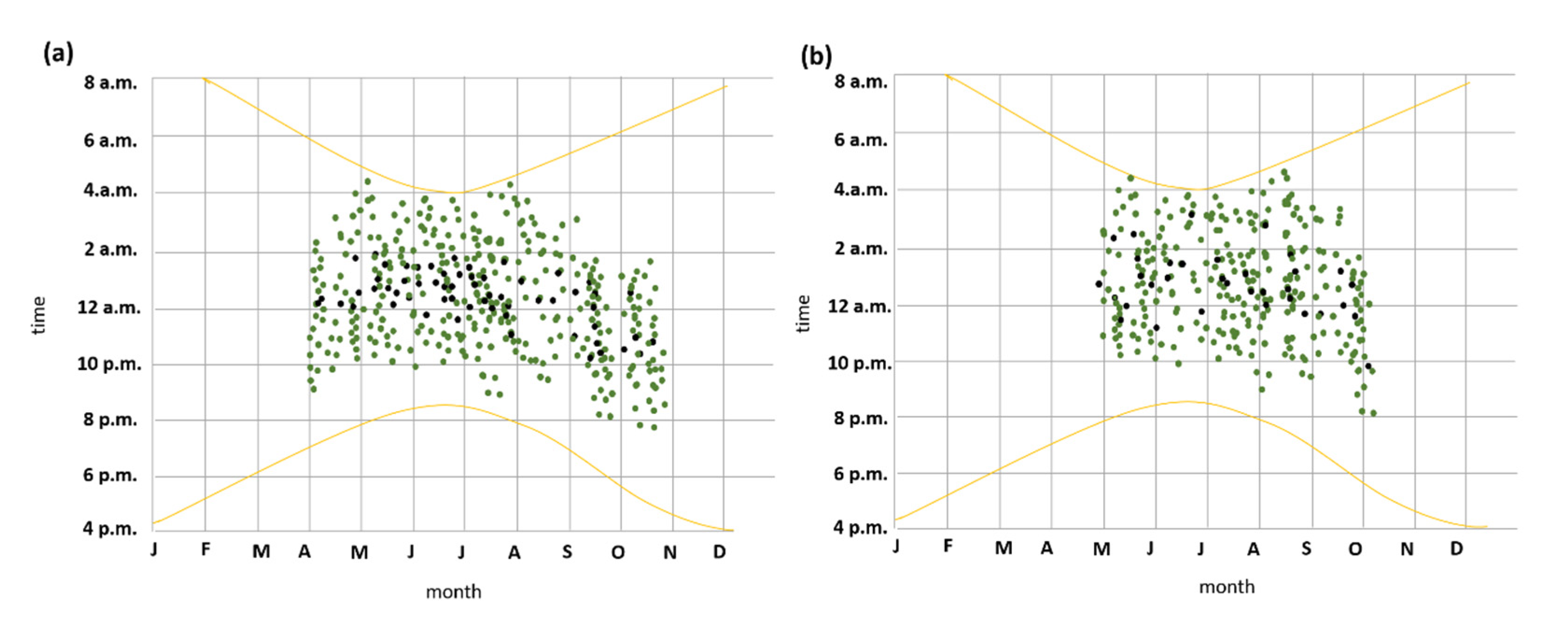
3.4. Seasonal and Nocturnal Activity Patterns
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, G.; Holderied, M.W. Bat echolocation calls: Adaptation and convergent evolution. Proc. R. Soc. B Biol. Sci. 2007, 274, 905–912. [Google Scholar] [CrossRef]
- Salinas-Ramos, V.B.; Ancillotto, L.; Bosso, L.; Sánchez-Cordero, V.; Russo, D. Interspecific competition in bats: State of knowledge and research challenges. Mammal Rev. 2020, 50, 68–81. [Google Scholar] [CrossRef]
- Peixoto, F.P.; Braga, P.H.P.; Mendes, P. A synthesis of ecological and evolutionary determinants of bat diversity across spatial scales. BMC Ecol. 2018, 18, 18. [Google Scholar] [CrossRef]
- Siemers, B.M.; Greif, S.; Borissov, I.; Voigt-Heucke, S.L.; Voigt, C.C. Divergent trophic levels in two cryptic sibling bat species. Oecologia 2011, 166, 69–78. [Google Scholar] [CrossRef]
- Razgour, O.; Clare, E.L.; Zeale, M.R.; Hanmer, J.; Schnell, I.B.; Rasmussen, M.; Gilbert, T.B.; Jones, G. High-throughput sequencing offers insight into mechanisms of resource partitioning in cryptic bat species. Ecol. Evol. 2011, 1, 556–570. [Google Scholar] [CrossRef]
- Ashrafi, S.; Beck, A.; Rutishauser, M.; Arlettaz, R.; Bontadina, F. Trophic niche partitioning of cryptic species of long-eared bats in Switzerland: Implications for conservation. Eur. J. Wildl. Res. 2011, 57, 843–849. [Google Scholar] [CrossRef]
- Sattler, T.; Bontadina, F.; Hirzel, A.H.; Arlettaz, R. Ecological niche modelling of two cryptic bat species calls for a reassessment of their conservation status. J. Appl. Ecol. 2007, 44, 1188–1199. [Google Scholar] [CrossRef]
- Weber, M.G.; Strauss, S.Y. Coexistence in close relatives: Beyond competition and reproductive isolation in sister taxa. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 359–381. [Google Scholar] [CrossRef]
- Reif, J.; Reifová, R.; Skoracka, A.; Kuczyński, L. Competition-driven niche segregation on a landscape scale: Evidence for escaping from syntopy towards allotopy in two coexisting sibling passerine species. J. Anim. Ecol. 2018, 87, 774–789. [Google Scholar] [CrossRef] [PubMed]
- Hardin, G. The competitive exclusion principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Schoener, T.W. Resource partitioning in ecological communities. Science 1974, 185, 27–39. [Google Scholar] [CrossRef]
- Capitán, J.A.; Cuenda, S.; Alonso, D. How similar can co-occurring species be in the presence of competition and ecological drift? J. R. Soc. Interface 2015, 12, 20150604. [Google Scholar] [CrossRef] [PubMed]
- Amarasekare, P.; Hoopes, M.F.; Mouquet, N.; Holyoak, M. Mechanisms of coexistence in competitive metacommunities. Am. Nat. 2004, 164, 310–326. [Google Scholar] [CrossRef]
