
Offshore Occurrence of a Migratory Bat, Pipistrellus nathusii, Depends on Seasonality and Weather Conditions


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
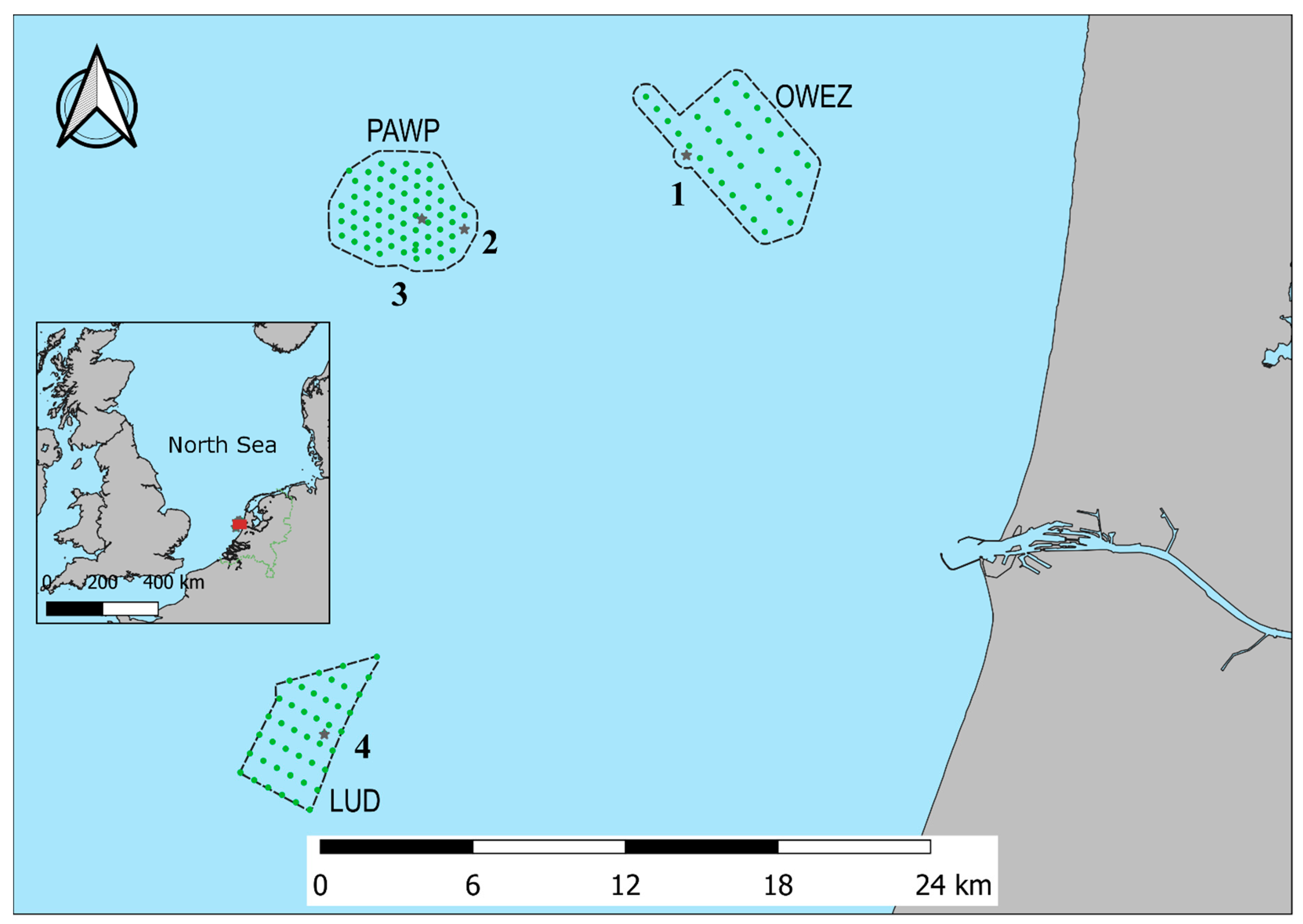
2.2. Equipment
2.3. Data Preparation
2.4. Statistical Analysis
3. Results
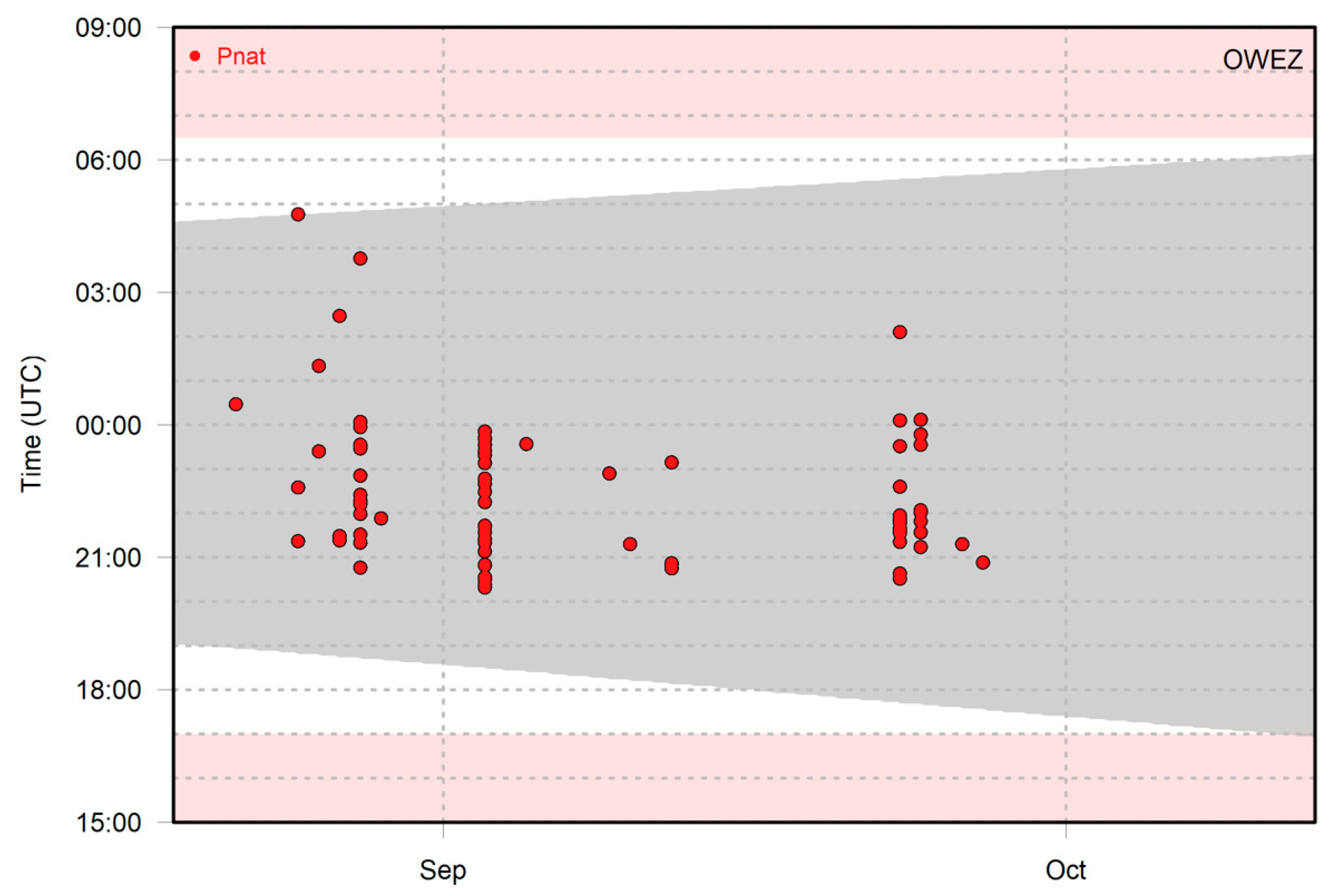
3.1. Monitoring Data
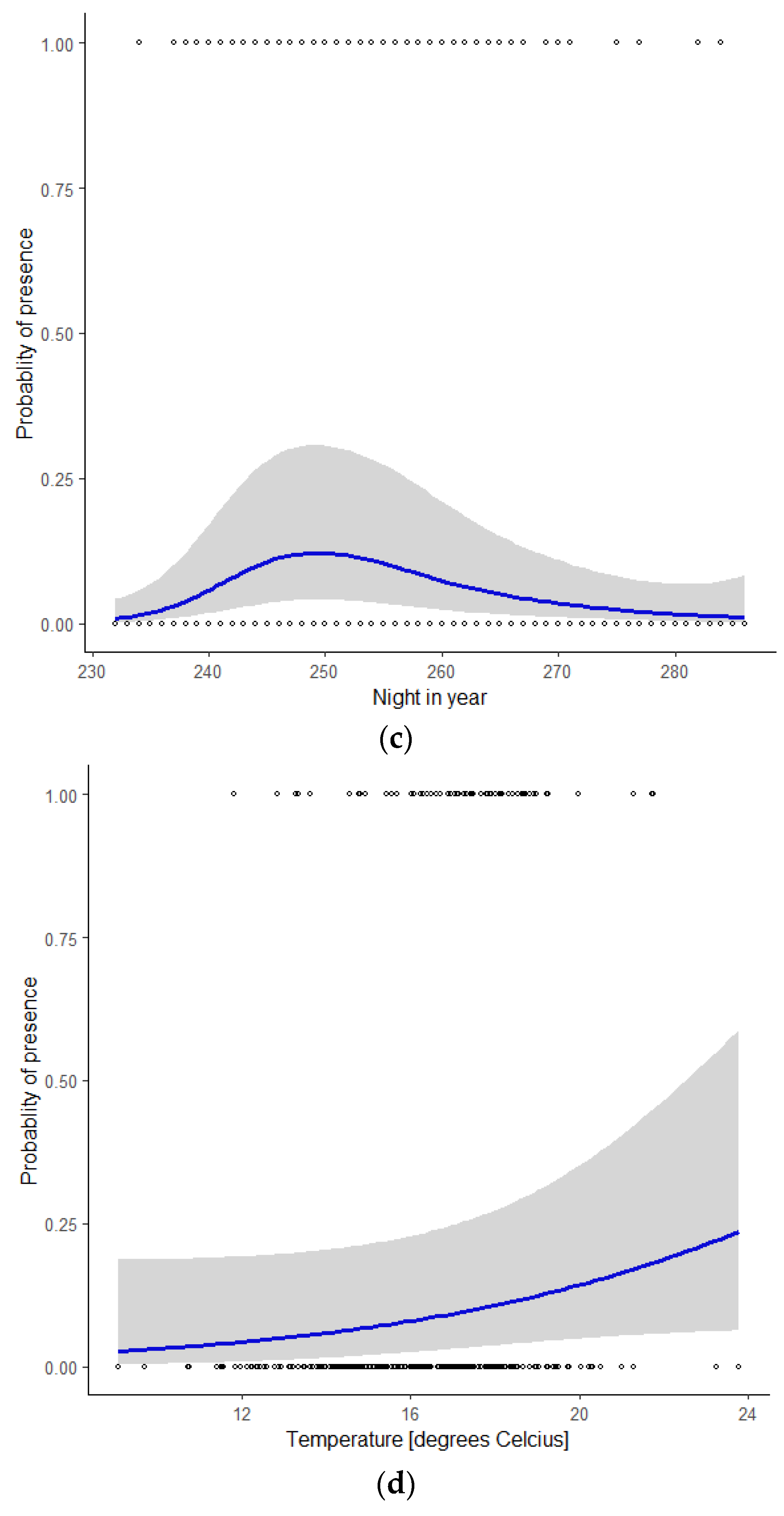
3.2. Graphical Representation of the Model
4. Discussion
5. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leung, D.Y.C.; Yang, Y. Wind energy development and its environmental impact: A review. Renew. Sustain. Energy Rev. 2012, 16, 1031–1039. [Google Scholar] [CrossRef]
- Lüdeke, J. Exploitation of Offshore Wind Energy. In Handbook on Marine Environment Protection; Salomon, M., Markus, T., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Bach, L.; Rahmel, U. Summary of wind turbine impacts on bats—Assessment of a conflict. Bremer Beiträge Nat. Nat. 2004, 7, 245–252. [Google Scholar]
- Kunz, T.H.; Arnett, E.B.; Erickson, W.P.; Johnson, G.D.; Larkin, R.P.; Strickland, M.D.; Thresher, R.W.; Tuttle, M.D. Ecological impacts of wind energy development on bats: Questions, hypotheses, and research needs. Front. Ecol. Environ. 2007, 5, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Arnett, E.B.; Brown, W.K.; Erickson, W.P.; Fiedler, J.K.; Hamilton, B.L.; Henry, T.H.; Jain, A.; Johnson, G.D.; Kerns, J.; Koford, R.R.; et al. Patterns of Bat Fatalities at Wind Energy Facilities in North America. J. Wildl. Manag. 2008, 72, 61–78. [Google Scholar] [CrossRef]
