
Genetic Analysis of the Fatty Acid Profile in Gilthead Seabream (Sparus aurata L.)

 ,
, 
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Rearing Conditions
2.2. Flesh Composition Quality
2.3. Chromatographic Analysis of Fatty Acid Methyl Esters
2.4. Microsatellite Genotyping and Parental Assignment
2.5. Statistical Analysis
p(u|G) ~ (0; G⊗A);
p(e|R) ~ (0; R⊗A);
3. Results and Discussion
3.1. Phenotyping
3.2. Microsatellite Genotyping and Parental Assignment
3.3. Genetic Parameters
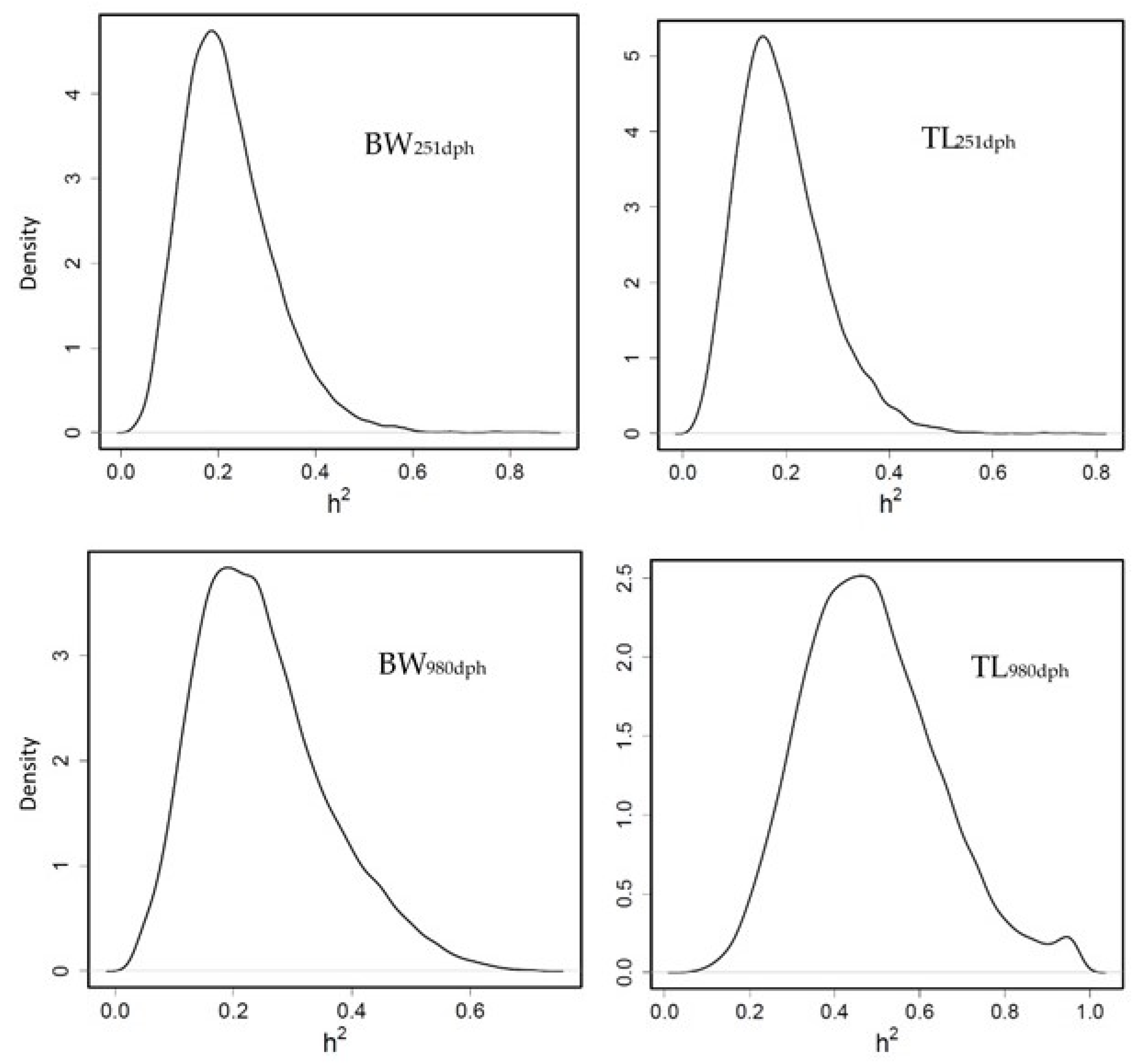
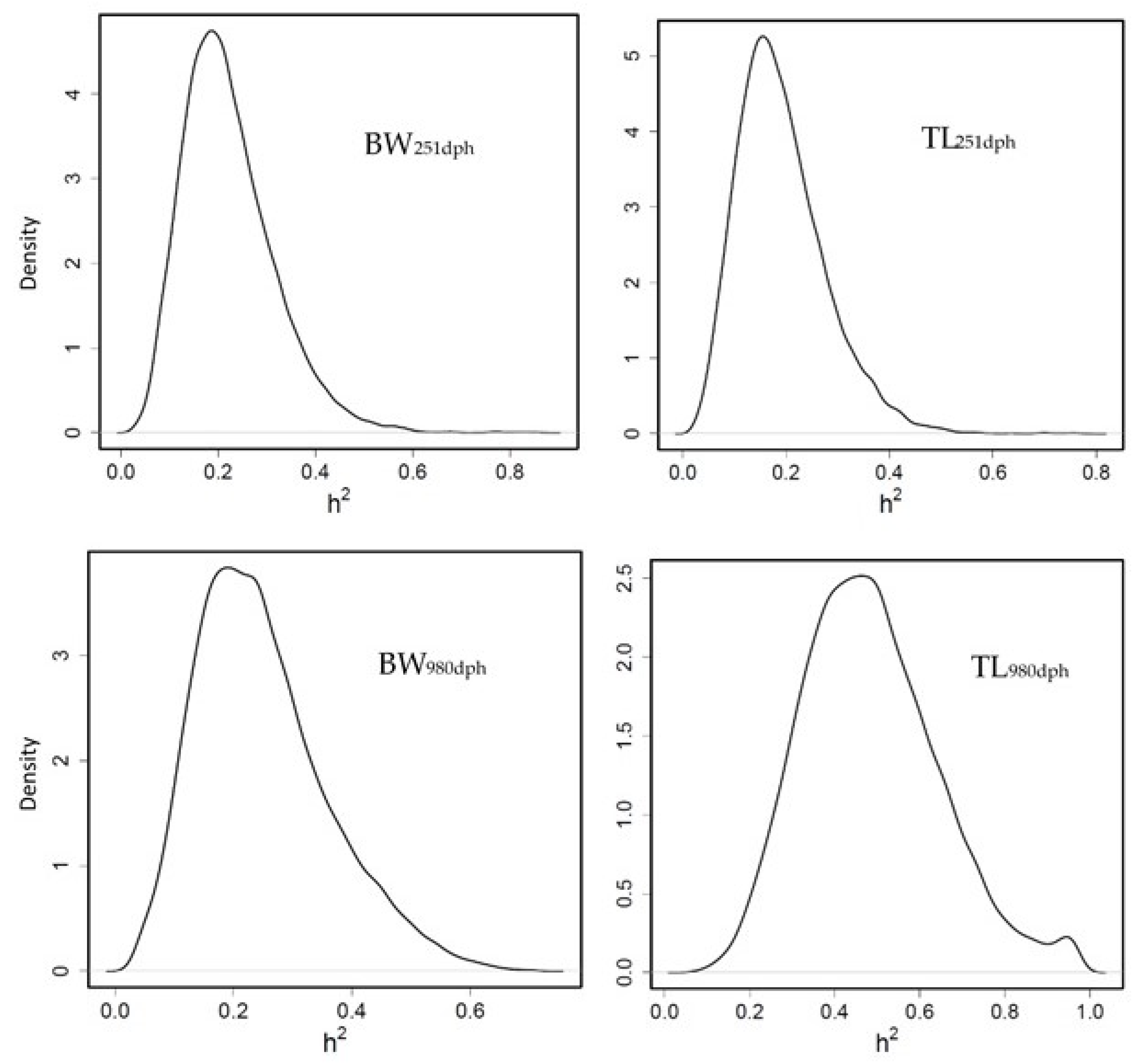
3.3.1. Heritability for Growth Traits
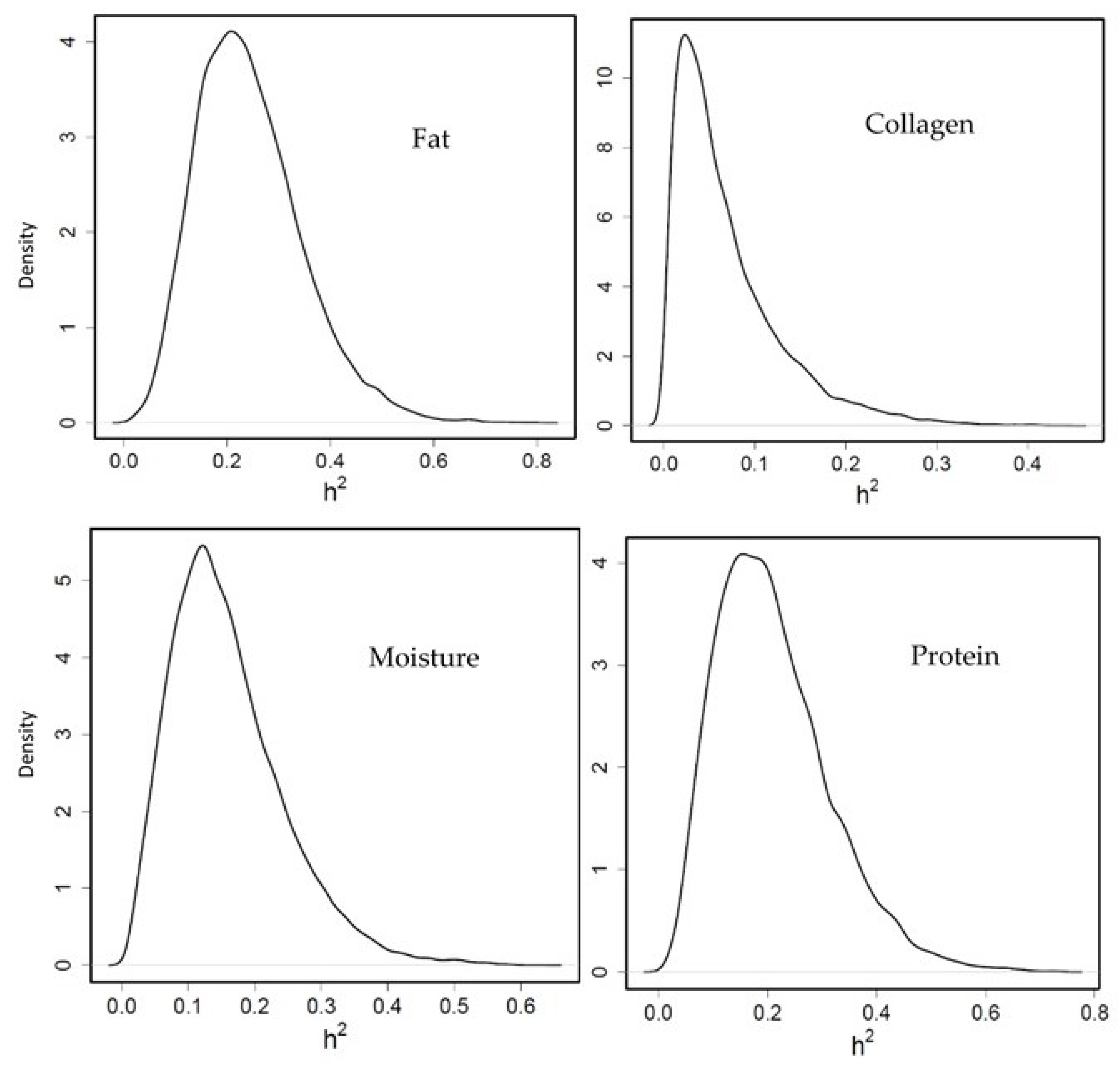
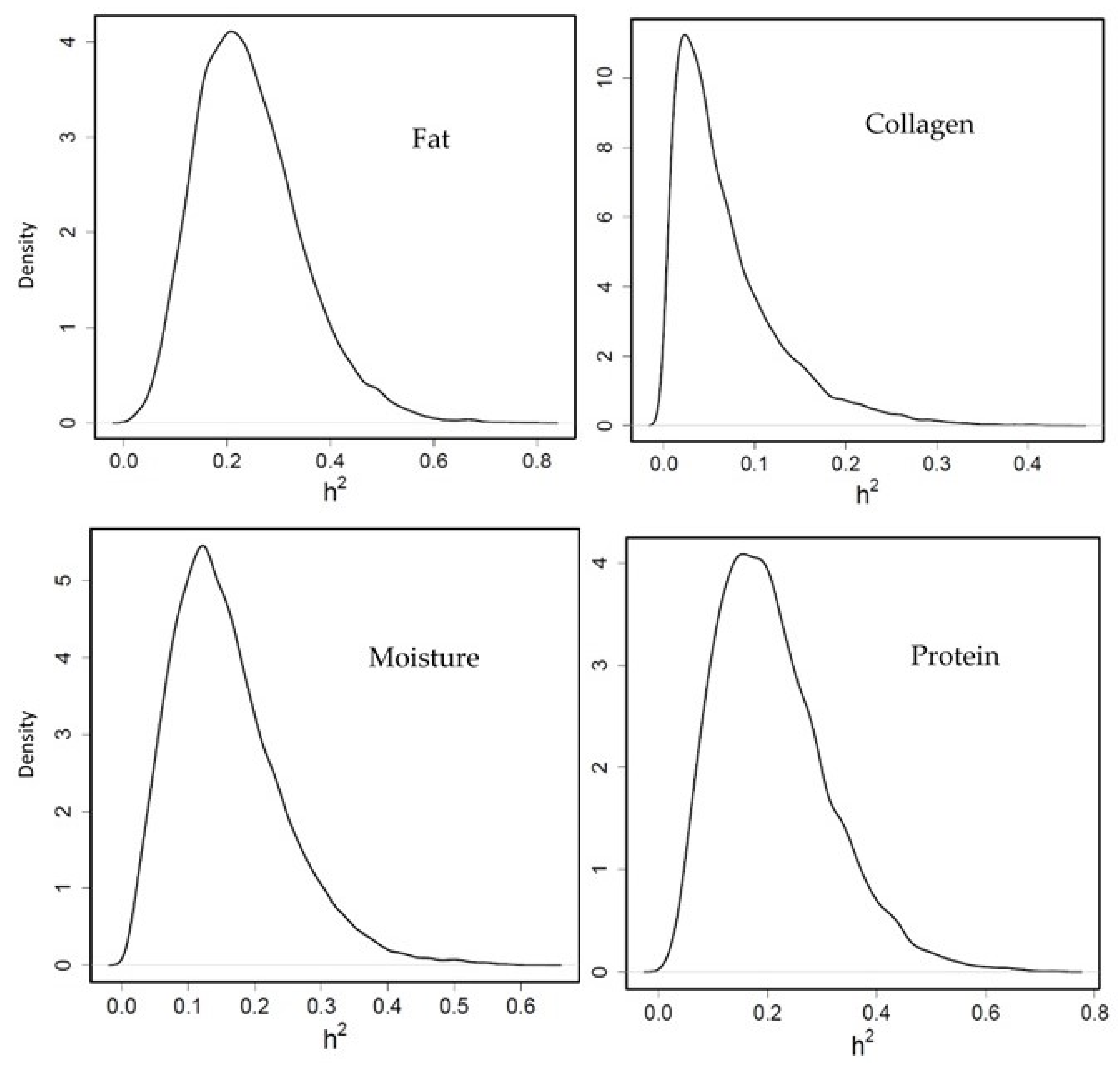
3.3.2. Heritability of Flesh Composition