- Basile, M.; Asbeck, T.; Cordeiro Pereira, J.M.; Mikusiński, G.; Storch, I. Species co-occurrence and management intensity modulate habitat preferences of forest birds. BMC Biol. 2021, 19, 210. [Google Scholar] [CrossRef]
- Spitzenberger, F.; Strelkov, P.P.; Winkler, H.; Haring, E. A preliminary revision of the genus Plecotus (Chiroptera, Vespertilionidae) based on genetic and morphological results. Zool. Scr. 2006, 35, 187–230. [Google Scholar] [CrossRef]
- Kiefer, A.; Mayer, F.; Kosuch, J.; Von Helversen, O.; Veith, M. Conflicting molecular phylogenies of European long-eared bats (Plecotus) can be explained by cryptic diversity. Mol. Phylogenet. Evol. 2002, 25, 557–566. [Google Scholar] [CrossRef]
- Benda, P.; Kiefer, A.; Hanák, V.; Veith, M. Systematic status of African populations of long-eared bats, genus Plecotus (Mammalia: Chiroptera). Folia Zool. 2004, 53, 47. [Google Scholar]
- Mayer, F.; Dietz, C.; Kiefer, A. Molecular species identification boots bat diversity. Front. Zool. 2007, 4, 4. [Google Scholar] [CrossRef][Green Version]
- Kiefer, A.; Veith, M. A new species of long-eared bat from Europe (Chiroptera: Vespertilionidae). Myotis 2001, 39, 5–16. [Google Scholar]
- Spitzenberger, F.; Strelkov, P.; Haring, E. Morphology and mitochondrial DNA sequences show that Plecotus alpinus Kiefer & Veith, 2002 and Plecotus microdontus Spitzenberger, 2002 are synonyms of Plecotus macrobullaris Kuzjakin, 1965. Nat. Croat. 2003, 12, 39. [Google Scholar]
- Ashrafi, S.; Bontadina, F.; Beck, A.; Pavlinić, I.; Arlettaz, R. Multiple morphological characters needed for field identification of cryptic long-eared bat species around the Swiss Alps. J. Zool. 2010, 281, 241–248. [Google Scholar] [CrossRef]
- Preatoni, D.G.; Spada, M.; Wauters, L.A.; Tosi, G.; Martinoli, A. Habitat use in the female Alpine long-eared bat (Plecotus macrobullaris): Does breeding make the difference? Acta Chiropt. 2011, 13, 355–364. [Google Scholar] [CrossRef]
- Mucedda, M.; Kiefer, A.; Pidinchedda, E.; Veith, M. A new species of long-eared bat (Chiroptera, Vespertilionidae) from Sardinia (Italy). Acta Chiropt. 2002, 4, 121–135. [Google Scholar] [CrossRef]
- Juste, J.; Ibáñez, C.; Muñoz, J.; Trujillo, D.; Benda, P.; Karataş, A.; Ruedi, M. Mitochondrial phylogeography of the long-eared bats (Plecotus) in the Mediterranean Palaearctic and Atlantic Islands. Mol. Phylogenet. Evol. 2004, 31, 1114–1126. [Google Scholar] [CrossRef] [PubMed]
- Ancillotto, L.; Bosso, L.; Smeraldo, S.; Mori, E.; Mazza, G.; Herkt, M.; Galimberti, A.; Ramazotti, F.; Russo, D. An African bat in Europe, Plecotus gaisleri: Biogeographic and ecological insights from molecular taxonomy and species distribution models. Ecol. Evol. 2020, 10, 5785–5800. [Google Scholar] [CrossRef] [PubMed]
- Stebbings, R.E. Identification and distribution of bats of the genus Plecotus in England. J. Zool. 1967, 153, 291–310. [Google Scholar] [CrossRef]