- Baerwald, E.F.; Edworthy, J.; Holder, M.; Barclay, R.M.R. A Large-Scale Mitigation Experiment to Reduce Bat Fatalities at Wind Energy Facilities. J. Wildl. Manag. 2009, 73, 1077–1081. [Google Scholar] [CrossRef]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Bat mortality at wind turbines in northwestern Europe. Acta Chiropterologica 2010, 12, 261–274. [Google Scholar] [CrossRef]
- Cryan, P.M.; Gorresen, P.M.; Hein, C.D.; Schirmacher, M.R.; Diehl, R.H.; Huso, M.M.; Hayman, D.T.; Fricker, P.D.; Bonaccorso, F.J.; Johnson, D.H.; et al. Behavior of bats at wind turbines. Proc. Natl. Acad. Sci. USA 2014, 111, 15126–15131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollins, K.M.; Meyerholz, D.K.; Johnson, G.D.; Capparella, A.P.; Loew, S.S. A forensic investigation into the etiology of bat mortality at a wind farm: Barotrauma or traumatic injury? Vet. Pathol. 2012, 49, 362–371. [Google Scholar] [CrossRef]
- Lawson, M.; Jenne, D.; Thresher, R.; Houck, D.; Wimsatt, J.; Straw, B. An investigation into the potential for wind turbines to cause barotrauma in bats. PLoS ONE 2020, 15, e0242485. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Behr, M.J.; Gendler, A.; Drake, D.; Dieterle, B.D.; Rudd, R.J.; Walrath, N.L. Investigating the causes of death for wind turbine-associated bat fatalities. J. Mammal. 