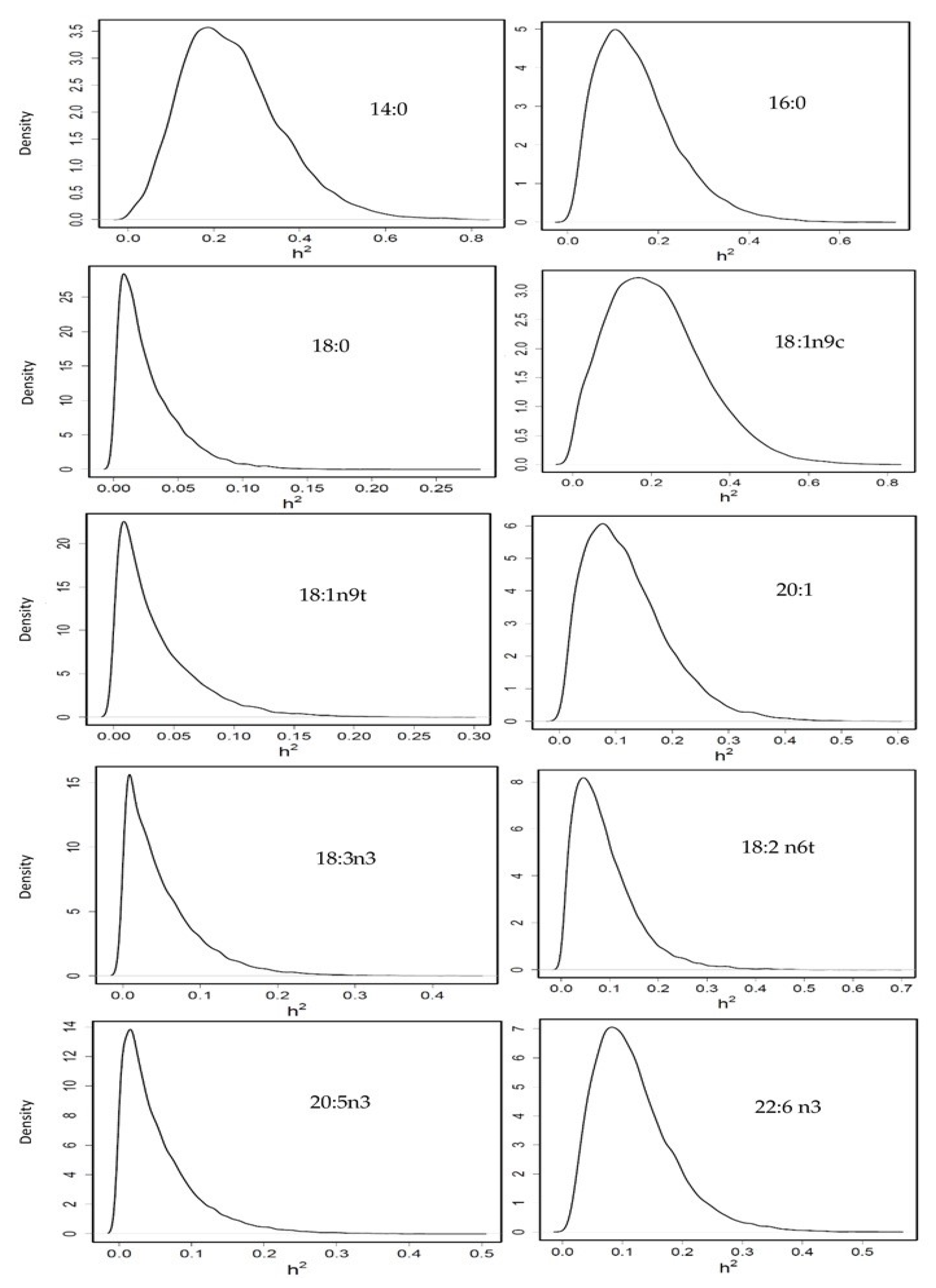
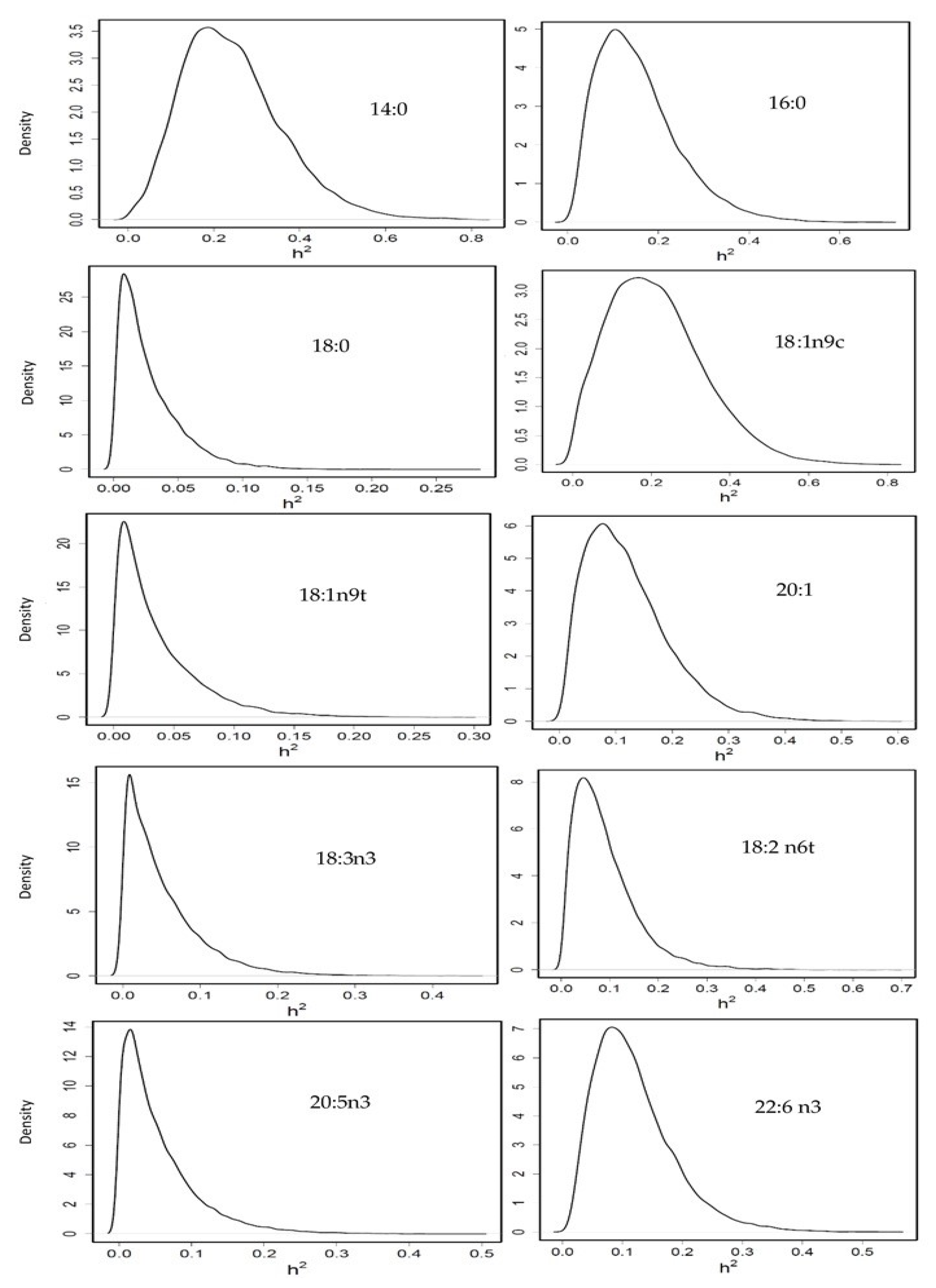
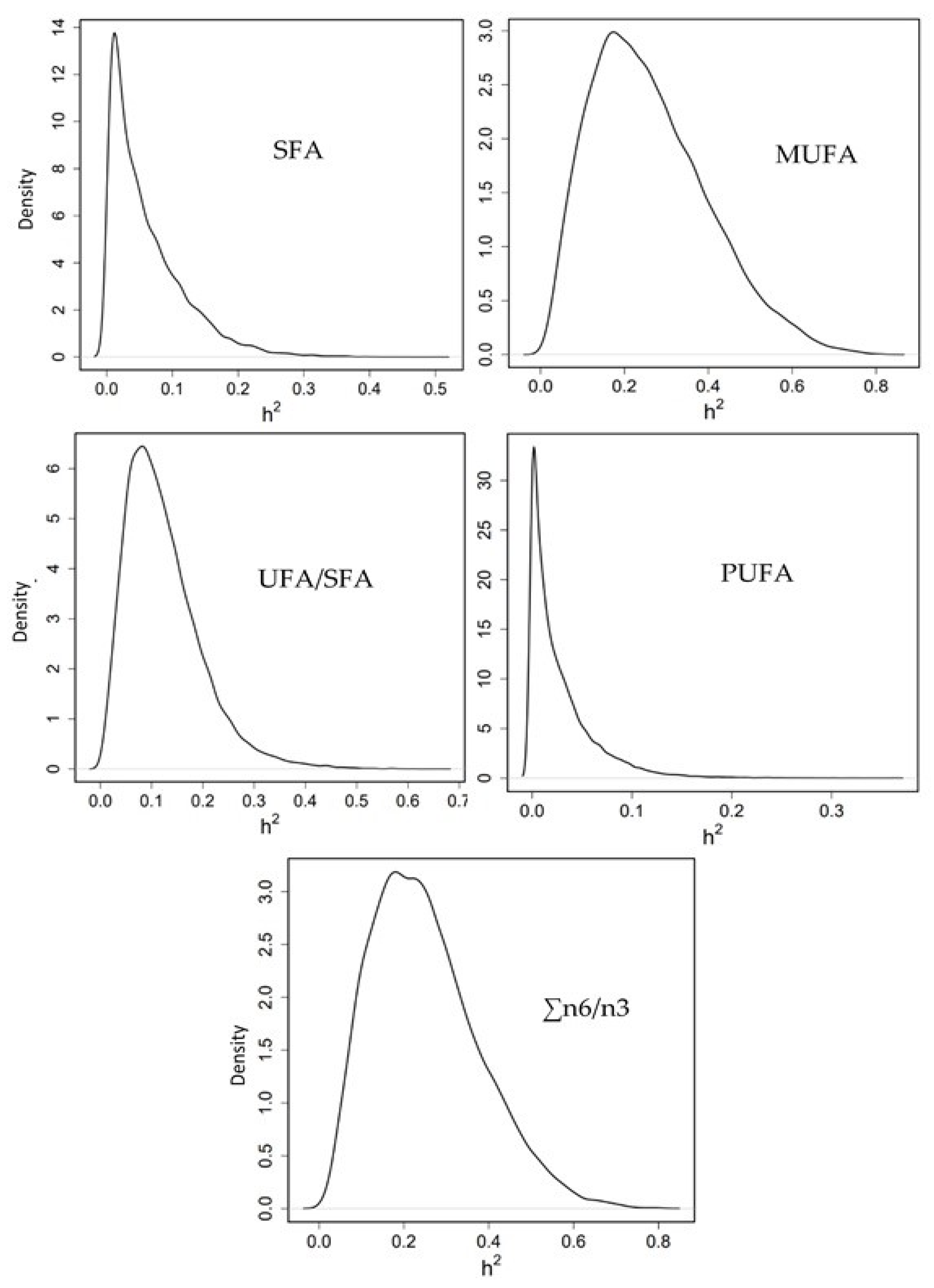
3.3.3. Heritability of Fatty Acid Profile
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- APROMAR. La Acuicultura en España; 2020; Volume 95 (In Spanish). Available online: http://www.apromar.es/ (accessed on 3 October 2021).
- FEAP. European Aquaculture Production Report; 2014. Brussels (Belgium). Available online: https://feap.info/ (accessed on 3 October 2021).
