
The Recent Trend in the Use of Multistrain Probiotics in Livestock Production: An Overview


Simple Summary
Abstract
1. Introduction
2. Common Probiotic Strains and Their Mode of Action
- (1)
- Competitive exclusion,
- (2)
- Bacterial antagonism, and
- (3)
- Immune system stimulation.
3. Advent of Combining Microorganisms
4. Multistrain Probiotic Use in Ruminants
5. Multistrain Probiotic Use in Poultry
6. Multistrain Probiotic Use in Swine
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, Y.; Cheng, H.; Tao, S. Environmental and human health challenges of industrial livestock and poultry farming in China and their mitigation. Environ. Int. 2017, 107, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Regulation 1831/2003/EC on Additives for Use in Animal Nutrition, Replacing Directive 70/524/EEC on Additives in Feeding-Stuffs. Official Journal of the European Union. 2003. Available online: http://ec.europa.eu/health/ph/others/antimicrob_resist/am_02_en.pdf (accessed on 3 September 2021).
- Cholewińska, P.; Czyż, K.; Nowakowski, P.; Wyrostek, A. The microbiome of the digestive system of ruminants—A review. Anim. Health Res. Rev. 2020, 21, 3–14. [Google Scholar] [CrossRef]
- Khan, R.U.; Shabana, N.; Kuldeep, D.; Karthik, K.; Ruchi, T.; Abdelrahman, M.M.; Alhidary, I.A.; Arshad, Z. Direct-Fed Microbial: Beneficial Applications, Modes of Action andProspects as a Safe Tool for Enhancing Ruminant Production andSafeguarding Health. Int. J. Pharmacol. 2016, 12, 220–231. [Google Scholar] [CrossRef]
- Quigley, J. Direct-fed microbials (probiotics) in calf diets. Bovine Alliance on management and Nutrition. 2011. Available online: http://nahms.aphis.usda.gov/dairy/ (accessed on 6 September 2021).
- Brashears, M.M.; Amezquita, A.; Jaroni, D. Lactic Acid Bacteria and Their Uses in Animal Feeding to Improve Food Safety. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2005; Volume 50, pp. 1–31. [Google Scholar] [CrossRef]
- Lilly, D.M.; Stillwell, R.H. Probiotics: Growth-Promoting Factors Produced by Microorganisms. Science 1965, 147, 747–748. [Google Scholar] [CrossRef]
- Bielecka, M. Probiotics in Food. In Chemical and Functional Properties of Food Components, 3rd ed.; Sikorski, Z., Ed.; CRC Press: Boca Raton, FL, USA, 2006; Volume 20061236, pp. 413–426. [Google Scholar] [CrossRef]
- Ahmed, S.T.; Islam, M.M.; Mun, H.-S.; Sim, H.-J.; Kim, Y.-J.; Yang, C.-J. Effects ofBacillus amyloliquefaciens as a probiotic strain on growth performance, cecal microflora, and fecal noxious gas emissions of broiler chickens. Poult. Sci. 2014, 93, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Amoah, K.; Huang, Q.C.; Tan, B.P.; Zhang, S.; Chi, S.Y.; Yang, Q.H.; Liu, H.Y.; Dong, X.H. Dietary supplementation of probiotic Bacillus coagulans ATCC 7050, improves the growth performance, intestinal morphology, microflora, immune response, and disease confrontation of Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 87, 796–808. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations and World Health Organization. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation; Food and Agriculture Organization of the United Nations, World Health Organization: Rome, Italy, 2006. [Google Scholar]
- Jiménez, G. Probiotics in Animal Nutrition—A Century of Research. 2012. Available online: https://www.allaboutfeed.net/animal-feed/feed-additives/probiotics-in-animal-nutrition-a-century-of-research/ (accessed on 6 September 2021).
- Yirga, H. The Use of Probiotics in Animal Nutrition. J. Probiotics Health 2015, 03. [Google Scholar] [CrossRef]
- Collado, M.C.; Gueimonde, M.; Hernández, M.; Sanz, Y.; Salminen, S. Adhesion of Selected Bifidobacterium Strains to Human Intestinal Mucus and the Role of Adhesion in Enteropathogen Exclusion. J. Food Prot. 2005, 68, 2672–2678. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef]
- Abdelqader, A.; Irshaid, R.; Al-Fataftah, A.-R. Effects of dietary probiotic inclusion on performance, eggshell quality, cecal microflora composition, and tibia traits of laying hens in the late phase of production. Trop. Anim. Health Prod. 2013, 45, 1017–1024. [Google Scholar] [CrossRef]