2011, 92, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.C.; Sörgel, K.; Šuba, J.; Pētersons, G.; Keišs, O. The insectivorous bat Pipistrellus nathusii uses a mixed-fuel strategy to power autumn migration. Proc. R. Soc. Lond. 2012, 279B, 3772–3778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehnert, L.S.; Kramer-Schadt, S.; Schönborn, S.; Lindecke, O.; Niermann, I.; Voigt, C.C. Windfarm facilities in Germany kill Noctule bats from near and far. PLoS ONE 2014, 9, e103106. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.C.; Lehnert, L.S.; Petersons, G.; Adorf, F.; Bach, L. Wildlife and renewable energy: German politics cross migratory bats. Eur. J. Wildl. Res. 2015, 61, 213–219. [Google Scholar] [CrossRef]
- Hayes, M.A. Bats killed in large numbers at United States wind energy facilities. Bioscience 2013, 63, 975–979. [Google Scholar]
- Peste, F.; Paula, A.; da Silva, L.P.; Bernardino, J.; Pereira, P.; Mascarenhas, M.; Costa, H.; Vieira, J.; Bastos, C.; Fonseca, C.; et al. How to mitigate impacts of wind farms on bats? A review of potential conservation measures in the European context. Environ. Impact Assess. Rev. 2015, 51, 10–22. [Google Scholar] [CrossRef]
- Arnett, E.B.; Huso, M.M.; Schirmacher, M.R.; Hayes, J.P. Altering turbine speed reduces bat mortality at wind-energy facilities. Front. Ecol. Environ. 2011, 9, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Korner-Nievergelt, F.; Brinkmann, R.; Niermann, I.; Behr, O. Estimating bat and bird mortality occurring at wind energy turbines from covariates and carcass searches using mixture models. PLoS ONE 2013, 8, e67997. [Google Scholar] [CrossRef] [Green Version]
