
Activity Pattern and Correlation between Bat and Insect Abundance at Wind Turbines in South Sweden

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
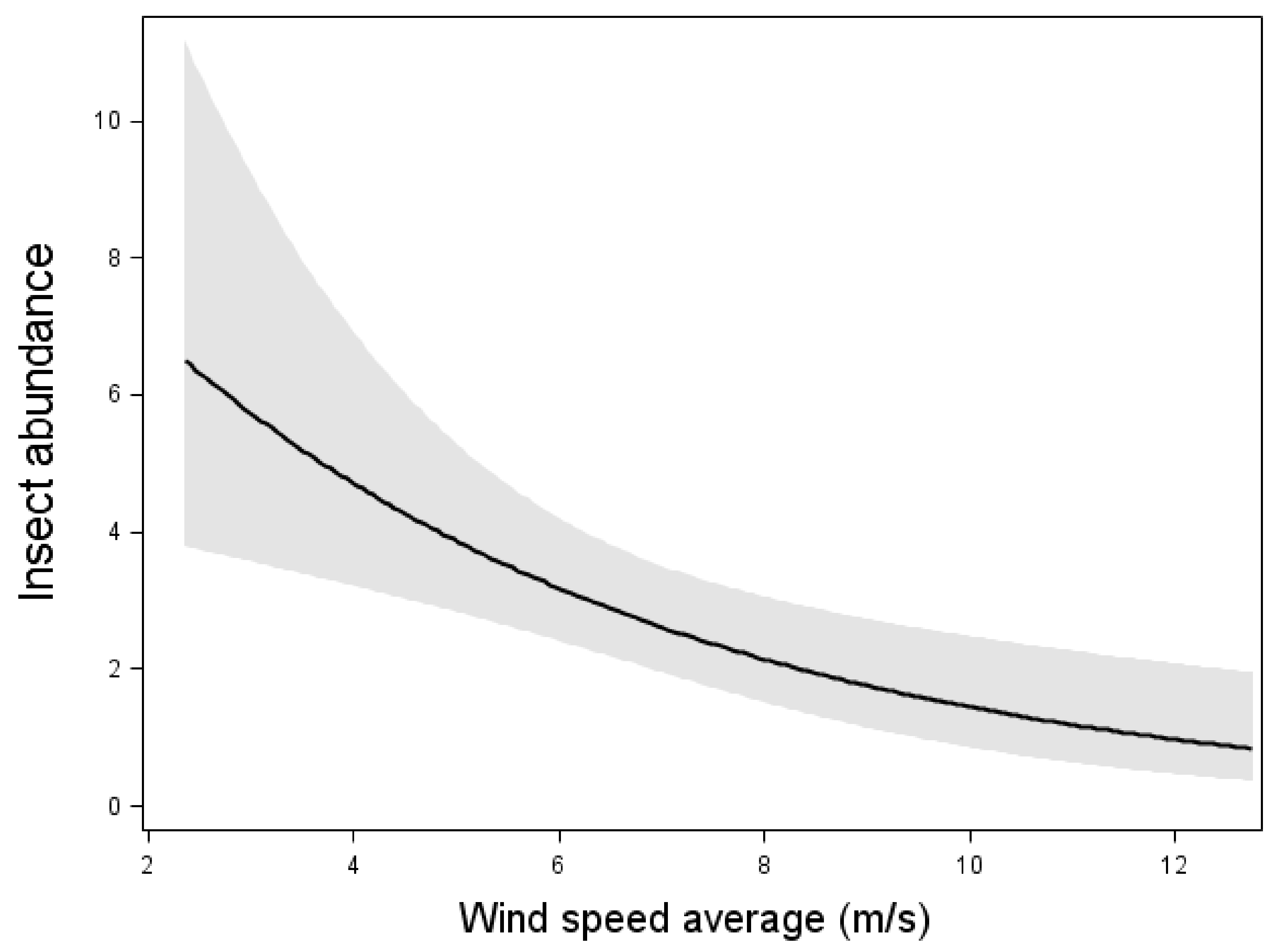
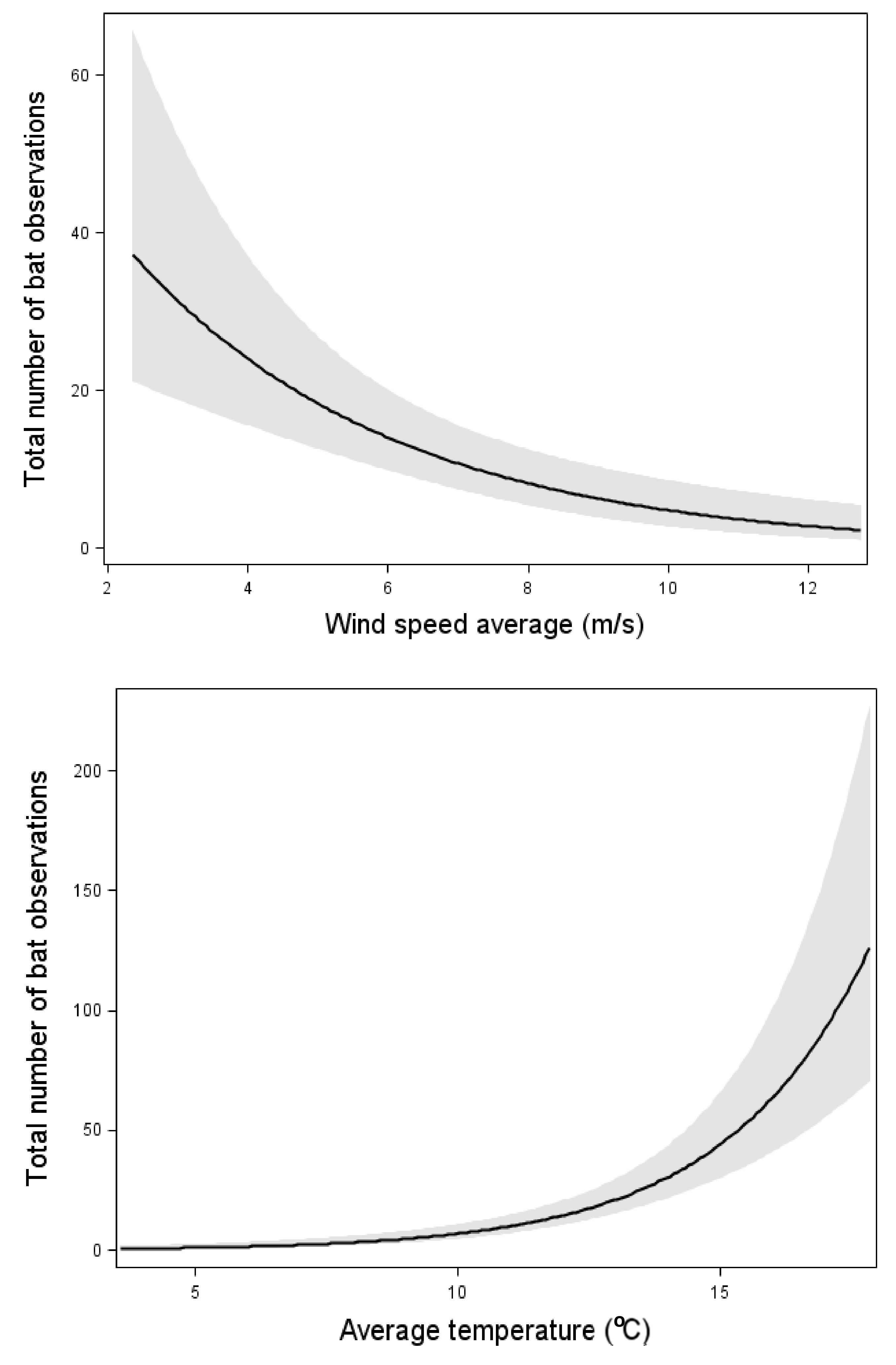
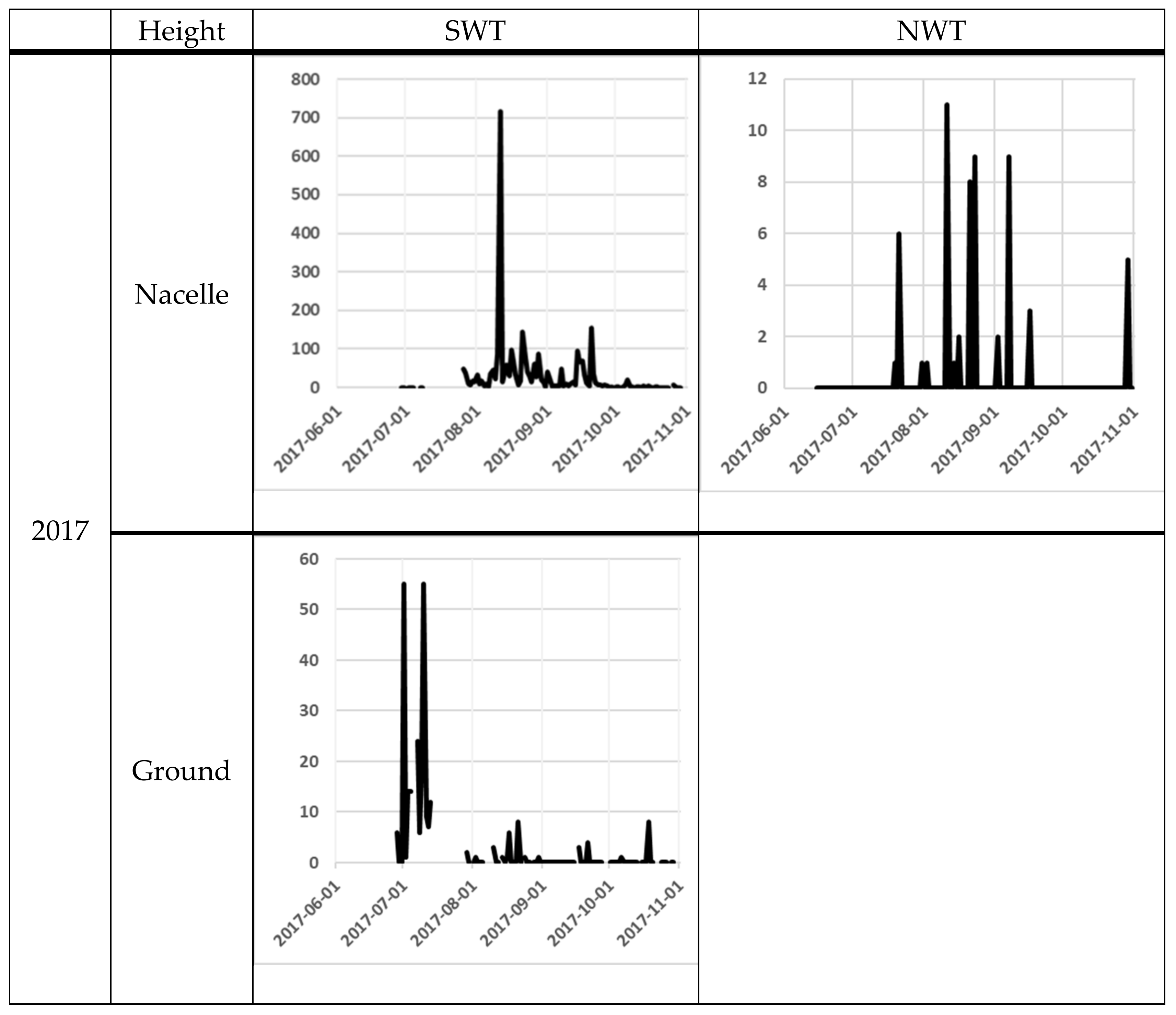
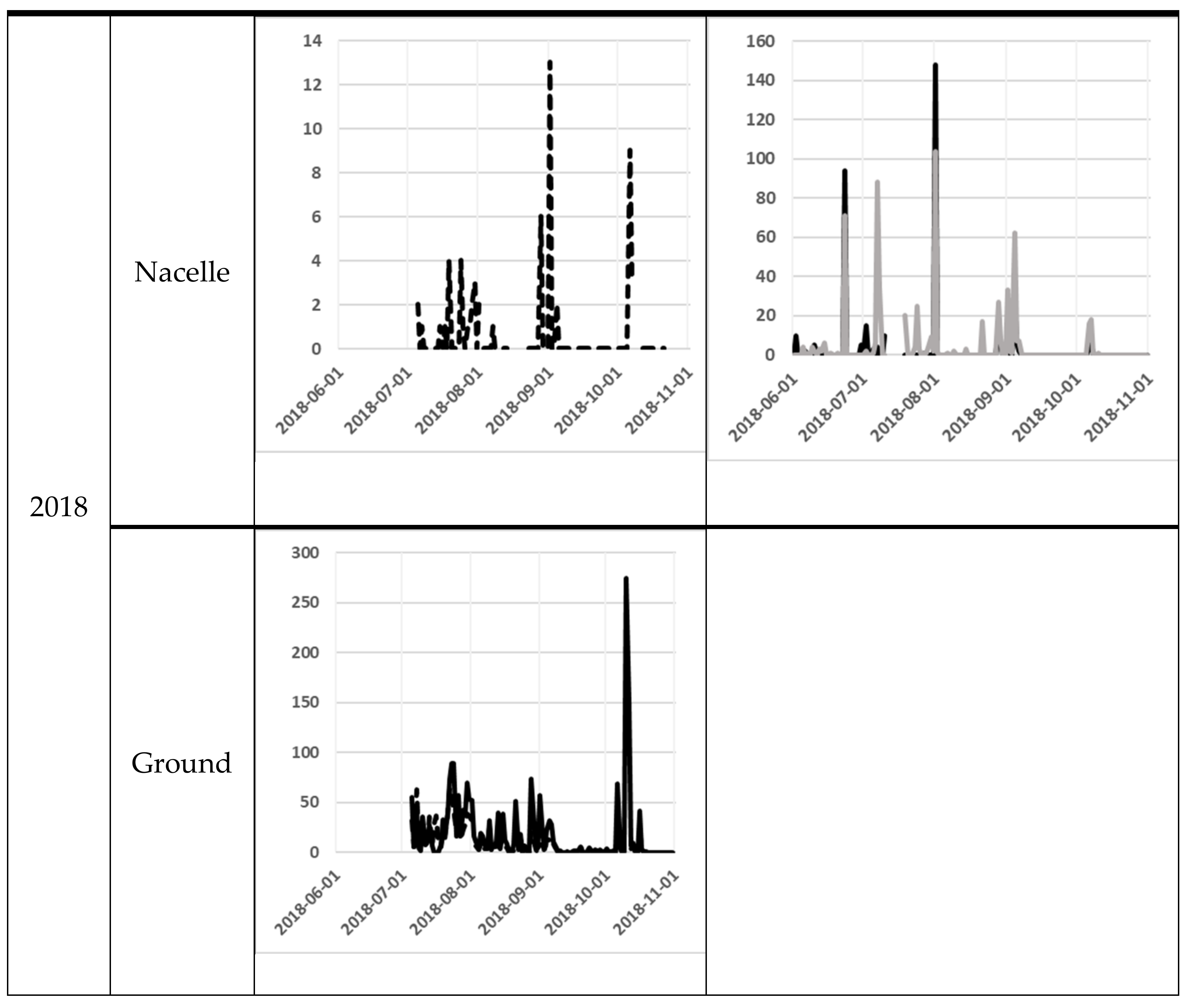
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Avisoft SWT | Avisoft NWT | D500X SWT |
|---|---|---|---|
| Pre-trigger | 0.5s | 0.5s | OFF |
| Hold | 2 | 4.5s | |
| Level | 0,35 | 0.35 | |
| Range | 14–70 kHz | 14–70 kHz | |
| Entropy | No | Y = 35% | |
| Reject wind/rain | No | No | |
| Whistle tracking | No | No | |
| Max file size | 0.12 min = 7.2 s | 0.09 min = 5.4 s | 5 s |
| Interval between recording | 5 s | ||
| Sample frequency | 500 | ||
| Trigger sens. | High | ||
| Input gain | 60 | ||
| Trig level | 30 | ||
| Bat call filter | Yes | Yes | |
| Bat call filter settings | Standard | Standard | |
| HP Filter | Yes | ||
| Time programme | |||
| Type | absolute timer | absolute timer | absolute timer |
| From | 19:00 | 19:00 | 19:00 |
| To | 06:00 | 06:00 | 06:00 |
References
- Hossain, J. Wind Energy, on the Shape of Near 100% RE Grid; A WWEA Technical Committee Report on Grid Integration; World Wind Energy Association: Bonn, Germany, 2015; Available online: https://www.scribd.com/document/316511808/WWEA-Report-Wind-Energy-2050 (accessed on 15 November 2021).
- Dai, K.; Bergot, A.; Liang, C.; Xiang, W.-N.; Huang, Z. Environmental issues associated with wind energy—A review. Renew. Energy 2015, 75, 911–921. [Google Scholar] [CrossRef]
- Arnett, E.B.; Baerwald, E.F. Impacts of wind energy development on bats: Implications for conservation. In Bat Evolution, Ecology and Conservation; Adams, R.A., Peterson, S.C., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Barré, K.; Le Viol, I.; Bas, Y.; Julliard, R.; Kerbiriou, C. Estimating habitat loss due to wind turbine avoidance by bats: Implications for European siting guidance. Biol. Conserv. 2018, 226, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Millon, L.; Julien, J.-F.; Julliard, R.; Kerbiriou, C. Bat activity in intensively farmed landscapes with wind turbines and offset measures. Ecol. Eng. 2015, 75, 250–257. [Google Scholar] [CrossRef]