- Mookiah, S.; Sieo, C.C.; Ramasamy, K.; Abdullah, N.; Ho, Y.W. Effects of dietary prebiotics, probiotic and synbiotics on performance, caecal bacterial populations and caecal fermentation concentrations of broiler chickens: Effects of dietary prebiotics, probiotic and synbiotics on performance. J. Sci. Food Agric. 2014, 94, 341–348. [Google Scholar] [CrossRef]
- Pedroso, A.A.; Hurley-Bacon, A.L.; Zedek, A.S.; Kwan, T.W.; Jordan, A.P.; Avellaneda, G.; Hofacre, C.L.; Oakley, B.B.; Collett, S.R.; Maurer, J.J.; et al. Can Probiotics Improve the Environmental Microbiome and Resistome of Commercial Poultry Production? Int. J. Environ. Res. Public. Health 2013, 10, 4534–4559. [Google Scholar] [CrossRef]
- van Doan, H.; Hoseinifar, S.H.; Dawood, M.A.O.; Chitmanat, C.; Tayyamath, K. Effects of Cordyceps militaris spent mushroom substrate and Lactobacillus plantarum on mucosal, serum immunology and growth performance of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 70, 87–94. [Google Scholar] [CrossRef]
- Zorriehzahra, M.J.; Delshad, S.T.; Adel, M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: An update on their multiple modes of action: A review. Vet. Q. 2016, 36, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Lambo, M.T.; Liu, D.; Li, X. The study of the potential application of nanofiber microcapsules loading lactobacillus in targeted delivery of digestive tract in vitro. LWT 2021, 148, 111692. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef]
- Shewale, R.N.; Sawale, P.D.; Khedkar, C.D.; Singh, A. Selection criteria for probiotics: A review. Int. J. Probiotics Prebiotics 2014, 9, 17–22. [Google Scholar]
- Binda, S.; Hill, C.; Johansen, E.; Obis, D.; Pot, B.; Sanders, M.E.; Tremblay, A.; Ouwehand, A.C. Criteria to Qualify Microorganisms as ‘Probiotic’ in Foods and Dietary Supplements. Front. Microbiol. 2020, 11, 1662. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Parker, C.; Tindall, B.; Garrity, G. International Code of Nomenclature of Prokaryotes: Prokaryotic Code (2008 Revision). Int. J. Syst. Evol. Microbiol. 2019, 69, S1–S111. [Google Scholar] [CrossRef]
- Fajardo, P.; Pastrana, L.; Méndez, J.; Rodríguez, I.; Fuciños, C.; Guerra, N.P. Effects of Feeding of Two Potentially Probiotic Preparations from Lactic Acid Bacteria on the Performance and Faecal Microflora of Broiler Chickens. Sci. World J. 2012, 2012, 1–9. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Sun, Y.-Z.; Wang, A.; Zhou, Z. Probiotics as Means of Diseases Control in Aquaculture, a Review of Current Knowledge and Future Perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Ohinaim, E.I.; Ofongo, R.T.S. The Effect of Probiotic and Prebiotic Feed Supplementationon Chicken Health and Gut Microflora: A Review. Int. J. Anim. Vet. Adv. 2012, 4, 135–143. [Google Scholar]
- Zeng, W.; Shen, J.; Bo, T.; Peng, L.; Xu, H.; Nasser, M.I.; Zhuang, Q.; Zhao, M. Cutting Edge: Probiotics and Fecal Microbiota Transplantation in Immunomodulation. J. Immunol. Res. 2019, 2019, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Yang, X.J.; Li, Z.J.; Sun, F.F.; Wu, X.H.; Yao, J.H. Reduced lesions in chickens with Clostridium perfringens-induced necrotic enteritis by Lactobacillus fermentum 1.2029. Poult. Sci. 2012, 91, 3065–3071. [Google Scholar] [CrossRef] [PubMed]
- Chaves, B.D.; Brashears, M.M.; Nightingale, K.K. Applications and safety considerations of Lactobacillus salivarius as a probiotic in animal and human health. J. Appl. Microbiol. 2017, 123, 18–28. [Google Scholar] [CrossRef]
- Safari, R.; Adel, M.; Lazado, C.C.; Caipang, C.M.A.; Dadar, M. Host-derived probiotics Enterococcus casseliflavus improves resistance against Streptococcus iniae infection in rainbow trout (Oncorhynchus mykiss) via immunomodulation. Fish Shellfish Immunol. 2016, 52, 198–205. [Google Scholar] [CrossRef]
- Arsène, M.M.; Davares, A.K.; Andreevna, S.L.; Vladimirovich, E.A.; Carime, B.Z.; Marouf, R.; Khelifi, I. The use of probiotics in animal feeding for safe production and as potential alternatives to antibiotics. Vet. World 2021, 14, 319–328. [Google Scholar] [CrossRef]
- Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.; Stuer-Lauridsen, B.; Eskesen, D. The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms 2014, 2, 92–110. [Google Scholar] [CrossRef]
- Liao, S.F.; Nyachoti, M. Using probiotics to improve swine gut health and nutrient utilization. Anim. Nutr. 2017, 3, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 31. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, H.M.; Koning, C.J.M.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Mackowiak, P.A. Recycling Metchnikoff: Probiotics, the Intestinal Microbiome and the Quest for Long Life. Front. Public Health 2013, 1, 52. [Google Scholar] [CrossRef]
- Sanders, M.E. Summary of Conclusions from a Consensus panel of Experts on health Attributes to Lactic Cultures: Significance to Fluid Milk Products Containing Cultures. J. Dairy Sci. 1993, 76, 1819–1828. [Google Scholar] [CrossRef]
- Famularo, G.; de Simone, C.; Matteuzzi, D.; Pirovano, F. Traditional and High Potency Probiotic Preparations for oral Bacteriotherapy. Biodrugs 1999, 12, 455–470. [Google Scholar] [CrossRef]
- Dunne, C.; Murphy, L.; Flynn, S.; O’Mahony, L.; O’Halloran, S.; Feeney, M.; Morrissey, D.; Thornton, G.; Fitzgerald, G.; Daly, C.; et al. Probiotics: From myth to reality. Demonstration of functionality in animal models of disease and in human clinical trials. Antonie Van Leeuwenhoek 1999, 76, 79–292. [Google Scholar] [CrossRef]
- Rolfe, R.D. The Role of Probiotic Cultures in the Control of Gastrointestinal Health. J. Nutr. 2000, 130, 396S–402S. [Google Scholar] [CrossRef]
- Kwoji, I.D.; Aiyegoro, O.A.; Okpeku, M.; Adeleke, M.A. Multistrain Probiotics: Synergy among Isolates Enhances Biological Activities. Biology 2021, 10, 322. [Google Scholar] [CrossRef]
- Douillard, F.P.; Mora, D.; Eijlander, R.T.; Wels, M.; de Vos, W.M. Comparative genomic analysis of the multispecies probiotic-marketed product VSL#3. PLoS ONE 2018, 13, e0192452. [Google Scholar] [CrossRef]
- Hammami, R.; Fernandez, B.; Lacroix, C.; Fliss, I. Anti-infective properties of bacteriocins: An update. Cell. Mol. Life Sci. 2013, 70, 2947–2967. [Google Scholar] [CrossRef]
- Oman, T.J.; van der Donk, W.A. Insights into the Mode of Action of the Two-Peptide Lantibiotic Haloduracin. ACS Chem. Biol. 2009, 4, 865–874. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Abdel-Daim, M.M.; van Doan, H. Probiotic application for sustainable aquaculture. Rev. Aquac. 2019, 11, 907–924. [Google Scholar] [CrossRef]
- Melo-Bolívar, J.F.; Pardo, R.Y.R.; Hume, M.E.; Díaz, L.M.V. Multistrain probiotics use in main commercially cultured freshwater fish: A systematic review of evidence. Rev. Aquac. 2021, raq.12543. [Google Scholar] [CrossRef]
- Ringø, E.; Hoseinifar, S.H.; Ghosh, K.; Doan, H.V.; Beck, B.R.; Song, S.K. Lactic Acid Bacteria in Finfish—An Update. Front. Microbiol. 2018, 9, 1818. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Huis, J. Bringing a probiotic-containing functional food to the market: Microbiological, product, regulatory and labeling issues. Antonie Van Leeuwenhoek 1999, 76, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Chapman, C.M.C.; Gibson, G.R.; Todd, S.; Rowland, I. Comparative in vitro inhibition of urinary tract pathogens by single- and multistrain probiotics. Eur. J. Nutr. 2013, 52, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H. Biosorption of Heavy Metals by Lactic Acid Bacteria for Detoxification. In Lactic Acid Bacteria 1887; Kanauchi, M., Ed.; Springer: New York, NY, USA, 2019; pp. 145–157. [Google Scholar] [CrossRef]
- Astolfi, M.L.; Protano, C.; Schiavi, E.; Marconi, E.; Capobianco, D.; Massimi, L.; Ristorini, M.; Baldassarre, M.E.; Laforgia, N.; Vitali, M.; et al. A prophylactic multistrain probiotic treatment to reduce the absorption of toxic elements: In-vitro study and biomonitoring of breast milk and infant stools. Environ. Int. 2019, 130, 104818. [Google Scholar] [CrossRef]