- Adams, E.M.; Gulka, J.; Williams, K.A. A review of the effectiveness of operational curtailment for reducing bat fatalities at terrestrial wind farms in North America. PLoS ONE 2021, 16, e0256382. [Google Scholar] [CrossRef]
- Whitby, M.; Schirmacher, M.; Frick, W. The State of the Science on Operational Minimization to Reduce Bat Fatality at Wind Energy Facilities; A report submitted to the National Renewable Energy Laboratory; Bat Conservation International: Austin, TX, USA, 2021. [Google Scholar]
- Arnett, E.B.; Hein, C.D.; Schirmacher, M.R.; Huso, M.M.P.; Szewczak, J.M. Evaluating the Effectiveness of an Ultrasonic Acoustic Deterrent for Reducing Bat Fatalities at Wind Turbines. PLoS ONE 2013, 8, e65794. [Google Scholar] [CrossRef]
- Gilmour, L.R.V.; Holderied, M.W.; Pickering, S.P.C.; Jones, G. Comparing acoustic and radar deterrence methods as mitigation measures to reduce human-bat impacts and conservation conflicts. PLoS ONE 2020, 15, e0228668. [Google Scholar] [CrossRef] [Green Version]
- Skiba, R. Die Fledermäuse im Bereich der Deutschen Nordsee unter Berücksichtigung der Gefährdungen durch Windenergieanlagen (WEA). Nyctalus (Neue Folge) 2007, 12, 199–220. [Google Scholar]
- Walter, G.; Matthes, H.; Joost, M. Fledermauszug über Nord- und Ostsee—Ergebnisse aus OffshoreUntersuchungen und deren Einordnung in das bisher bekannte Bild zum Zuggeschehen. Nyctalus 2007, 12, 221–223. [Google Scholar]
- Boshamer, J.P.C.; Bekker, J.P. Nathusius’ pipistrelles (Pipistrellus nathusii) and other species of bats on offshore platforms in the Dutch sector of the North Sea. Lutra 2008, 51, 17–36. [Google Scholar]