- Millon, L.; Colin, C.; Brescia, F.; Kerbiriou, C. Wind turbines impact bat activity, leading to high losses of habitat use in a biodiversity hotspot. Ecol. Eng. 2018, 112, 51–54. [Google Scholar] [CrossRef]
- Kunz, T.H.; Arnett, E.B.; Erickson, W.P.; Hoar, A.R.; Johnson, G.D.; Larkin, R.P.; Strickland, M.D.; Thresher, R.W.; Tuttle, M.D. Ecological impacts of wind energy development on bats: Questions, research needs, and hypotheses. Front. Ecol. Environ. 2007, 5, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Cryan, P.M.; Barclay, R.M.R. Causes of Bat Fatalities at Wind Turbines: Hypotheses and Predictions. J. Mammal. 2009, 90, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Jansson, S.; Malmqvist, E.; Brydegaard, M.; Åkesson, S.; Rydell, J. A Scheimpflug lidar used to observe insect swarming at a wind turbine. Ecol. Indic. 2020, 117, 106578. [Google Scholar] [CrossRef]
- Long, C.V.; Flint, J.A.; Lepper, P.A. Insect attraction to wind turbines: Does color play a role? Eur. J. Wildl. Res. 2011, 57, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Downes, J.A. Swarming and mating flight of Diptera. Annu. Rev. Entomol. 1969, 14, 271. [Google Scholar] [CrossRef]
- Geerts, B.; Miao, Q. A simple numerical model of the flight behavior of small insects in the atmospheric convective boundary layer. Environ. Entomol. 2005, 34, 353–360. [Google Scholar] [CrossRef]
- Svensson, B.W.; Solbreck, B. Stövsländor på väg i lufthavet. Fångster i sugfällor på en TV-mast i Värmland. Entomologisk Tidskrift 2008, 129, 29–39. [Google Scholar]