- Swift, S.M. Long-Eared Bats; T. & A.D. Poyser Ltd.: Manchester, UK, 1998; p. 182. [Google Scholar]
- Starik, N.; Göttert, T.; Heitlinger, E.; Zeller, U. Bat community responses to structural habitat complexity resulting from management practices within different land use types—A case study from north-eastern Germany. Acta Chiropt. 2018, 20, 387–405. [Google Scholar] [CrossRef]
- Zeller, U.; Starik, N.; Göttert, T. Biodiversity, land use and ecosystem services—An organismic and comparative approachto different geographical regions. Glob. Ecol. Conserv. 2017, 10, 114–125. [Google Scholar] [CrossRef]
- Zeller, U.; Perry, G.; Starik, N.; Göttert, T. Organisms in Nature—Evolutionary Perspective and Ecological Significance; Humboldt-Universität zu Berlin: Berlin, Germany, 2020; p. 55. [Google Scholar]
- Dietz, C.; von Helversen, O. Illustrated Identification Key to the Bats of Europe; Electronic Publication: Tuebingen & Erlangen, Germany, 2004; pp. 62–67. [Google Scholar]
- Norberg, U.M.; Rayner, J.M. Ecological morphology and flight in bats (Mammalia; Chiroptera): Wing adaptations, flight performance, foraging strategy and echolocation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1987, 316, 335–427. [Google Scholar]
- Blood, B.R.; McFarlane, D.A. A new method for calculating the wing area of bats. Mammalia 1988, 52, 600–603. [Google Scholar] [CrossRef][Green Version]
- Dwyer, P.D. The breeding biology of Miniopterus schreibersi blepotis (Termminck)(Chiroptera) in north-eastern NSW. Aust. J. Zool. 1988, 11, 219–240. [Google Scholar] [CrossRef]
- Jones, C.; Suttkus, R.D. Wing loading in Plecotus rafinesquii. J. Mammal. 1971, 52, 458–460. [Google Scholar] [CrossRef]
- Nicoll, M.E.; Suttie, J.M. The sheath-tailed bat Coleura seychellensis (Chiroptera: Emballonuridae) in the Seychelles islands. J. Zool. 1982, 197, 421–426. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing tables of statistical test. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Barataud, M. Écologie acoustique des chiroptères d‘Europe. In Identification des Espèces, Études de Leurs Habitats et Comportements de Chasse; Biotope Editions Mèze; Muséum National d’Histoire Naturelle: Paris, France, 2012; p. 344. [Google Scholar]
- Obrist, M.K.; Boesch, R.; Flückiger, P.F. Variability in echolocation call design of 26 Swiss bat species: Consequences, limits and options for automated field identification with a synergetic pattern recognition approach. Mammalia 2004, 68, 307–322. [Google Scholar] [CrossRef]
- Skiba, R. Fledermäuse: Kennzeichen, Echoortung und Detektoranwendung, 2nd ed.; Die Neue Brehm-Bücherei VerlagsKG Wolf: Magedeburg, Germany, 2009. [Google Scholar]
- Russ, J. British Bat Calls: A Guide to Species Identification; Pelagic Publishing Ltd.: Exeter, UK, 2012. [Google Scholar]