- Mccullough, M.L.; Feskanich, D.; Stampfer, M.J.; Giovannucci, E.L.; Rimm, E.B.; Hu, F.B.; Spiegelman, D.; Hunter, D.J.; Colditz, G.A.; Willett, W.C. Diet quality and major chronic disease risk in men and women: Moving toward improved dietary guidance. Am. J. Clin. Nutr. 2002, 76, 1261–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustsson, K.; Michaud, D.S.; Rimm, E.B.; Leitzmann, M.F.; Stampfer, M.J.; Willett, W.C.; Giovannucci, E. A Prospective Study of Intake of Fish and Marine Fatty Acids and Prostate Cancer. Cancer Epidemiol. Prev. Biomark. 2003, 12, 64–67. [Google Scholar]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Tocher, D.R. Omega-3 Long-Chain Polyunsaturated Fatty Acids and Aquaculture in Perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Calder, P.C. N Ϫ 3 Polyunsaturated Fatty Acids, Inflammation, and Inflammatory. Am. J. Clin. Nutr. 2006, 83, 1505S–1519S. [Google Scholar] [CrossRef]
- Yaqoob, P.; Calder, P.C. Fatty Acids and Immune Function: New Insights into Mechanisms. Br. J. Nutr. 2007, 98, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Sun, B.; Guan, W.; Bi, Y.; Li, P.; Ma, J.; Chen, F.; Pan, Q.; Xie, Q. N-3 assential fatty acids in Nile tilapia, Oreochromis niloticus: Effects of linolenic acid on non-specific immunity and anti-inflammatory responses in juveline fish. Aquaculture 2016, 450, 250–257. [Google Scholar] [CrossRef]
- Burr, G.O.; Burr, M.M. On the Nature and Role of the Fatty Acids Essential in Nutrition. J. Biogical Chem. 1930, 86, 587–621. [Google Scholar] [CrossRef]
- Rivers, J.P.W.; Frankel, T.L. Essential Fatty Acid Deficiency. Br. Med Bull. 1981, 37, 59–64. [Google Scholar] [CrossRef]
- Sciences, N. Nutritional Requirements of Fish. Nutr. Rev. 1993, 52, 417–426. [Google Scholar] [CrossRef]
- Graham, C.; Calder, P.C. Conversion of α-Linolenic Acid to Longer-Chain Polyunsaturated Fatty Acids in Human Adults. Reprod. Nutr. Dev. 2005, 45, 581–597. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty Acid Requirements in Ontogeny of Marine and Freshwater Fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Senso, L.; Suárez, M.D.; Ruiz-Cara, T.; García-Gallego, M. On the Possible Effects of Harvesting Season and Chilled Storage on the Fatty Acid Profile of the Fillet of Farmed Gilthead Sea Bream (Sparus aurata). Food Chem. 2007, 101, 298–307. [Google Scholar] [CrossRef]
- Benedito-Palos, L.; Navarro, J.C.; Sitjà-Bobadilla, A.; Gordon Bell, J.; Kaushik, S.; Pérez-Sánchez, J. High Levels of Vegetable Oils in Plant Protein-Rich Diets Fed to Gilthead Sea Bream (Sparus aurata L.): Growth Performance, Muscle Fatty Acid Profiles and Histological Alterations of Target Tissues. Br. J. Nutr. 2008, 100, 992–1003. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, M.S.; Montero, D.; Robaina, L.; Caballero, M.J.; Rosenlund, G.; Ginés, R. Alterations in Fillet Fatty Acid Profile and Flesh Quality in Gilthead Seabream (Sparus aurata) Fed Vegetable Oils for a Long Term Period. Recovery of Fatty Acid Profiles by Fish Oil Feeding. Aquaculture 2005, 250, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Ballester-Lozano, G.F.; Benedito-Palos, L.; Navarro, J.C.; Kaushik, S.; Pérez-Sánchez, J. Prediction of Fillet Fatty Acid Composition of Market-Size Gilthead Sea Bream (Sparus aurata) Using a Regression Modelling Approach. Aquaculture 2011, 319, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The Lipids, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2002; pp. 182–246. [Google Scholar]
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Sushchik, N.N.; Makhutova, O.N.; Rudchenko, A.E.; Glushchenko, L.A.; Shulepina, S.P.; Kolmakova, A.A.; Gladyshev, M.I. Comparison of Fatty Acid Contents in Major Lipid Classes of Seven Salmonid Species from Siberian Arctic Lakes. Biomolecules 2020, 10, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhutova, O.N.; Stoyanov, K.N. Fatty Acid Content and Composition in Tissues of Baikal Grayling (Thymallus baicalensis), with a Special Focus on DHA Synthesis. Aquac. Int. 2021. [Google Scholar] [CrossRef]
- Perera, E.; Simó-Mirabet, P.; Shin, H.S.; Rosell-Moll, E.; Naya-Catalá, F.; de las Heras, V.; Martos-Sitcha, J.A.; Karalazos, V.; Armero, E.; Arizcun, M.; et al. Selection for Growth Is Associated in Gilthead Sea Bream (Sparus aurata)with Diet Flexibility, Changes in Growth Patterns and Higher Intestine Plasticity. Aquaculture 2019, 507, 349–360. [Google Scholar] [CrossRef]