- Reynolds, D.R.; Chapman, J.W.; Edwards, A.S.; Smith, A.D.; Wood, C.R.; Barlow, J.F.; Woiwod, I.P. Radar studies of the vertical distribution of insects migrating over southern Britain: The influence of temperature inversions on nocturnal layer concentrations. Bull. Entomol. Res. 2005, 95, 259–274. [Google Scholar] [CrossRef] [Green Version]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Mortality of bats at wind turbines links to nocturnal insect migration? Eur. J. Wildl. Res. 2010, 56, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Valdez, E.W.; Cryan, P.M. Insect Prey Eaten by Hoary Bats (Lasiurus cinereus) Prior to Fatal Collisions with Wind Turbines. West. North Am. Nat. 2013, 73, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Bennet, V.J.; Hale, A.M.; Williams, D.A. When the excrement hits the fan: Fecal surveys reveal species-specific bat activity at wind turbines. Mamm. Biol. 2017, 87, 125–129. [Google Scholar] [CrossRef]
- Foo, C.F.; Bennet, V.J.; Hale, A.M.; Korstian, J.M.; Schildt, A.J.; Williams, D.A. Increasing evidence that bats actively forage at wind turbines. PeerJ 2017, 5, e3985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimer, J.P.; Baerwald, E.F.; Barclay, R.M.R. Echolocation activity of migratory bats at a wind energy facility: Testing the feeding-attraction hypothesis to explain fatalities. J. Mammal. 2018, 99, 1472–1477. [Google Scholar] [CrossRef]
- Rydell, J.; Bogdanowicz, W.; Boonman, A.; Pettersson, S.; Sucheka, E.; Pomorski, J.J. Bats may eat diurnal flies that rest on wind turbines. Mamm. Biol. 2016, 81, 331–339. [Google Scholar] [CrossRef]
- Hurst, J.; Biedermann, M.; Dietz, C.; Dietz, M.; Karst, I.; Krannich, E.; Petermann, R.; Schorcht, W.; Brinkmann, R. Flädermäuse und windkraft im wald. Naturschutz und biologische vielfalt: Heft 153; Bundesamt für Naturschutz: Bonn, Germany, 2016. [Google Scholar]
- Rydell, J.; Ottvall, R.; Pettersson, S.; Green, M. Vindkraftens påverkan på fåglar och fladdermöss: Uppdaterad Syntesrapport 2017; Naturvårdaverket: Stockholm, Sweden, 2017. [Google Scholar]
- Behr, O.; Brinkmann, R.; Hochradel, K.; Mages, J.; Korner-Nievergelt, F.; Niermann, I.; Reich, M.; Simon, R.; Weber, N.; Nagy, M. Mitigating bat mortality with turbine-specific curtailment algorithms: A model based approach. In Wind Energy and Wildlife Interactions; Köppel, J., Ed.; Springer: Cham, Switzerland, 2017; pp. 135–160. [Google Scholar]