- Entwistle, A. Roost Ecology of the Brown Long-Eared Bat (Plecotus auritus, Linnaeus 1758) in North-East Scotland. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 1994. [Google Scholar]
- Battersby, J.E. A Comparison of the Roost Ecology of the Brown Long-Eared Bat Plecotus auritus and the Serotine Bat Eptesicus serotinus. Ph.D. Thesis, University of Sussex, Brighton, UK, 1999. [Google Scholar]
- Beck, A. Fecal analyses of European bat species. Myotis 1995, 32, 109–119. [Google Scholar]
- Whitaker, J.O. Food habits analysis of insectivorous bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 171–189. [Google Scholar]
- McAney, C.M.; Shiel, C.; Sullivan, C.; Fairley, J. The Analysis of Bat Droppings; Occasional Publications of the Mammal Society 14; Mammal Society: London, UK, 1991. [Google Scholar]
- Shiel, C.B.; McAney, C.M.; Fairley, J.S. Analysis of the diet of Natterer’s bat Myotis nattereri and the common long-eared bat Plecotus auritus in the West of Ireland. J. Zool. 1991, 223, 299–305. [Google Scholar] [CrossRef]
- Obrtel, R.; Holišová, V. Trophic niches of Apodemus flavicollis and Clethrionomys glareolus in a lowland forest. Acta Sci. Nat. 1974, 8, 1–37. [Google Scholar]
- Bauerová, Z. Contribution to the trophic bionomics of Myotis emarginatus. Folia Zool. 1986, 35, 305–310. [Google Scholar]
- Krebs, C.J. Ecological Methodology, 2nd ed.; Addison-Welsey Educational Publishers Inc.: Menlo Park, CA, USA, 1999. [Google Scholar]
- Jones, G.; Morton, M. Radio-tracking studies on habitat use by greater horseshoe bats (Rhinolophus ferrumequinum). In Wildlife Telemetry. Remote Monitoring and Tracking of Animals; Priede, I.G., Swift, S.M., Eds.; Ellis Horwood Ltd.: Chichester, UK, 1992; pp. 521–537. [Google Scholar]
- Duvergé, P.L. Foraging Activity, Habitat Use, Development of Juveniles, and Diet of the Greater Horseshoe Bat (Rhinolophus ferrumequinum–Schreber 1774) in South-West England. Ph.D. Thesis, University of Bristol, Bristol, UK, 1996. [Google Scholar]
- O’Donnell, C.F. Home range and use of space by Chalinolobus tuberculatus, a temperate rainforest bat from New Zealand. J. Zool. 2001, 253, 253–264. [Google Scholar] [CrossRef]
- Van Winkle, W. Comparison of several probabilistic home-range models. J. Wildl. Manag. 1975, 39, 118–123. [Google Scholar] [CrossRef]
- Harris, S.; Cresswell, W.J.; Forde, P.G.; Trewhella, W.J.; Woollard, T.; Wray, S. Home-range analysis using radio-tracking data—A review of problems and techniques particularly as applied to the study of mammals. Mammal Rev. 1990, 20, 97–123. [Google Scholar] [CrossRef]
- Kenward, R.E. Ranging behaviour and population dynamics of grey squirrels. In Behavioural Ecology; Sibly, R.M., Smith, R.H., Eds.; Blackwell Scientific: Oxford, UK, 1985; pp. 319–330. [Google Scholar]