- Daisley, B.A.; Monachese, M.; Trinder, M.; Bisanz, J.E.; Chmiel, J.A.; Burton, J.P.; Reid, G. Immobilization of cadmium and lead by Lactobacillus rhamnosus GR-1 mitigates apical-to-basolateral heavy metal translocation in a Caco-2 model of the intestinal epithelium. Gut Microbes 2019, 10, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Fredua-Agyeman, M.; Stapleton, P.; Basit, A.W.; Gaisford, S. Microcalorimetric evaluation of a multistrain probiotic: Interspecies inhibition between probiotic strains. J. Funct. Foods 2017, 36, 357–361. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Mulder, L.; Everts, H.; Van Espen, D.C.; Van Der Wal, E.; Klaassen, G.; Rouwers, S.M.; Hartemink, R.; Rombouts, F.M.; Beynen, A.C. Health and Growth of Veal Calves Fed Milk Replacers With or Without Probiotics. J. Dairy Sci. 2005, 88, 2154–2165. [Google Scholar] [CrossRef]
- Giannenas, I.; Papadopoulos, E.; Tsalie, E.; Triantafillou, E.L.; Henikl, S.; Teichmann, K.; Tontis, D. Assessment of dietary supplementation with probiotics on performance, intestinal morphology and microflora of chickens infected with Eimeria tenella. Vet. Parasitol. 2012, 188, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Sykiotis, G.P.; Nishimura, M.; Bodmer, R.; Bohmann, D. Declining signal dependence of N rf2- M af S -regulated gene expression correlates with aging phenotypes. Aging Cell 2013, 12, 554–562. [Google Scholar] [CrossRef]
- Monteverde, V.; Congiu, F.; Vazzana, I.; Dara, S.; di Pietro, S.; Piccione, G. Serum lipid profile modification related to polyunsaturated fatty acid supplementation in thoroughbred horses. J. Appl. Anim. Res. 2017, 45, 615–618. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, Y.; Xu, Q.; Zheng, N.; Zhao, S.; Huang, G.; Wang, J. Ruminal microbiota–host interaction and its effect on nutrient metabolism. Anim. Nutr. 2021, 7, 49–55. [Google Scholar] [CrossRef]
- O’Hara, E.; Neves, A.L.A.; Song, Y.; Guan, L.L. The Role of the Gut Microbiome in Cattle Production and Health: Driver or Passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef]
- Kritas, S.K.; Govaris, A.; Christodoulopoulos, G.; Burriel, A.R. Effect of Bacillus licheniformis and Bacillus subtilis Supplementation of Ewe’s Feed on Sheep Milk Production and Young Lamb Mortality. J. Vet. Med. Ser. A 2006, 53, 170–173. [Google Scholar] [CrossRef]
- Adjei-Fremah, S.; Ekwemalor, K.; Asiamah, E.K.; Ismail, H.; Ibrahim, S.; Worku, M. Effect of probiotic supplementation on growth and global gene expression in dairy cows. J. Appl. Anim. Res. 2018, 46, 257–263. [Google Scholar] [CrossRef]
- Whitley, N.C.; Cazac, D.; Rude, B.J.; Jackson-O’Brien, D.; Parveen, S. Use of a commercial probiotic supplement in meat goats1. J. Anim. Sci. 2009, 87, 723–728. [Google Scholar] [CrossRef]
- Hasunuma, T.; Kawashima, K.; Nakayama, H.; Murakami, T.; Kanagawa, H.; Ishii, T.; Akiyama, K.; Yasuda, K.; Terada, F.; Kushibiki, S. Effect of cellooligosaccharide or synbiotic feeding on growth performance, fecal condition and hormone concentrations in Holstein calves: Cellobiose or Synbiotic Feeding in Calves. Anim. Sci. J. 2011, 82, 543–548. [Google Scholar] [CrossRef]
- Vibhute, V.; Shelke, R.; Chavan, S.; Nage, S. Effect of Probiotics Supplementation on the Performance of Lactating Crossbred Cows. Vet. World 2011, 4, 557. [Google Scholar] [CrossRef]
- Renaud, D.L.; Kelton, D.F.; Weese, J.S.; Noble, C.; Duffield, T.F. Evaluation of a multispecies probiotic as a supportive treatment for diarrhea in dairy calves: A randomized clinical trial. J. Dairy Sci. 2019, 102, 4498–4505. [Google Scholar] [CrossRef]
- Rao, Y.Y.N.K.A.; Kumar, C.V.S.D.S.; Lekha, M.S. Effect of Feeding Multi-Strain Probiotic on Feed Intake and Milk Production Performance in Murrah Buffaloes. Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 409–417. [Google Scholar] [CrossRef]
- Kembabazi, B.; Ondiek, J.O.; Migwi, P.K. Effect of single or mixed strain probiotics on milk yield of dairy cows. Livest. Res. Rural Dev. 2021, 33. Available online: http://www.lrrd.org/lrrd33/1/brend3307.html (accessed on 4 August 2021).
- Thomas, A.D. Supplementation of Two Novel Probiotics in the Dietof Lactating Dairy Cows. Iowa State University. 2017. Available online: http://lib.dr.iastate.edu/etd/16110 (accessed on 4 August 2021).
- Olchowy, T.W.J.; Soust, M.; Alawneh, J. The effect of a commercial probiotic product on the milk quality of dairy cows. J. Dairy Sci. 2019, 102, 2188–2195. [Google Scholar] [CrossRef]