- Baerwald, E.F.; Barclay, R.M.R. Patterns of activity and fatality of migratory bats at a wind energy facility in Alberta, Canada. J. Wildl. Manag. 2011, 75, 1103–1114. [Google Scholar] [CrossRef]
- Roemer, C.; Disca, T.; Coulon, A.; Bas, Y. Bat flight height monitored from wind masts predicts mortality risk at wind farms. Biol. Conserv. 2017, 215, 116–122. [Google Scholar] [CrossRef]
- Baerwald, E.F.; Edworthy, J.; Holder, M.; Barclay, R.M.R. A large-scale mitigation experiment to reduce bat fatalities and wind energy facilities. J. Wildl. Manag. 2009, 73, 1077–1081. [Google Scholar] [CrossRef]
- Arnett, E.B.; Baerwald, E.F.; Mathews, F.; Rodrigues, L.; Rodrigues-Durán, A.; Rydell, J.; Villegas-Patraca, R.; Voigt, C.C. Impacts of wind energy development on bats: A global perspective. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Cham, Switzerland, 2013. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.M.; Arnett, E.B.; Stevens, R.D.; Wallace, M.C. Reducing bat fatalities at wind facilities while improving the economic efficiency of operational mitigation. J. Mammal. 2017, 98, 378–385. [Google Scholar] [CrossRef]
- Hayes, M.A.; Hooton, L.A.; Gilland, K.L.; Grandgent, C.; Smith, R.L.; Lindsay, S.R.; Collins, J.D.; Schumacher, S.M.; Rabie, P.A.; Gruver, J.C.; et al. A smart curtailment approach for reducing bat fatalities and curtailment time at wind energy facilities. Ecol. Appl. 2019, 29, e01881. [Google Scholar] [CrossRef]
- Smallwood, K.S.; Bell, D.A. Effects of wind turbine curtailment on bird and bat fatalities. J. Wildl. Manag. 2020, 84, 685–696. [Google Scholar] [CrossRef]
- Rodrigues, L.; Bach, L.; Dubourg-Savage, M.-J.; Karapandža, B.; Kovač, D.; Kervyn, T.; Dekker, J.; Kepel, A.; Bach, P.; Collins, J.; et al. Guidelines for Consideration of Bats in Wind Farm Projects—Revision 2014; Eurobats Publication Series No. 6. UNEP/EUROBATS Secretariat: Bonn, Germany, 2015. [Google Scholar]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods, 3rd ed.; Blackwell Science: Oxford, UK, 2000. [Google Scholar]