- Kenward, R.E. A Manual for Wildlife Radio Tagging; Academic Press: London, UK, 2000. [Google Scholar]
- Calhoun, J.B.; Casby, J.U. Calculation of home range and density of small mammals. Public Health Monogr. 1958, 55, 1–24. [Google Scholar] [PubMed]
- Jennrich, R.I.; Turner, F.B. Measurement of non-circular home range. J. Theor. Biol. 1969, 22, 27–237. [Google Scholar] [CrossRef]
- Worton, B.J. Using Monte Carlo simulation to evaluate kernel-based home range estimators. J. Wildl. Manag. 1995, 59, 794–800. [Google Scholar] [CrossRef]
- Kenward, R.E.; Clarke, R.T.; Hodder, K.H.; Walls, S.S. Density and linkage estimators of home range: Nearest-neighbor clustering defines multinuclear cores. Ecology 2001, 82, 1905–1920. [Google Scholar] [CrossRef]
- Davidson-Watts, I.; Jones, G. Differences in foraging behaviour between Pipistrellus pipistrellus (Schreber, 1774) and Pipistrellus pygmaeus (Leach, 1825). J. Zool. 2006, 268, 5. [Google Scholar] [CrossRef]
- Entwistle, A.C.; Racey, P.A.; Speakman, J.R. Habitat exploitation by a gleaning bat, Plecotus auritus. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 921–931. [Google Scholar]
- Kiefer, A.; Veith, M. Untersuchungen zu Raumbedarf und Interaktion von Populationen des Grauen Langohrs, Plecotus austriacus (Fischer, 1829), im Nahegebiet. Nyctalus Neue Folge 1998, 6, 531. [Google Scholar]
- Ashrafi, S.; Rutishauser, M.; Ecker, K.; Obrist, M.K.; Arlettaz, R.; Bontadina, F. Habitat selection of three cryptic Plecotus bat species in the European Alps reveals contrasting implications for conservation. Biodivers. Conserv. 2013, 22, 2751–2766. [Google Scholar] [CrossRef]
- Bogdanowicz, W.; Fenton, M.B.; Daleszczyk, K. The relationships between echolocation calls, morphology and diet in insectivorous bats. J. Zool. 1999, 247, 381–393. [Google Scholar] [CrossRef]
- Findley, J.S.; Studier, E.H.; Wilson, D.E. Morphologic properties of bat wings. J. Mammal. 1972, 53, 429–444. [Google Scholar] [CrossRef]
- Van Valen, L. Morphological variation and width of ecological niche. Amer. Natur. 1965, 99, 377–390. [Google Scholar] [CrossRef]
- Siemers, B.M.; Swift, S.M. Differences in sensory ecology contribute to resource partitioning in the bats Myotis bechsteinii and Myotis nattereri (Chiroptera: Vespertilionidae). Behav. Ecol. Sociobiol. 2006, 59, 373–380. [Google Scholar] [CrossRef]
- Aldridge, H. Manoeuvrability and ecological segregation in the little brown (Myotis lucifugus) and Yuma (M. yumanensis) bats (Chiroptera: Vespertilionidae). Can. J. Zool. 1986, 64, 1878–1882. [Google Scholar] [CrossRef]
- Jones, G.; Morton, M.; Hughes, P.M.; Budden, R.M. Echolocation, flight morphology and foraging strategies of some West African hipposiderid bats. J. Zool. 1993, 230, 385–400. [Google Scholar] [CrossRef]