- Ofori-Mensah, S.; Yıldız, M.; Arslan, M.; Eldem, V.; Gelibolu, S. Substitution of Fish Oil with Camelina or Chia Oils in Gilthead Sea Bream (Sparus aurata L.) Diets: Effect on Growth Performance, Fatty Acid Composition, Haematology and Gene Expression. Aquac. Nutr. 2020, 26, 1943–1957. [Google Scholar] [CrossRef]
- Vallecillos, A.; Chaves-Pozo, E.; Arizcun, M.; Perez, R.; Afonso, J.M.; Berbel, C.; Pérez-Sánchez, J.; María-Dolores, E.; Armero, E. Genetic Parameters for Photobacterium Damselae Subsp. Piscicida Resistance, Immunological Markers and Body Weight in Gilthead Seabream (Sparus aurata). Aquaculture 2021, 543, 736892. [Google Scholar] [CrossRef]
- García-Celdrán, M.; Ramis, G.; Manchado, M.; Estévez, A.; Navarro, A.; Armero, E. Estimates of Heritabilities and Genetic Correlations of Raw Flesh Quality Traits in a Reared Gilthead Sea Bream (Sparus aurata L.) Population Sourced from Broodstocks along the Spanish Coasts. Aquaculture 2015, 446, 181–186. [Google Scholar] [CrossRef]
- Elalfy, I.; Shin, H.S.; Negrín-Báez, D.; Navarro, A.; Zamorano, M.J.; Manchado, M.; Afonso, J.M. Genetic Parameters for Quality Traits by Non-Invasive Methods and Their G x E Interactions in Ocean Cages and Estuaries on Gilthead Seabream (Sparus aurata). Aquaculture 2021, 537, 736462. [Google Scholar] [CrossRef]
- Leaver, M.J.; Taggart, J.B.; Villeneuve, L.; Bron, J.E.; Guy, D.R.; Bishop, S.C.; Houston, R.D.; Matika, O.; Tocher, D.R. Heritability and Mechanisms of N- 3 Long Chain Polyunsaturated Fatty Acid Deposition in the Flesh of Atlantic Salmon. Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Ponzoni, R.W.; Abu-Bakar, K.R.; Hamzah, A.; Khaw, H.L.; Yee, H.Y. Correlated Response in Fillet Weight and Yield to Selection for Increased Harvest Weight in Genetically Improved Farmed Tilapia (GIFT Strain), Oreochromis niloticus. Aquaculture 2010, 305, 1–5. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, S.; Chen, J.; Zhang, Q.; Liu, Y.; You, C.; Monroig, Ó.; Tocher, D.R.; Li, Y. Hepatocyte Nuclear Factor 4α (HNF4α) Is a Transcription Factor of Vertebrate Fatty Acyl Desaturase Gene as Identified in Marine Teleost Siganus canaliculatus. PLoS ONE 2016, 11, e0160361. [Google Scholar] [CrossRef]
- Dong, Y.; Zhao, J.; Chen, J.; Wang, S.; Liu, Y.; Zhang, Q.; You, C.; Monroig, Ó.; Tocher, D.R.; Li, Y. Cloning and Characterization of ∆6/∆5 Fatty Acyl Desaturase (Fad) Gene Promoter in the Marine Teleost Siganus canaliculatus. Gene 2018, 647, 174–180. [Google Scholar] [CrossRef]
- Navarro, A.; Oliva, V.; Zamorano, M.J.; Ginés, R.; Izquierdo, M.S.; Astorga, N.; Afonso, J.M. Evaluation of PIT System as a Method to Tag Fingerlings of Gilthead Seabream (Sparus auratus L.): Effects on Growth, Mortality and Tag Loss. Aquaculture 2006, 257, 309–315. [Google Scholar] [CrossRef]
- ISO 15884:2002, Milk Fat, Preparation of Fatty Acid Methyl Ester; ISO: Geneva, Switzerland, 2002.
- Lee-Montero, I.; Navarro, A.; Borrell, Y.; García-Celdrán, M.; Martín, N.; Negrín-Báez, D.; Blanco, G.; Armero, E.; Berbel, C.; Zamorano, M.J.; et al. Development of the First Standardised Panel of Two New Microsatellite Multiplex PCRs for Gilthead Seabream (Sparus aurata L.). Anim. Genet. 2013, 44, 533–546. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research-an Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandeputte, M.; Mauger, S.; Dupont-Nivet, M. An Evaluation of Allowing for Mismatches as a Way to Manage Genotyping Errors in Parentage Assignment by Exclusion. Mol. Ecol. Notes 2006, 6, 265–267. [Google Scholar] [CrossRef]
- IBM SPSS Statistics for Windows; IBM Corp: Armonk, NY, USA, 2017.
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Aguilar, I.; Legarra, A.; Vitezica, Z. BLUPF90 Family of Programs; University of Georgia: Athens, GA, USA, 2015; pp. 1–125. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/.https://feap.info/ (accessed on 3 October 2021).
- Suomela, J.P.; Lundén, S.; Kaimainen, M.; Mattila, S.; Kallio, H.; Airaksinen, S. Effects of Origin and Season on the Lipids and Sensory Quality of European Whitefish (Coregonus lavaretus). Food Chem. 2016, 197, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Fountoulaki, E.; Vasilaki, A.; Hurtado, R.; Grigorakis, K.; Karacostas, I.; Nengas, I.; Rigos, G.; Kotzamanis, Y.; Venou, B.; Alexis, M.N. Fish Oil Substitution by Vegetable Oils in Commercial Diets for Gilthead Sea Bream (Sparus aurata L.); Effects on Growth Performance, Flesh Quality and Fillet Fatty Acid Profile. Recovery of Fatty Acid Profiles by a Fish Oil Finishing Diet under Fluctuating Water Temperatures. Aquaculture 2009, 289, 317–326. [Google Scholar] [CrossRef]