- Rydell, J.; Entwistle, A.; Racey, P.A. Timing of foraging flights of three species of bats in relation to insect activity and predation risk. Oikos 1996, 76, 243–252. [Google Scholar] [CrossRef]
- Park, K.J.; Cristinacce, A. Use of sewage treatment works as foraging sites by insectivorous bats. Anim. Conserv. 2009, 9, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, B.; Racey, P.A. The aversive effect of electromagnetic radiation on foraging bats-a possible means of discouraging bats from approaching wind turbines. PLoS ONE 2009, 4, e6246. [Google Scholar] [CrossRef] [Green Version]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Littell, R.; Stroup, W.; Freund, R. SAS for Linear Models, 4th ed.; SAS Institute Inc.: Cary, NC, USA, 2002; ISBN 9781599941424. [Google Scholar]
- Voigt, C.C.; Lehnert, L.S.; Petersons, G.; Adorf, F.; Bach, L. Wildlife and renewable energy: German politics cross migratory bats. Eur. J. Wildl. Res. 2015, 61, 213–219. [Google Scholar] [CrossRef]
- Ahlén, I.; Bach, L.; Baagøe, H.J.; Pettersson, J. Fladdermöss Och Havsbaserade Vindkraftverk Studerade I Södra Skandinavien—Rapport 5748; Naturvårdsverket: Stockholm, Sweden, 2007. [Google Scholar]
- Ahlén, I.; Baagøe, H.J.; Bach, L. Behavior of Scandinavian bats during migration and foraging at sea. J. Mammol. 2009, 90, 1318–1323. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.C.; Russo, D.; Runkel, V.; Goerlitz, H.R. Limitations of acoustic monitoring at wind turbines to evaluate fatality risk of bats. Mammal Rev. 2021, 51, 559–570. [Google Scholar] [CrossRef]
- Barataud, M. Acoustic Ecology of European Bats, Species Identification, Study of Their Habitats and Foraging Behaviour; Muséum National d’Histoire Naturelle: Paris, France, 2015. [Google Scholar]
- Budenz, T.; Gessner, B.; Luttmann, J.; Molitor, F.; Servatius, K.; Veith, M. Up and down: B. barbastellus explore lattice towers. Hystrix 2017, 28, 272–276. [Google Scholar]