- Lagerveld, S.; Jonge Poerink, B.; Haselager, R.; Verdaat, H. Bats in Dutch offshore wind farms in autumn 2012. Lutra 2014, 57, 61–69. [Google Scholar]
- Hüppop, O.; Hill, R. Migration phenology and behaviour of bats at a research platform in the south-eastern North Sea. Lutra 2016, 59, 5–22. [Google Scholar]
- Hüppop, O.; Michalik, B.; Bach, L.; Hill, R.; Pelletier, S. Migratory birds and bats. In Wildlife and Windfarms, Conflicts and Solutions; Perrow, M.R., Ed.; Pelagic Publishing: Essex, UK, 2019; pp. 142–173. [Google Scholar]
- Ahlén, I.; Baagøe, H.J.; Bach, L. Behavior of Scandinavian Bats during Migration and Foraging at Sea. J. Mammal. 2009, 90, 1318–1323. [Google Scholar] [CrossRef] [Green Version]
- Lack, D. Migration across the southern North Sea studied by radar part 5. Movements in August, winter and spring, and conclusion. Ibis 1963, 105, 461–492. [Google Scholar] [CrossRef]
- Lack, D. Migration across the southern North Sea studied by radar part 5. Ibis 1963, 105, 1–54. [Google Scholar] [CrossRef]
- Barataud, M. Acoustic Ecology of European Bats: Species Identification, Study of Their Habitats and Foraging Behavior; University of Chicago Press: Chicago, IL, USA, 2016; ISBN 987-2-85653-771-8. [Google Scholar]
- Lazaridis, E. Lunar: Lunar Phase & Distance, Seasons and Other Environmental Factors (Version 0.1-04). 2014. Available online: http://statistics.lazaridis.eu (accessed on 30 June 2017).