- Deng, Q.; Odhiambo, J.F.; Farooq, U.; Lam, T.; Dunn, S.M.; Ametaj, B.N. Intravaginal probiotics modulated metabolic status and improved milk production and composition of transition dairy cows1. J. Anim. Sci. 2016, 94, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Cao, G.T.; Ferket, P.R.; Liu, T.T.; Zhou, L.; Zhang, L.; Xiao, Y.P.; Chen, A.G. Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens. Poult. Sci. 2012, 91, 2121–2129. [Google Scholar] [CrossRef] [PubMed]
- Dlamini, Z.C.; Langa, R.L.S.; Aiyegoro, O.A.; Okoh, A.I. Effects of probiotics on growth performance, blood parameters, and antibody stimulation in piglets. S. Afr. J. Anim. Sci. 2017, 47, 765. [Google Scholar] [CrossRef]
- Lan, R.X.; Lee, S.I.; Kim, I.H. Effects of multistrain probiotics on growth performance, nutrient digestibility, blood profiles, faecal microbial shedding, faecal score and noxious gas emission in weaning pigs. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Kim, Y.H.; Kim, I.H. Effects of two bacillus strains probiotic supplement on reproduction performance, nutrient digestibility, blood profile, fecal score, excreta odor contents and fecal microflora in lactation sows, and growth performance in sucking piglets. Livest. Sci. 2021, 244, 104293. [Google Scholar] [CrossRef]
- Ishaq, S.L.; Kim, C.J.; Reis, D.; Wright, A.-D.G. Fibrolytic Bacteria Isolated from the Rumen of North American Moose (Alces alces) and Their Use as a Probiotic in Neonatal Lambs. PLoS ONE 2015, 10, e0144804. [Google Scholar] [CrossRef]
- Salvedia, C.; Supangco, E.; Vega, R.; Elegado, F.; Rayos, A. Effect of Probiotic Feeding on Milk Yield and Components of Crossbred Dairy Goats. Philipp. J. Vet. Anim. Sci. 2015, 41, 21–30. [Google Scholar]
- Maake, T.W.; Adeleke, M.; Aiyegoro, O.A. Effect of lactic acid bacteria administered as feed supplement on the weight gain and ruminal pH in two South African goat breeds. Trans. R. Soc. S. Afr. 2021, 76, 35–40. [Google Scholar] [CrossRef]
- Aalaei, M.; Khatibjoo, A.; Zaghari, M.; Taherpour, K.; Gharaei, M.A.; Soltani, M. Comparison of single- and multistrain probiotics effects on broiler breeder performance, egg production, egg quality and hatchability. Br. Poult. Sci. 2018, 59, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Shreedhar, J.N.; Patil, M.; Kumar, P. Effect of Probiotics Supplementation on Milk Yield and Its Composition in Lactating Holstein Fresien and Deoni Cross Bred Cows. J. Med. Bioeng. 2016, 5, 19–23. [Google Scholar] [CrossRef][Green Version]
- Yoo, J.; Kim, S. Probiotics and Prebiotics: Present Status and Future Perspectives on Metabolic Disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Yan, W.; Ma, Y.; Fang, J. The impact of probiotics on gut health via alternation of immune status of monogastric animals. Anim. Nutr. 2021, 7, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, A.M.; Ibrahim, N.S.; Shehata, A.M.; Mohamed, N.G.; Abdel-Moneim, A.-M.E. Impact of multistrain probiotic, citric acid, garlic powder or their combinations on performance, ileal histomorphometry, microbial enumeration and humoral immunity of broiler chickens. Trop. Anim. Health Prod. 2021, 53, 115. [Google Scholar] [CrossRef] [PubMed]
- Fenton, H.; McManamon, R.; Howerth, E.W. Anseriformes, Ciconiiformes, Charadriiformes, and Gruiformes. In Pathology of Wildlife and Zoo Animals; Elsevier: Amsterdam, The Netherlands, 2018; pp. 697–721. [Google Scholar] [CrossRef]
- Mbuthia, P.G.; Njagi, L.W.; Nyaga, P.N.; Bebora, L.C.; Minga, U.; Kamundia, J.; Olsen, J.E. Pasteurella multocida in scavenging family chickens and ducks: Carrier status, age susceptibility and transmission between species. Avian Pathol. 2008, 37, 51–57. [Google Scholar] [CrossRef]
- Rajabi, S.; Darban, D.; Tabatabaei, R.R.; Hosseini, F. Antimicrobial effect of spore-forming probiotics Bacillus laterosporus and Bacillus megaterium against Listeria monocytogenes. Arch. Microbiol. 2020, 202, 2791–2797. [Google Scholar] [CrossRef]
- Rahimi, S.; Kathariou, S.; Fletcher, O.; Grimes, J.L. Effect of a direct-fed microbial and prebiotic on performance and intestinal histomorophology of turkey poults challenged with Salmonella and Campylobacter. Poult. Sci. 2019, 98, 6572–6578. [Google Scholar] [CrossRef]