- Solick, D.; Pham, D.; Nasman, K.; Bay, K. Bat Activity Rates do not Predict Bat Fatality Rates at Wind Energy Facilities. Acta Chiropterologica 2020, 22, 135–146. [Google Scholar] [CrossRef]
- Zimmerling, J.R.; Francis, C.M. Bat mortality due to wind turbines in Canada. J. Wildl. Manag. 2016, 80, 1360–1369. [Google Scholar] [CrossRef]
- Roemer, C.; Bas, Y.; Disca, T.; Coulon, A. Influence of landscape and time of year on bat-wind turbines collision risks. Landscape Ecol. 2019, 34, 2869–2881. [Google Scholar] [CrossRef]
- Cryan, P.M.; Gorresen, P.M.; Hein, C.D.; Schumacher, M.R.; Diehl, R.H.; Huso, M.M.; Hayman, D.T.S.; Fricker, P.D.; Bonaccorso, F.J.; Johnson, D.H.; et al. Behavior of bats at wind turbines. Proc. Natl. Acad. Sci. USA 2014, 111, 15126–15131. [Google Scholar] [CrossRef] [Green Version]
- Lintott, P.R.; Richardson, S.M.; Hosken, D.J.; Fensome, S.A.; Mathews, F. Ecological impact assessments fail to reduce risk of bat casualties at wind farms. Curr. Biol. 2016, 26, 1119–1136. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.M.; Lintott, P.R.; Hosken, D.J.; Economou, T.; Mathews, F. Peaks in bat activity at turbines and the implications for mitigating the impact of wind energy developments on bats. Sci. Rep. 2021, 11, 3636. [Google Scholar] [CrossRef]
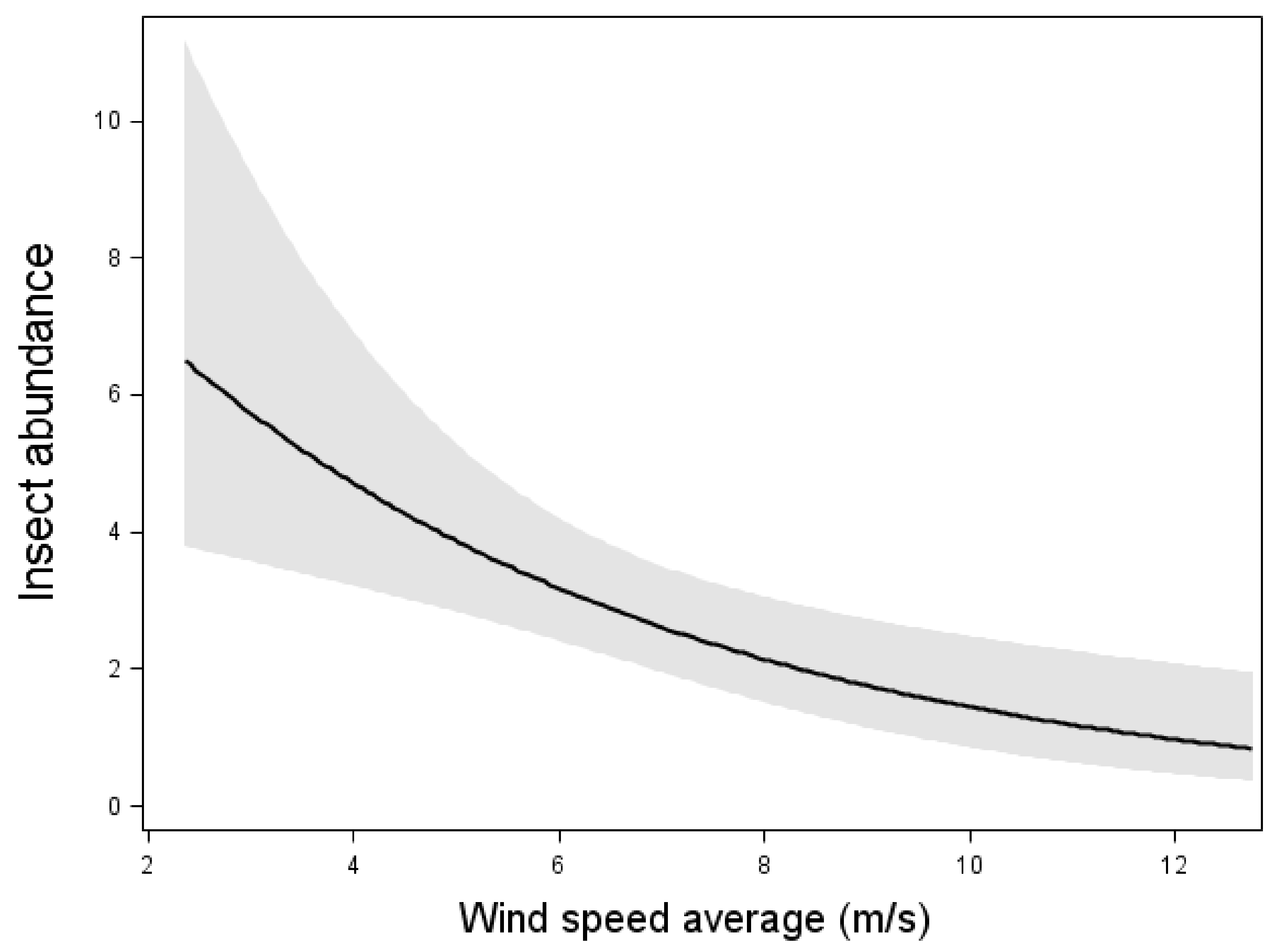
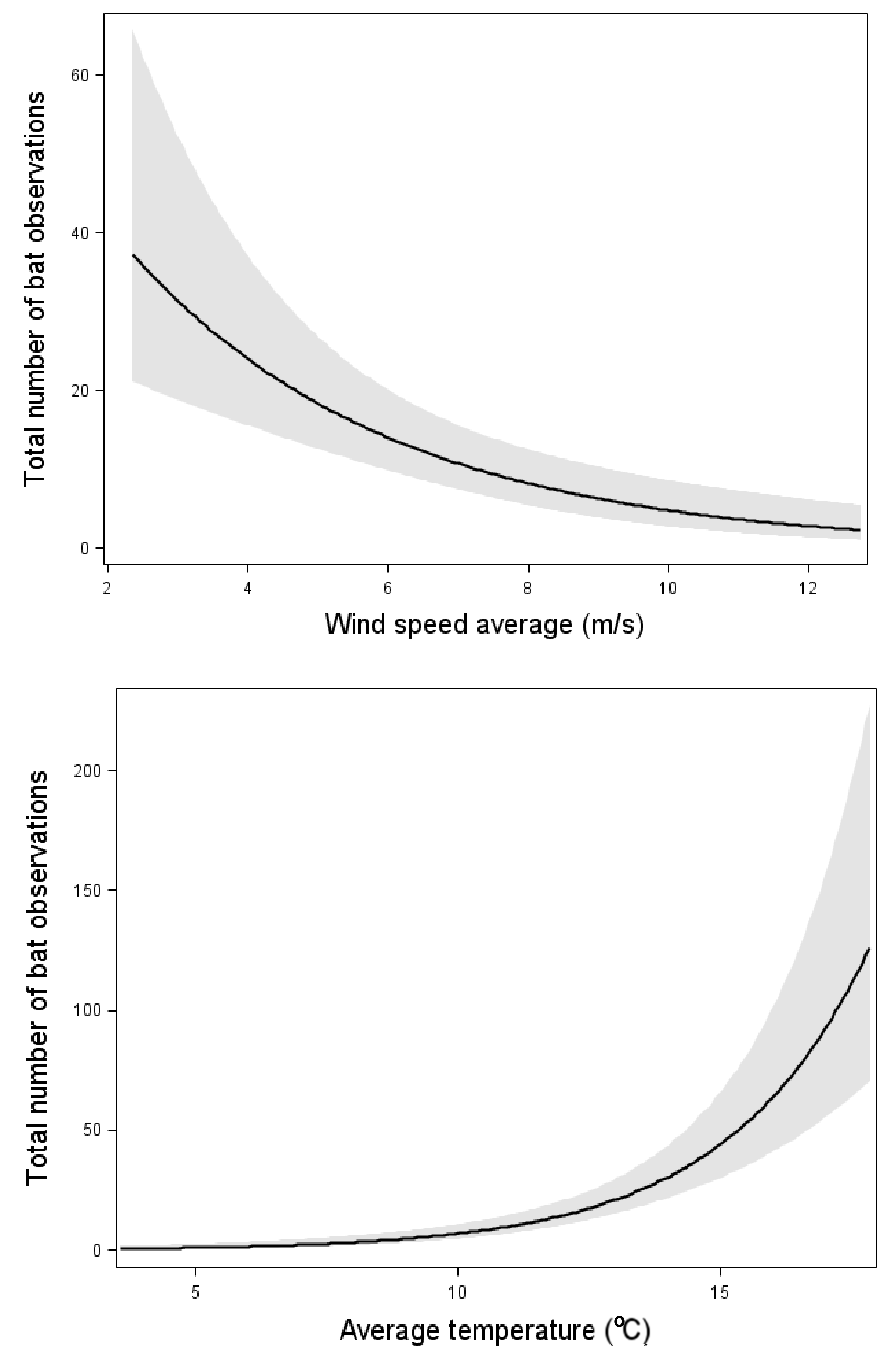
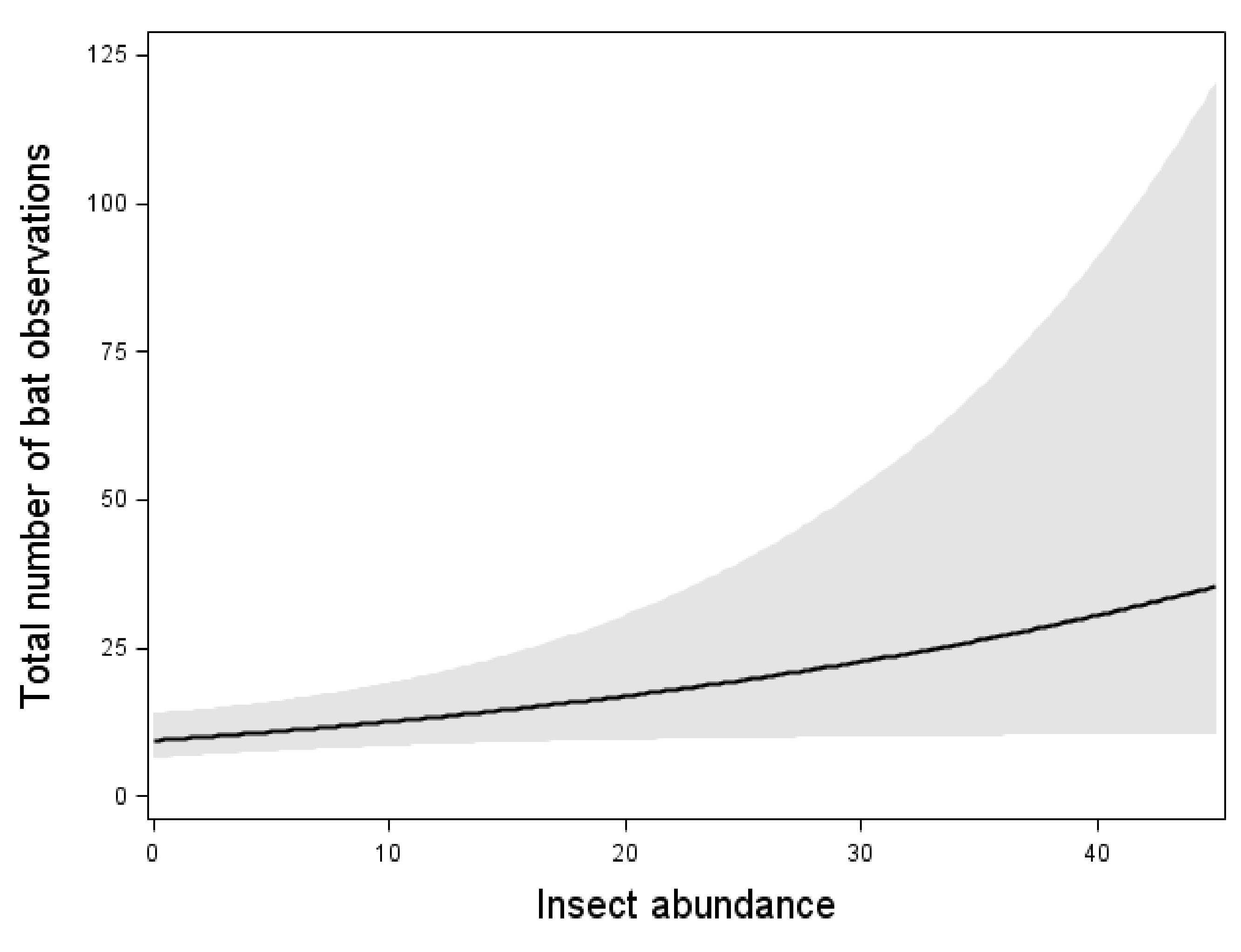
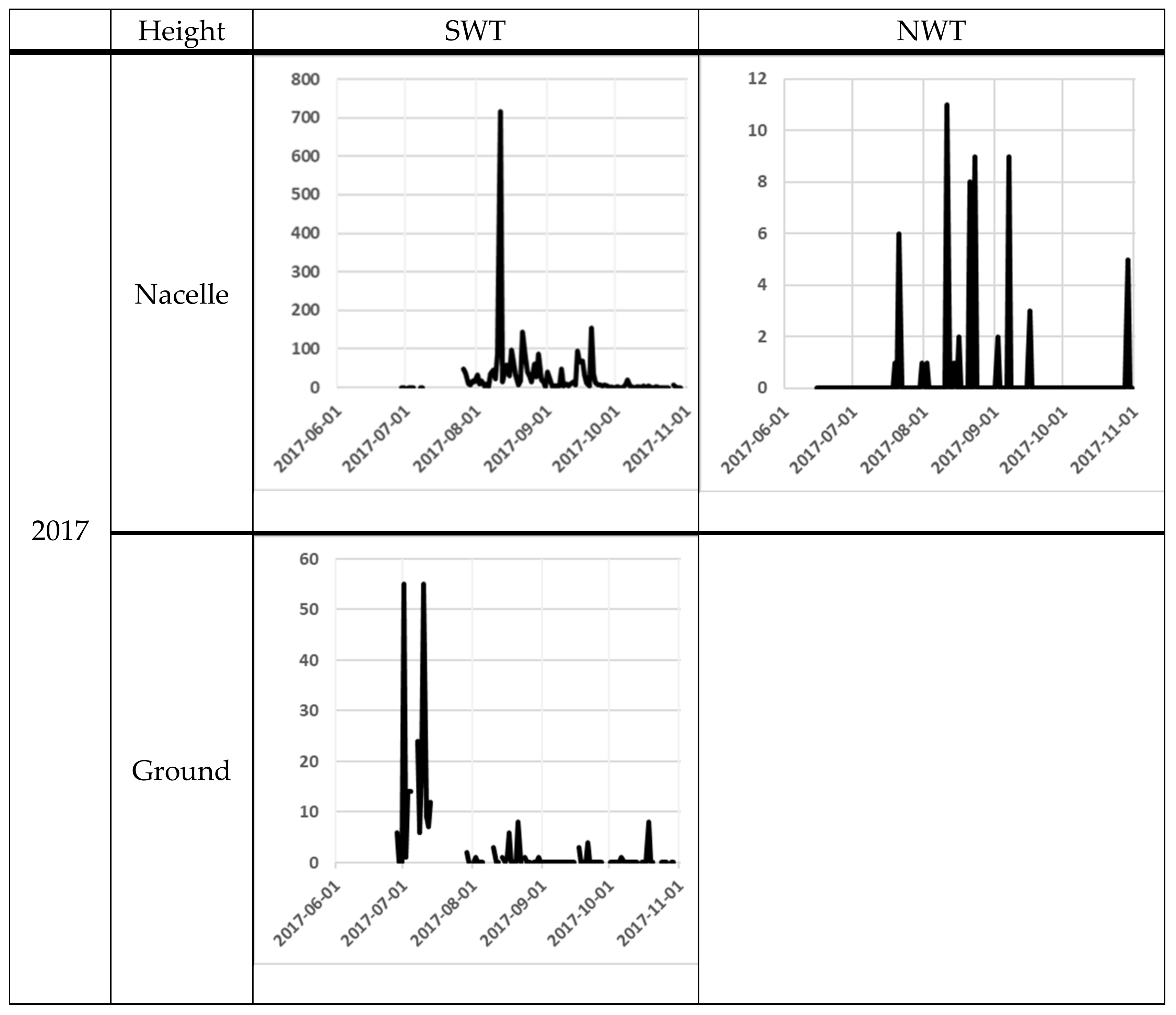

| Year | Height | Variable | SWT | NWT1 | NWT2 | Total | |
|---|---|---|---|---|---|---|---|
| Avisoft | D500X | Avisoft | Avisoft | ||||
| 2017 | Nacelle | First night | 29 June | - | 15 June | - | - |
| Last night | 29 October | - | 31 October | - | - | ||
| Number of nights | 104 | - | 139 | - | 243 | ||
| Number of recordings | 19943 | - | 6785 | - | 26728 | ||
| Total bat activity | 2718 | - | 59 | - | 2777 | ||
| Bat activity per night | 26.1 ± 7.3 | - | 0.4 ± 0.1 | - | 11.4 ± 3.2 | ||
| Ground | First night | 28 June | - | ||||
| Last night | 29 October | - | |||||
| Number of nights | 91 | - | - | - | 91 | ||
| Number of recordings | 15442 | - | - | - | 15442 | ||