- Zuur, A.F.; Ieno, E.N. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 2016, 7, 636–645. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 5 July 2021).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.A. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Mitchell-Jones, A.J.; Amori, G.; Bogdanowicz, W.; Krystufek, B.; Reijnders, P.J.H.; Spitzenberger, F.; Stubbe, M.; Thissen, J.B.M.; Vohralik, V.; Zima, J. The Atlas of European Mammals; Poyser: London, UK, 1999. [Google Scholar]
- Russ, J.M.; Hutson, A.M.; Montgomery, W.I.; Racey, P.A.; Speakman, J.R. The status of Nathusius’ pipistrelle (Pipistrellus nathusii Keyserling & Blasius, 1839) in the British Isles. J. Zool. 2001, 254, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Hutterer, R.; Ivanova, T.; Meyer-Cords, C.; Rodrigues, L. Bat Migrations in Europe: A Review of Banding Data and Literature; Naturschutz und Biologische Vielfalt: Bonn, Germany, 2005; Volume 28, pp. 1–176. [Google Scholar]
- Pētersons, G. Seasonal migrations of north-eastern populations of Nathusius’ bat Pipistrellus nathusii (Chiroptera). Myotis 2004, 41, 29–56. [Google Scholar]
- Alcalde, J.T.; Jiménez, M.; Brila, I.; Vintulis, V.; Voigt, C.C.; Pētersons, G. Transcontinental 2200 km migration of a Nathusius’ pipistrelle (Pipistrellus nathusii) across Europe. Mammalia 2021, 85, 161–163. [Google Scholar] [CrossRef]
- Sachanowicz, K.; Ciechanowski, M.; Tryjanowski, P.; Kosicki, J.Z. Wintering range of Pipistrellus nathusii (Chiroptera) in Central Europe: Has the species extended to the north-east using urban heat islands? Mammalia 2019, 83, 260–271. [Google Scholar] [CrossRef]
- Jahelkova, H.; Horacek, L. Mating system of a migratory bat, Nathusius’ pipistrelle (Pipistrellus nathusii): Different male strategies. Acta Chiropterologica 2011, 16, 139–147. [Google Scholar] [CrossRef]
- Petersen, A.; Jensen, J.K.; Jenkins, P.; Bloch, D.; Ingimarsson, F. A review of the occurrence of bats (Chiroptera) on islands in the North East Atlantic and on North Sea installations. Acta Cropterologica 2014, 16, 169–195. [Google Scholar] [CrossRef] [Green Version]
- Fleming, T.H.; Eby, P. Ecology of bat migration. In Bat Ecology; Kunz, T.H., Fenton, M.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 156–208. [Google Scholar]
- Clerc, J.; Rogers, E.J.; McGuire, L.P. Testing Predictions of Optimal Migration Theory in Migratory Bats. Front. Ecol. Evol. 2021, 9, 686379. [Google Scholar] [CrossRef]
- Hedenström, A. Optimal migration strategies in bats. J. Mammal. 2009, 90, 1298–1309. [Google Scholar] [CrossRef] [Green Version]
- Alerstam, T.; Lindström, A. Optimal bird migration: The relative importance of time, energy and safety. In Bird Migration: Physiology and Ecophysiology; Gwinner, E., Ed.; Springer: Berlin, Germany, 1990; pp. 331–351. [Google Scholar]
- Alerstam, T. Bird Migration; First paperback edition; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Berthold, P. Vogelzug: Eine Einfühfrung und Kurze Aktuelle Gesamtübersicht; Wissenschaftliche Buchgesellschaft: Darmstadt, Germany, 1990. [Google Scholar]
- Berthold, P.; Querner, U. Genetic basis of migratory behavior in European warblers. Science 1981, 212, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Gwinner, E. Annual rhythms of gonadal size, migratory disposition and molt in garden warblers Sylvia borin exposed in winter to an equatorial or a southern hemisphere photoperiod. Ornis Scand. 1987, 18, 251–256. [Google Scholar] [CrossRef]