- Smialek, M.; Burchardt, S.; Koncicki, A. The influence of probiotic supplementation in broiler chickens on population and carcass contamination with Campylobacter spp.—Field study. Res. Vet. Sci. 2018, 118, 312–316. [Google Scholar] [CrossRef]
- Olnood, C.G.; Beski, S.S.M.; Choct, M.; Iji, P.A. Use of Lactobacillus johnsonii in broilers challenged with Salmonella sofia. Anim. Nutr. 2015, 1, 203–212. [Google Scholar] [CrossRef]
- Grosu-Tudor, S.-S.; Stancu, M.-M.; Pelinescu, D.; Zamfir, M. Characterization of some bacteriocins produced by lactic acid bacteria isolated from fermented foods. World J. Microbiol. Biotechnol. 2014, 30, 2459–2469. [Google Scholar] [CrossRef]
- Reuben, R.C.; Sarkar, S.L.; Ibnat, H.; Setu, M.A.A.; Roy, P.C.; Jahid, I.K. Novel multistrain probiotics reduces Pasteurella multocida induced fowl cholera mortality in broilers. Sci. Rep. 2021, 11, 8885. [Google Scholar] [CrossRef]
- Kazemi, S.A.; Ahmadi, H.; Torshizi, M.A.K. Evaluating two multistrain probiotics on growth performance, intestinal morphology, lipid oxidation and ileal microflora in chickens. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1399–1407. [Google Scholar] [CrossRef]
- Aalaei, M.; Khatibjoo, A.; Zaghari, M.; Taherpou, K.; Akbari-Gharaei, M.; Soltani, M. Effect of single- and multistrain probiotics on broiler breeder performance, immunity and intestinal toll-like receptors expression. J. Appl. Anim. Res. 2019, 47, 236–242. [Google Scholar] [CrossRef]
- Dobrowolski, P.; Tomaszewska, E.; Klebaniuk, R.; Tomczyk-Warunek, A.; Szymańczyk, S.; Donaldson, J.; Świetlicka, I.; Mielnik-Błaszczak, M.; Kuc, D.; Muszyński, S. Structural changes in the small intestine of female turkeys receiving a probiotic preparation are dose and region dependent. Animal 2019, 13, 2773–2781. [Google Scholar] [CrossRef] [PubMed]
- Abou-Kassem, D.E.; Elsadek, M.F.; Abdel-Moneim, A.E.; Mahgoub, S.A.; Elaraby, G.M.; Taha, A.E.; Elshafie, M.M.; Alkhawtani, D.M.; Abd El-Hack, M.E.; Ashour, E.A. Growth, carcass characteristics, meat quality, and microbial aspects of growing quail fed diets enriched with two different types of probiotics (Bacillus toyonensis and Bifidobacterium bifidum). Poult. Sci. 2021, 100, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Qadis, A.Q.; Kim, Y.-H.; Ikuta, K.; Ichijo, T.; Sato, S. Effects of a bacterial probiotic on ruminal pH and volatile fatty acids during subacute ruminal acidosis (SARA) in cattle. J. Vet. Med. Sci. 2016, 78, 1595–1600. [Google Scholar] [CrossRef][Green Version]
- Mostafa, T.H.; Elsayed, F.A.; Ahmed, M.A.; Elkholany, M.A. Effect of using some Feed additives (TW-Probiotics) in Dairy Cow rations on Production and Reproductive Performance. Egypt. J. Anim. Prod. 2014, 51, 1–11. [Google Scholar]
- El-Garhi, M.S.; Soltan, M.A.; Ahmed, H.A.; Mervat, A.A.L.; Galal, M.; El-Bordeny, N.E. Assessment Impact of Using Locally Produced Probiotic Bacteria on the Productive And Reproductive Performance of Holstein Dairy Cows. Assiut Vet. Med. J. 2019, 65, 39–50. [Google Scholar] [CrossRef]
- Xu, H.; Huang, W.; Hou, Q.; Kwok, L.Y.; Sun, Z.; Ma, H.; Zhao, F.; Lee, Y.K.; Zhang, H. The effects of probiotics administration on the milk production, milk components and fecal bacteria microbiota of dairy cows. Sci. Bull. 2017, 62, 767–774. [Google Scholar] [CrossRef]
- Direkvandi, E.; Mohammadabadi, T.; Salem, A.Z.M. Oral administration of lactate producing bacteria alone or combined with Saccharomyces cerevisiae and Megasphaera elsdenii on performance of fattening lambs. J. Appl. Anim. Res. 2020, 48, 235–243. [Google Scholar] [CrossRef]
- Qorbanpour, M.; Fahim, T.; Javandel, F.; Nosrati, M.; Paz, E.; Seidavi, A.; Ragni, M.; Laudadio, V.; Tufarelli, V. Effect of Dietary Ginger (Zingiber officinale Roscoe) and Multi-Strain Probiotic on Growth and Carcass Traits, Blood Biochemistry, Immune Responses and Intestinal Microflora in Broiler Chickens. Animals 2018, 8, 117. [Google Scholar] [CrossRef]