| Total bat activity | 262 | - | - | - | 262 | ||
| Bat activity per night | 2.9 ± 0.9 | - | - | - | 2.9 ± 0.9 | ||
| 2018 | Nacelle | First night | 5 July | 5 July | 1 June | 1 June | - |
| Last night | 30 July | 21 October | 31 October | 31 October | - | ||
| Number of nights | 26 | 95 | 137 | 145 | 403 | ||
| Number of recordings | 19 | 157764 | 456 | 830 | 159069 | ||
| Total bat activity | 6 | 57 | 362 | 605 | 1 030 | ||
| Bat activity per night | 0.2 ± 0.1 | 0.6 ± 0.2 | 2.6 ± 1.3 | 4.2 ± 1.2 | 2.5 ± 0.6 | ||
| Ground | First night | 5 July | 5 July | - | - | - | |
| Last night | 31 October | 31 October | - | - | - | ||
| Number of nights | 119 | 114 | - | - | 233 | ||
| Number of recordings | 4000 | 30806 | - | - | 34806 | ||
| Total bat activity | 2130 | 1090 | - | - | 3220 | ||
| Bat activity per night | 17.9 ± 3.1 | 9.6 ± 1.3 | - | - | 13.8 ± 1.3 | ||
| Site | Wind speed (m/s) | Temperature (°C) | Precipitation on Rainy Nights (mm) | |||
|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017 | 2018 | 2017 | 2018 | |
| SWT | 7.3 ± 0.2 | 7.4 ± 0.2 | 11.8 ± 0.3 | 13.9 ± 0.5 | 3.1 ± 0.9 | 3.2 ± 1.4 |
| NWT1 | 6.7 ± 0.2 | 6.8 ± 0.2 | 11.7 ± 0.3 | 14.4 ± 0.4 | ||
| NWT2 | - | 6.6 ± 0.2 | - | 14.5 ± 0.4 | ||
| Variable | All Species | Pipistrellus pygmaeus | Eptesicus nilssonii1 | Myotis spp. | Nyctalus noctula | Vespertilio murinus |
|---|---|---|---|---|---|---|
| Wind | (19.85) *** | (17.24) *** | ni | (21.93) *** | (8.36) ** | ni |
| Temperature | (56.14) *** | (84.34) *** | (77.73) *** | ni | ni | (8.61) ** |
| Precipitation | ni | ni | ni | (2.10) ns | (3.21) ns | (2.86) ns |
| Insect abundance | (4.10) * | ni | (5.71) * | ni | (24.43) *** | (46.38) *** |
| Turbine (Height) | Species | 2017 | 2018 | Results from Wilcox test |
|---|---|---|---|---|
| NWT1 (nacelle-height) | E. nilssonii | 0.14 ± 0.06 | 1.20 ± 0.69 | W = 9230, p = 0.28 |
| P. pygmaeus | 0.11 ± 0.07 | 0.45 ± 0.29 | W = 9441, p = 0.72 | |
| Myotis spp. | 0 | 0 | - | |
| N. noctula | 0.13 ± 0.07 | 0.73 ± 0.36 | W = 9156, p = 0.19 | |