- McGuire, L.P.; Guglielmo, C.G. What can birds tell us about the migration physiology of bats? J. Mammal. 2009, 90, 1290–1297. [Google Scholar] [CrossRef] [Green Version]
- Dechmann, D.K.N.; Wikelski, M.; Ellis-Soto, D.; Safi, K.; O’Mara, M.T. Determinants of spring migration departure decision in a bat. Biol. Lett. 2017, 13, 20170395. [Google Scholar] [CrossRef] [PubMed]
- Šuba, J.; Pētersons, G.; Rydell, J. Fly-and-forage strategy in the bat Pipistrellus nathusii during autumn migration. Acta Chiropterologica 2012, 14, 379–385. [Google Scholar]
- Voigt, C.C.; Sörgel, K.; Dechmann, D.K.N. Refuelling while flying: Foraging bats combust food rapidly and directly to fuel flight. Ecology 2010, 91, 2908–2917. [Google Scholar] [CrossRef]
- Cryan, P.M.; Brown, A.C. Migration of bats past a remote island offers clues toward the problem of bat fatalities at wind turbines. Biol. Conserv. 2007, 139, 1–11. [Google Scholar] [CrossRef]
- Johnson, J.B.; Gates, J.E.; Zegre, N.P. Monitoring seasonal bat activity on a coastal Barrier Island in Maryland. Environ. Monit. Assess. 2011, 173, 685–699. [Google Scholar] [CrossRef]
- Sjollema, A.L.; Gates, J.E.; Hilderbrand, R.H.; Sherwell, J. Offshore Activity of Bats along the Mid-Atlantic Coast. Northeast. Nat. 2014, 21, 154–163. [Google Scholar] [CrossRef]
- Brabant, R.; Laurent, Y.; Jonge Poerink, B.; Degraer, S. Activity and Behaviour of Nathusius’ Pipistrelle Pipistrellus nathusii at Low and High Altitude in a North Sea Offshore Wind Farm. Acta Chiropterologica 2019, 21, 341–348. [Google Scholar] [CrossRef]
- Hatch, S.K.; Connelly, E.E.; Divoll, T.J.; Stenhouse, I.J.; Williams, K.A. Offshore Observations of Eastern Red Bats (Lasiurus borealis) in the Mid-Atlantic United States Using Multiple Survey Methods. PLoS ONE 2013, 8, e83803. [Google Scholar] [CrossRef] [Green Version]
- National Nathusius’ Pipistrelle Project. Available online: https://www.bats.org.uk/our-work/national-bat-monitoring-programme/surveys/national-nathusius-pipistrelle-survey (accessed on 30 September 2021).
- Brinkmann, R.; Behr, O.; Niermann, I.; Reich, M. Entwicklung von Methoden zur Untersuchung und Reduktion des Kollisionsrisikos von Fledermäusen an Onshore-Windenergieanlagen; Volume 4 Umwelt und Raum; Cuvillier Verlag: Göttingen, Germany, 2011. [Google Scholar]
- Drake, V.A.; Gatehouse, A.G. Insect Migration: Tracking Resources through Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Chapman, J.W.; Reynolds, D.R.; Smith, A.D.; Smith, E.T.; Woiwod, I.P. An aerial netting study of insects migrating at high altitude over England. Bull. Entomol. Res. 2004, 94, 123–136. [Google Scholar] [CrossRef]
- Drake, V.; Reynolds, D. Radar Entomology: Observing Insect Flight and Migration; Cabi: Wallingford, UK, 2012. [Google Scholar]
- Šuba, J. Migrating Nathusius’s pipistrelles Pipistrellus nathusii (Chiroptera: Vespertilionidae) optimise flight speed and maintain acoustic contact with the ground. Environ. Exp. Biol. 2014, 12, 7–14. [Google Scholar]
- Walters, C.L.; Collen, A.; Lucas, T.; Mroz, K.; Sayer, C.A.; Jones, K.E. Challenges of Using Bioacoustics to Globally Monitor Bats. In Bat Evolution, Ecology, and Conservation; Adams, R., Pedersen, S., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar] [CrossRef]
- Voigt, C.C.; Russo, D.; Runkel, V.; Goerlitz, H.R. Limitations of acoustic monitoring at wind turbines to evaluate fatality risk of bats. Mam. Rev. 2021, 51, 559–570. [Google Scholar] [CrossRef]