- Fuhrmann, M.; Seitz, A. Nocturnal activity of the brown long-eared bat (Plecotus auritus L.1758): Data from radio-tracking in the Lenneburg forest near Mainz (Germany). In Wildlife Telemetry: Remote Monitoring and Tracking of Animals; Priede, I.G., Swift, S.M., Eds.; Ellis Horwood Ltd.: Chichester, UK, 1992; pp. 538–548. [Google Scholar]
- Barataud, M. Eléments sur le comportement alimentaire des Oreillards brun et gris, Plecotus auritus (Linnaeus, 1758) et Plecotus austriacus (Fischer, 1829). Le Rhinolophe 1990, 7, 3–10. [Google Scholar]
- Burland, T.M.; Barratt, E.M.; Beaumont, M.A.; Racey, P.A. Population genetic structure and gene flow in a gleaning bat, Plecotus auritus. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1999, 266, 975–980. [Google Scholar] [CrossRef]
- Flückiger, P.F.; Beck, A. Observations on the habitat use for hunting by Plecotus austriacus (Fischer, 1829). Myotis 1995, 32, 121–122. [Google Scholar]
- Hutterer, R.; Ivanova, T.; Meyer-Cords, C.; Rodrigues, L. Bat Migration in Europe: A Review of Banding Data and Literature; Naturschutz und Biologische Vielfalt: Münster, Germany, 2005; Volume 28. [Google Scholar]
- Adams, R.A.; Thibault, K.M. Temporal resource partitioning by bats at water holes. J. Zool. 2006, 270, 466–472. [Google Scholar] [CrossRef]
- Swift, S.M.; Racey, P.A. Resource partitioning in two species of vespertilionid bats (Chiroptera) occupying the same roost. J. Zool. 1983, 200, 249–259. [Google Scholar] [CrossRef]
- Catto, C.M.C.; Hutson, A.M.; Raccey, P.A.; Stephenson, P.J. Foraging behaviour and habitat use of the serotine bat (Eptesicus serotinus) in southern England. J. Zool. 1996, 238, 623–633. [Google Scholar] [CrossRef]
- Belwood, J.J.; Fenton, M.B. Variation in the diet of Myotis lucifugus (Chiroptera: Vespertilionidae). Can. J. Zool. 1976, 54, 1674–1678. [Google Scholar] [CrossRef]
- Fenton, M.B.; Morris, G.K. Opportunistic feeding by desert bats (Myotis spp.). Can. J. Zool. 1976, 54, 526–530. [Google Scholar] [CrossRef]
- Swift, S.M.; Racey, P.A.; Avery, M.I. Feeding ecology of Pipistrellus pipistrellus (Chiroptera: Vespertilionidae) during pregnancy and lactation. II. Diet. J. Anim. Ecol. 1985, 54, 217–225. [Google Scholar] [CrossRef]
- Rydell, J. Feeding territoriality in female northern bats, Eptesicus nilssoni. Ethology 1986, 72, 329–337. [Google Scholar] [CrossRef]
- Swift, S.; Racey, P. Gleaning as a foraging strategy in Natterer’s bat Myotis nattereri. Behav. Ecol. Sociobiol. 2002, 52, 408–416. [Google Scholar]
- Whitaker, J.O.; Karataş, A. Food and feeding habits of some bats from Turkey. Acta Chiropt. 2009, 11, 393–403. [Google Scholar]
- Andriollo, T.; Michaux, J.R.; Ruedi, M. Food for everyone: Differential feeding habits of cryptic bat species inferred from DNA metabarcoding. Mol. Ecol. 2021, 30, 4584–4600. [Google Scholar] [CrossRef] [PubMed]