| V. murinus | 0.04 ± 0.08 | 0.18 ± 0.07 | W = 9167, p = 0.12 | |
| SWT (ground level) | E. nilssonii | 1. 37 ± 0.48 | 2.84 ± 0.63 | W = 3046.5, p < 0.01 |
| P. pygmaeus | 0.62 ± 0.21 | 7.77 ± 2.72 | W = 2504, p < 0.01 | |
| Myotis spp. | 0.54 ± 0.20 | 2.71 ± 0.73 | W = 2583, p < 0.01 | |
| N. noctula | 0.15 ± 0.07 | 2.37 ± 0.69 | W = 3303, p < 0.01 | |
| V. murinus | 0.15 ± 0.08 | 0.30 ± 0.09 | W = 3907, p = 0.19 |
| Year | Species | SWT | NWT1 | NWT2 | Results from Wilcox Test |
|---|---|---|---|---|---|
| 2017 | E. nilssonii | 8.13 ± 5.3 | 0.12 ± 0.07 | W = 76, p < 0.01 | |
| P. pygmaeus | 8.38 ± 1.8 | 0.14 ± 0.09 | W = 87, p < 0.01 | ||
| Myotis spp. | 7.95 ± 1.9 | 0 | W = 90, p < 0.01 | ||
| N. noctula | 0.45 ± 0.1 | 0.17 ± 0.10 | W = 60, p < 0.05 | ||
| V. murinus | 0.69 ± 0.2 | 0.06 ± 0.04 | W = 68, p < 0.01 | ||
| 2018 | E. nilssonii | 1.20 ± 0.73 | 1.74 ± 0.76 | W = 10, p < 0.05 | |
| P. pygmaeus | 0.45 ± 0.30 | 0.58 ± 0.38 | W = 95, p = 0.59 | ||
| Myotis spp. | 0 | 0 | - | ||
| N. noctula | 0.73 ± 0.38 | 1.12 ± 0.41 | W = 11, p < 0.01 | ||
| V. murinus | 0.18 ± 0.08 | 0.57 ± 0.17 | W = 10, p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Jong, J.; Millon, L.; Håstad, O.; Victorsson, J. Activity Pattern and Correlation between Bat and Insect Abundance at Wind Turbines in South Sweden. Animals 2021, 11, 3269. https://doi.org/10.3390/ani11113269
de Jong J, Millon L, Håstad O, Victorsson J. Activity Pattern and Correlation between Bat and Insect Abundance at Wind Turbines in South Sweden. Animals. 2021; 11(11):3269. https://doi.org/10.3390/ani11113269
Chicago/Turabian Stylede Jong, Johnny, Lara Millon, Olle Håstad, and Jonas Victorsson. 2021. "Activity Pattern and Correlation between Bat and Insect Abundance at Wind Turbines in South Sweden" Animals 11, no. 11: 3269. https://doi.org/10.3390/ani11113269
APA Stylede Jong, J., Millon, L., Håstad, O., & Victorsson, J. (2021). Activity Pattern and Correlation between Bat and Insect Abundance at Wind Turbines in South Sweden. Animals, 11(11), 3269. https://doi.org/10.3390/ani11113269