- Marques, T.A.; Thomas, L.; Martin, S.W.; Mellinger, D.K.; Ward, J.A.; Moretti, D.J.; Harris, D.; Tyack, P.L. Estimating animal population density using passive acoustics. Biol. Rev. 2013, 88, 287–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, C.; Moss, C.F. When echolocating bats do not echolocate. Commun. Integr. Biol. 2008, 1, 161–162. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, A.J.; Weller, T.J. Inconspicuous echolocation in hoary bats (Lasiurus cinereus). Proc. R. Soc. B Biol. Sci. 2018, 285, 20180441. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, A.J.; Weller, T.J.; Hopkins, A.; Yovel, Y. Silence and reduced echolocation during flight are associated with social behaviors in male hoary bats (Lasiurus cinereus). Sci. Rep. 2021, 11, 18637. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E.W.; Hennessey, J.P., Jr. On the Use of Power Laws for Estimates of Wind Power Potential. J. Appl. Meteorol. Climatol. 1978, 17, 390–394. [Google Scholar] [CrossRef]
- Chapman, J.W.; Reynolds, D.R.; Smith, A.D. Vertical-Looking Radar: A New Tool for Monitoring High-Altitude Insect Migration. BioScience 2003, 53, 503–511. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monitoring Location | Geographical Location | Distance to Shore [Km] | Heigh Above Sea Level [M] | Microphone Direction | |
|---|---|---|---|---|---|
| 1 | OWEZ mast | 52°61′ N, 4°39′ E | 15 | 15 | East |
| 2 | PAWP WT22 | 52°58′ N, 4°27′ E | 23 | 15 | East |
| 3 | PAWP OHVS | 52°59′ N, 4°24′ E | 25 | 15 | East |
| 4 | LUD OHVS | 52°40′ N, 4°17′ E | 25 | 15 | East |
| OWEZ Mast | PAWP WT22 | PAWP OHVS | LUD OHVS | Total | |
|---|---|---|---|---|---|
| 2012 | 29 August–20 October | 4–23 September | - | - | |
| N | 45 | 19 | 0 | 0 | 64 |
| 2013 | 1 July–15 October | 5 August–2 October | - | - | |
| N | 55 | 43 | 0 | 0 | 98 |
| 2014 | 1 July–14 October | - | 1 July–15 October | - | |
| N | 55 | 0 | 55 | 0 | 110 |
| 2015 | - | - | 1 July–20 October | 1 July–9 October | |
| N | 0 | 0 | 55 | 43 | 98 |
| 2016 | - | - | 1 July–17 October | 1 July–24 October | |
| N | 0 | 0 | 55 | 55 | 110 |
| Total | 155 | 62 | 165 | 98 | 480 |
| Optimal Model: | ||||
|---|---|---|---|---|
| Parametric Coefficients: | Estimate | Std. Error | Z Value | Pr (>|z|) 1 |
| Intercept | −1.59188 | 1.69250 | −0.941 | 0.3469 |
| Windspeed | −0.54433 | 0.08546 | −6.370 | 1.9 × 10 −10 *** |
| Temperature | 0.16386 | 0.09691 | 1.691 | 0.0908 |
| Significance of smooth terms | Edf | Ref.df | Chi.sq | p-value 1 |
| s (night in year) | 3.533 | 3.533 | 18.39 | 0.000487 *** |
| s (wind direction) | 4.130 | 8.000 | 36.05 | <2 × 10−16 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagerveld, S.; Jonge Poerink, B.; Geelhoed, S.C.V. Offshore Occurrence of a Migratory Bat, Pipistrellus nathusii, Depends on Seasonality and Weather Conditions. Animals 2021, 11, 3442. https://doi.org/10.3390/ani11123442
Lagerveld S, Jonge Poerink B, Geelhoed SCV. Offshore Occurrence of a Migratory Bat, Pipistrellus nathusii, Depends on Seasonality and Weather Conditions. Animals. 2021; 11(12):3442. https://doi.org/10.3390/ani11123442
Chicago/Turabian StyleLagerveld, Sander, Bob Jonge Poerink, and Steve C. V. Geelhoed. 2021. "Offshore Occurrence of a Migratory Bat, Pipistrellus nathusii, Depends on Seasonality and Weather Conditions" Animals 11, no. 12: 3442. https://doi.org/10.3390/ani11123442
APA StyleLagerveld, S., Jonge Poerink, B., & Geelhoed, S. C. V. (2021). Offshore Occurrence of a Migratory Bat, Pipistrellus nathusii, Depends on Seasonality and Weather Conditions. Animals, 11(12), 3442. https://doi.org/10.3390/ani11123442