- Fenton, M.B. Science and the conservation of bats. J. Mammal. 1997, 78, 1–14. [Google Scholar] [CrossRef]
- Swartz, S.M.; Freeman, P.W.; Stockwell, E.F. Ecomorphology of bats: Comparative and experimental approaches relating structural design to ecology. In Bat Ecology; Kunz, T.H., Fenton, M.B., Eds.; The University of Chicago Press: Chicago, IL, USA, 2003; pp. 257–300. [Google Scholar]
- Adams, R.A. Size-specific resource use in juvenile little brown bats, Myotis lucifugus (Chiroptera: Vespertilionidae): Is there an ontogenetic shift? Can. J. Zool. 1996, 74, 1204–1210. [Google Scholar] [CrossRef]
- Aldridge, H.D.J.N.; Rautenbach, I.L. Morphology, echolocation and resource partitioning in insectivorous bats. J. Anim. Ecol. 1987, 56, 763–778. [Google Scholar] [CrossRef]
- Brigham, R.M.; Grindal, S.D.; Firman, M.C.; Morissette, J.L. The influence of structural clutter on activity patterns of insectivorous bats. Can. J. Zool. 1997, 75, 131–136. [Google Scholar] [CrossRef]
- Stockwell, E.F. Morphology and flight manoeuvrability in New World leaf-nosed bats (Chiroptera: Phyllostomidae). J. Zool. 2001, 254, 505–514. [Google Scholar] [CrossRef]
- Kalcounis, M.C.; Brigham, R.M. Intraspecific variation in wing loading affects habitat use by little brown bats (Myotis lucifugus). Can. J. Zool. 1995, 73, 89–95. [Google Scholar] [CrossRef]
- Saunders, M.B.; Barclay, R.M. Ecomorphology of insectivorous bats: A test of predictions using two morphologically similar species. Ecology 1992, 73, 1335–1345. [Google Scholar] [CrossRef]
- Rhodes, M.P. Assessment of sources of variance and patterns of overlap in microchiropteran wing morphology in southeast Queensland, Australia. Can. J. Zool. 2002, 80, 450–460. [Google Scholar] [CrossRef]
- Horácek, I.; Bogdanowicz, W.; Dulid, B. Plecotus austriacus (Fischer, 1829)—Graues Langohr. In Handbuch der Säugetiere Europas, 4th ed.; Krapp, F., Ed.; Aula-Verlag: Wiebesheim, Germany, 2004; pp. 1001–1049. [Google Scholar]
- Rudolf, B.U. Graues Langohr Plecotus austriacus (Fischer 1829). In Fledermäuse in Bayern; Meschede, A., Rudolph, B.U., Eds.; Eugen Ulmer: Stuttgart, Germany, 2004; pp. 333–339. [Google Scholar]
- Razgour, O.; Whitby, D.; Dahlberg, E.; Barlow, K.; Hanmer, J.; Haysom, K.; McFarlane, H.; Wicks, L.; Williams, C.; Jones, G. Conserving Grey Long-Eared Bats (Plecotus austriacus) in Our Landscape: A Conservation Management Plan; Bat Conservation Trust: London, UK, 2013; p. 36. [Google Scholar]
- Mathews, F.; Kubasiewicz, L.M.; Gurnell, J.; Harrower, C.A.; McDonald, R.A.; Shore, R.F. A Review of the Population and Conservation Status of British Mammals; A report by the mammal society under contract to Natural England, Natural Resources Wales and Scottish Natural Heritage; Natural England: Peterborough, UK, 2018.