- Talebi, A.; Amirzadeh, B.; Mokhtari, B.; Gahri, H. Effects of a multistrain probiotic (PrimaLac) on performance and antibody responses to Newcastle disease virus and infectious bursal disease virus vaccination in broiler chickens. Avian Pathol. 2008, 37, 509–512. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Kwiecień, M.; Dobrowolski, P.; Klebaniuk, R.; Muszyński, S.; Olcha, M.; Blicharski, T.; Grela, E.R. Dose-dependent effects of probiotic supplementation on bone characteristics and mineralisation in meat-type female turkeys. Anim. Prod. Sci. 2018, 58, 507. [Google Scholar] [CrossRef]
- Knap, P.W.; Wang, L. Pig breeding for improved feed efficiency. In Feed Efficiency in Swine; Patience, J.F., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Willing, B.P.; Malik, G.; van Kessel, A.G. Nutrition and Gut Health in Swine. In Sustainable Swine Nutrition; Chiba, L.I., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2012; pp. 197–213. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, N.; Zhou, M.; Tu, Y.; Deng, K.; Diao, Q. Effects of dietary probiotics on growth performance, faecal microbiota and serum profiles in weaned piglets. Anim. Prod. Sci. 2014, 54, 616. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Nyachoti, C.M.; Kim, I.H. Evaluation of effect of probiotics mixture supplementation on growth performance, nutrient digestibility, faecal bacterial enumeration, and noxious gas emission in weaning pigs. Ital. J. Anim. Sci. 2019, 18, 466–473. [Google Scholar] [CrossRef]
- Zhao, P.Y.; Kim, I.H. Effect of direct-fed microbial on growth performance, nutrient digestibility, fecal noxious gas emission, fecal microbial flora and diarrhea score in weanling pigs. Anim. Feed Sci. Technol. 2015, 200, 86–92. [Google Scholar] [CrossRef]
- Kong, Q.; He, G.-Q.; Jia, J.-L.; Zhu, Q.-L.; Ruan, H. Oral Administration of Clostridium butyricum for Modulating Gastrointestinal Microflora in Mice. Curr. Microbiol. 2011, 62, 512–517. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Kim, I.H. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult. Sci. 2014, 93, 364–370. [Google Scholar] [CrossRef]
- Agazzi, A. The Beneficial Role of Probiotics in Monogastric Animal Nutrition and Health. J. Dairy Vet. Anim. Res. 2015, 2. [Google Scholar] [CrossRef]
- Hayakawa, T.; Masuda, T.; Kurosawa, D.; Tsukahara, T. Dietary administration of probiotics to sows and/or their neonates improves the reproductive performance, incidence of post-weaning diarrhea and histopathological parameters in the intestine of weaned piglets: Beneficial Effects of Probiotics in Pigs. Anim. Sci. J. 2016, 87, 1501–1510. [Google Scholar] [CrossRef]
- Liu, W.C.; Ye, M.; Liao, J.H.; Zhao, Z.H.; Kim, I.H.; An, L.L. Application of Complex Probiotics in Swine Nutrition—A Review. Ann. Anim. Sci. 2018, 18, 335–350. [Google Scholar] [CrossRef]
- Bohmer, B.M.; Kramer, W.; Roth-Maier, D.A. Dietary probiotic supplementation and resulting effects on performance, health status, and microbial characteristics of primiparous sows. J. Anim. Physiol. Anim. Nutr. 2006, 90, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulos, C.; Georgoulakis, I.E.; Tzivara, A.; Kritas, S.K.; Siochu, A.; Kyriakis, S.C. Field evaluation of the efficacy of a probiotic containing Bacillus licheniformis and Bacillus subtilis spores, on the health status and performance of sows and their litters. J. Anim. Physiol. Anim. Nutr. 2004, 88, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.M.; Ghule, S.; Muthukumar, S.; Banik, A.; Maji, C. Effects of Dietary Supplementation of a Single-and a Multi-Strain Probiotic on Growth Performance and Intestinal Histomorphology of Commercial Broiler Chickens. Int. J. Poult. Sci. 2020, 19, 363–371. [Google Scholar] [CrossRef]
- Ramlucken, U.; Ramchuran, S.O.; Moonsamy, G.; Lalloo, R.; Thantsha, M.S.; van Rensburg, C.J. A novel Bacillus based multistrain probiotic improves growth performance and intestinal properties of Clostridium perfringens challenged broilers. Poult. Sci. 2020, 99, 331–341. [Google Scholar] [CrossRef]
- Biswas, A.; Dev, K.; Tyagi, P.K.; Mandal, A. The effect of multistrain probiotics as feed additives on performance, immunity, expression of nutrient transporter genes and gut morphometry in broiler chickens. Anim. Biosci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.H.; Lee, J.; Kong, C. Effects of Multi Strain Probiotics on Egg Production and Quality in Laying Hens Fed Diets Containing Food Waste Product. Int. J. Poult. Sci. 2014, 14, 19–22. [Google Scholar] [CrossRef]