- Sabbagh, M.; Schiavone, R.; Brizzi, G.; Sicuro, B.; Zilli, L.; Vilella, S. Poultry By-Product Meal as an Alternative to Fish Meal in the Juvenile Gilthead Seabream (Sparus aurata) Diet. Aquaculture 2019, 511, 734220. [Google Scholar] [CrossRef]
- Navarro, A.; Zamorano, M.J.; Hildebrandt, S.; Ginés, R.; Aguilera, C.; Afonso, J.M. Estimates of Heritabilities and Genetic Correlations for Body Composition Traits and G × E Interactions, in Gilthead Seabream (Sparus auratus L.). Aquaculture 2009, 295, 183–187. [Google Scholar] [CrossRef]
- Nolasco-Alzaga, H.R.; Perez-Enriquez, R.; Enez, F.; Bestin, A.; Palacios-Mechetnov, E.; Haffray, P. Quantitative Genetic Parameters of Growth and Fatty Acid Content in the Hemolymph of the Whiteleg Shrimp Litopenaeus Vannamei. Aquaculture 2018, 482, 17–23. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FA | D4 | D6 |
|---|---|---|
| 14:0 | 2.91 ± 0.09 | 1.89 ± 0.04 |
| 16:0 | 15.5 ± 0.11 | 11.4 ± 0.01 |
| 18:0 | 5.16 ± 0.34 | 4.31 ± 0.02 |
| SFA | 24.3 ± 0.09 | 17.5 ±0.08 |
| 16:1 | 4.05 ± 0.08 | 2.66 ± 0.00 |
| 18:1 * | 29.9 ± 0.18 | 42.9 ± 0.30 |
| C 20:1 | 3.03 ± 0.08 | 2.38 ± 0.08 |
| C 22:1 | 2.05 ± 0.32 | 1.17 ± 0.15 |
| MUFA | 39.2 ± 0.13 | 49.7 ± 0.54 |
| 18:2n6 | 18.9 ± 0.21 | 20.1 ± 0.31 |
| 18:3n3 | 4.61 ± 0.26 | 6.05 ± 0.03 |
| 20:4n6 | 1.18 ± 0.59 | 0.00 ± 0.00 |
| 20:5n3 | 4.58 ± 0.03 | 2.81 ± 1.40 |
| 22:6n3 | 6.19 ± 0.52 | 4.01 ± 0.06 |
| PUFA | 36.4 ± 0.04 | 32.6 ± 0.45 |
| Offspring | Covariate BW980dph | ||||
|---|---|---|---|---|---|
| Traits | n | Mean | S.E. | b | S.E. |
| BW251dph (g) | 392 | 43.7 | 0.95 | NI | |
| TL251dph (cm) | 392 | 13.8 | 0.09 | NI | |
| BW980dph (g) | 392 | 450.2 | 4.14 | NI | |
| TL980dph (cm) | 392 | 28.7 | 0.11 | NI | |
| Fat (%) | 389 | 10.1 | 0.12 | 0.006 * | 0.002 |
| Collagen (%) | 388 | 1.74 | 0.02 | 0.001 * | 0.000 |
| Moisture (%) | 389 | 64.6 | 0.11 | −0.007 * | 0.001 |
| Protein (%) | 389 | 19.8 | 0.06 | <0.000 | 0.001 |
| FA | Offspring | Covariate Fat Percentage | |||
|---|---|---|---|---|---|
| n | LSM | S.E. | b | S.E. | |
| 14:0 | 394 | 3.29 | 0.03 | 0.011 | 0.015 |
| 16:0 | 371 | 17.9 | 0.23 | 0.018 | 0.090 |
| 18:0 | 392 | 4.86 | 0.11 | −0.019 | 0.045 |
| SFA | 397 | 28.3 | 0.30 | −0.002 | 0.130 |
| 16:1 | 397 | 0.32 | 0.04 | −0.018 | 0.015 |
| 18:1n9c | 392 | 33.6 | 0.70 | 0.232 | 0.277 |
| 18:1n9t | 333 | 3.55 | 0.11 | −0.005 | 0.047 |
| 20:1 | 392 | 2.38 | 0.03 | −0.004 | 0.014 |
| 22:1 | 397 | 0.80 | 0.02 | −0.014 | 0.008 |
| MUFA | 397 | 44.38 | 0.57 | 0.291 | 0.209 |
| 18:2n6 | 334 | 15.6 | 0.21 | −0.054 | 0.084 |
| 18:3n3 | 389 | 3.77 | 0.05 | 0.055 * | 0.022 |
| 20:4n6 | 399 | 0.63 | 0.02 | −0.017 | 0.006 |
| 20:5n3 | 381 | 2.88 | 0.04 | 0.008 | 0.016 |
| 22:6n3 | 384 | 6.84 | 0.12 | −0.097 * | 0.048 |
| PUFA | 397 | 29.84 | 045 | −0.400 | 0.165 |
| ∑ n6/n3 | 394 | 1.05 | 0.35 | 0.249 | 0.140 |
| UFA/SFA | 392 | 2.97 | 0.08 | −0.029 | 0.034 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vallecillos, A.; Marín, M.; Bortoletti, M.; López, J.; Afonso, J.M.; Ramis, G.; Arizcun, M.; María-Dolores, E.; Armero, E. Genetic Analysis of the Fatty Acid Profile in Gilthead Seabream (Sparus aurata L.). Animals 2021, 11, 2889. https://doi.org/10.3390/ani11102889
Vallecillos A, Marín M, Bortoletti M, López J, Afonso JM, Ramis G, Arizcun M, María-Dolores E, Armero E. Genetic Analysis of the Fatty Acid Profile in Gilthead Seabream (Sparus aurata L.). Animals. 2021; 11(10):2889. https://doi.org/10.3390/ani11102889
Chicago/Turabian StyleVallecillos, Antonio, María Marín, Martina Bortoletti, Javier López, Juan M. Afonso, Guillermo Ramis, Marta Arizcun, Emilio María-Dolores, and Eva Armero. 2021. "Genetic Analysis of the Fatty Acid Profile in Gilthead Seabream (Sparus aurata L.)" Animals 11, no. 10: 2889. https://doi.org/10.3390/ani11102889
APA StyleVallecillos, A., Marín, M., Bortoletti, M., López, J., Afonso, J. M., Ramis, G., Arizcun, M., María-Dolores, E., & Armero, E. (2021). Genetic Analysis of the Fatty Acid Profile in Gilthead Seabream (Sparus aurata L.). Animals, 11(10), 2889. https://doi.org/10.3390/ani11102889