- Gazaryan, S.; Godlevska, L. Plecotus austriacus (amended version of 2020 assessment). The IUCN Red List of Threatened Species. IUCN 2021, e.T85533333A195862345. Available online: https://www.iucnredlist.org/species/85533333/195862345 (accessed on 2 November 2021).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. auritus | P. austriacus | |
|---|---|---|
| Activity area total (100% MCP)/ha * | 544.54 ± 295.89 | 2828.3 ± 1269.43 |
| Activity area per night (100% MCP)/ha * | 101.46 ± 71.33 | 548.19 ± 85.57 |
| Core area (85% clusters)/ha * | 49.2 ± 25.6 | 149.7 ± 0.07 |
| Travel distance per night/km * | 4.23 ± 2.8 | 12.72 ± 3.7 |
| Max.nightly distance to foraging areas/km * | 2.01 ± 0.78 | 6.16 ± 2.12 |
| Number of foraging areas | 7.2 ± 1.3 | 9.54 ± 1.52 |
| Number of roosts | 2.4 ± 1.14 | 1 |
| Emergence/min after sunset | 30 ± 10 | 25 ± 15 |
| Foraging time/min * | 311 ± 135 | 347 ± 117 |
| Night roosting/min * | 40 ± 20 | 25 ± 10 |
| Plecotus auritus | Plecotus austriacus |
|---|---|
| Preservation of standing dead trees and trees with cavities or crevice roosts in older stands | Maintance or creation of hedgerows, rows of trees, copses and hedge strips as connecting lines of widely dispersed foraging habitats |
| Establishment of network of biotope tree candidates in younger stands (future trees/Z-trees) | Preservation of remaining unimproved grasslands and foster availability of new foraging habitats |
| Establishment of smaller, but well-connected, forest areas with deciduous and mixed stands near water bodies as hunting habitats (ideally primeval beech forests, with a mosaic of optimal stages (beech forest), decaying stages (patchy crown layer) and growing stages (multi-layered structure)) | Increase the availability of unmanaged field margins at arable or pasture field edges through management practices and promotion of structurally rich village edges (e.g., meadow orchards) |
| Optimization of forest edges (graded woodland fringes, flower-rich inner forest edges, shrub-rich outer forest edges) and forest meadows, loose multi-layered forests gaps and light shafts | Inclusion of and cooperation with land owners, land managers, farmers, and roost owners to increase awareness and identify roosts in buildings |
| Preservation of old buildings, especially in small dispersed settlements | |
| Avoidance of fertilizers and agrochemicals on grassland sites by farmers but also in private gardens in rural settlements | |
| Leaving standing and lying dead wood in any relevant bat habitat to increases insect diversity | |
| Reconstruction of wetland sites or creation of new wetland biotopes to establish productive foraging habitats. | |
| Consideration of differential arrival at and departure from maternity colonies with regard to renovation projects of buildings (e.g., renovation of churches) in the scheduling of construction measures (start and end of renovation measures). | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starik, N.; Göttert, T.; Zeller, U. Spatial Behavior and Habitat Use of Two Sympatric Bat Species. Animals 2021, 11, 3460. https://doi.org/10.3390/ani11123460
Starik N, Göttert T, Zeller U. Spatial Behavior and Habitat Use of Two Sympatric Bat Species. Animals. 2021; 11(12):3460. https://doi.org/10.3390/ani11123460
Chicago/Turabian StyleStarik, Nicole, Thomas Göttert, and Ulrich Zeller. 2021. "Spatial Behavior and Habitat Use of Two Sympatric Bat Species" Animals 11, no. 12: 3460. https://doi.org/10.3390/ani11123460
APA StyleStarik, N., Göttert, T., & Zeller, U. (2021). Spatial Behavior and Habitat Use of Two Sympatric Bat Species. Animals, 11(12), 3460. https://doi.org/10.3390/ani11123460