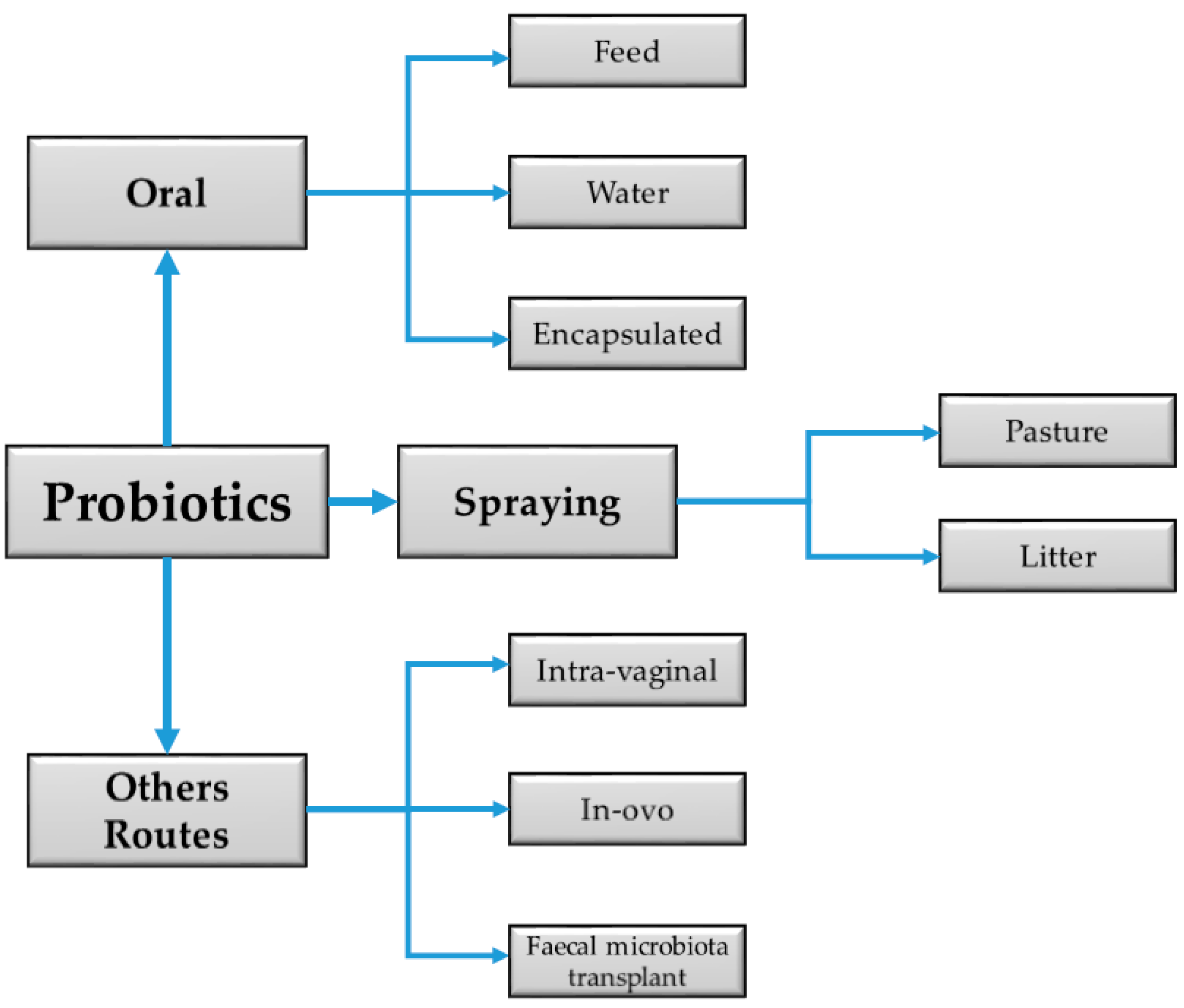{kind=link}
| Multistrain | Cell Count | Mode of Administration/Dose | Host | Duration | Effect | No Effect | Ref. |
|---|---|---|---|---|---|---|---|
| Bacillus foraminis, B. firmus B. licheniformis, Staphylococcus saprophyticus bovis | 107 CFU/g | Oral inoculant using a syringe (1 mL/day at 1–2 weeks, 2 mL/day at 3–9 weeks) | Neonate lamb | 9 weeks |
| No effect on BWG and wool quality | [76] |
| P. acidilactici 3G3 L. plantarum BS S. cerevisiae 2030 | 5 × 109 CFU/mL | Orally using a syringe (6 mL) | Dairy goats | 9 weeks |
| No effect on total milk yield, glucose, hemoglobin, and RBC count | [77] |
| E. faecalis L. rhamnous | 2 × 109 CFU/mL | Orally using dosing gun (5 mL) | South African goats | 30 days |
| No effect on feed intake | [78] |
| L. acidophilus L. casei B. thermophilum E. faecium | 107 CFU/g | (Orally) mixed with concentrate | Lactating Ewes | 8 weeks |
| Rumen conversion pathway of Fatty acid was not altered | [79] |
| L. acidophilus L. plantarum B. bifidum, B. subtilis, A. oryzae | 1 × 108 9.8 × 107 2 × 106 CFU/g | Orally (3 g or 20 g/cow/day mixed with diet) | Pre-partum dairy cow | 6 months |
| No effect on BW, birth weight of calves, blood biochemical concentrations | [80] |
| (Locally produced probiotic bacteria) containing: L. farraginis L. reuteri L. rhamnosus | 108 CFU/g DM | Orally (mixed with diet) | Pre-partum dairy cows | 3 months |
| No effect on milk lactose, solid non-fat, and ash | [81] |
| L. casei Zhang L. plantarum P-8 | 1.3 × 109 (50 g/head/day) | Orally (mixed with basal diet) | Lactating primiparous dairy cows | 4 weeks |
| No effect on milk fat, protein, and lactose | [82] |
| L. acidophilus S. cerevisiae E. faecium A. oryza B. subtilis | 50 mL/day | Orally (mixed with endotoxin-free water) | Dairy cows | 60 days |
| No effect on BW, PCV, and total protein concentration in plasma | [83] |
| L. fermentum L. plantarum M. elsdenii S. cerevisiae | 4.5 × 108 4.5 × 108 4.5 × 108 1.4 × 1010 | Orally (dosing of 50 mL microbial suspension) | Fattening lamb | 63 days |
| No effect on feed intake and blood metabolite | [84] |
| Multistrain | Cell Count | Mode of Administration/Dose | Host | Duration | Effect | No Effect | Ref. |
|---|---|---|---|---|---|---|---|
| L. acidophilus L. casei E. faecium B. thermophilum | 1 × 108 CFU/g | Orally 1–2 weeks; 0.9 3–4 weeks; 0.454 5–6 weeks; 0.225 g/kg Lyophilized mixture added to the diet | Male broiler chicks | 42 days |
| No effect on growth, carcass parameter, and blood biochemistry | [99] |
| L. acidophilus L. casei E. faecium B. bifidium | n.s | Orally (via non-chlorinated water) | Broiler chickens | 42 days |
| No effect on antibody titer | [100] |
| L. salivarius L. reuteri L. crispatus L. johnsonii | 1 × 105 1 × 106 1 × 107 CFU/egg | In-ovo (100 μL/egg injected on 18th embryonic day) | Broiler chickens | 1 day |
| No effect on T-cell in the spleen | [101] |
| B. subtilis CPB 011 B. subtilis CPB 029 B. subtilis HP 1.6 B. subtilis D 014 B. velezensis CBP 020 B. velezensis CPB 035 | 1 × 109 CFU/g | Orally (100 g/ton mixed with feed) | C. perfringens challenged male broiler chicken | 35 days |
| n.s | [102] |
| L. acidophilus B. subtilis DSM 17299 C. butyricum. | 2 × 105 CFU/kg | Orally (mixed with diet) | Broiler chickens | 5 weeks |
| No effect on IgG, lymphocyte, RBC, and WBC. | [103] |
| L. acidophilus L. casei B. thermophillum E. faecium | 2.5 × 107 CFU/g | Orally (0.1 g/kg supplemented in basal diet) | Broiler breeder (51 weeks old) | 10 weeks |
| No effect on hatchability, egg quality, mortality, fertility, BW, GIT function, or nutrient digestibility, and Lactobacillus spp. population | [66] |
| A. oryzae B. subtilis S. cerevisiae L. plantarum Rhodopseudomonas capsulate | 1 × 109 1 × 109 1 × 109 1 × 109 1 × 107 CFU/g | Orally (0.1, 0.2 and 0.4% supplemented in basal diet) | Laying hens (40 weeks old) | 3 weeks |
| No effect on yolk color and hen productivity | [104] |
| Bacillus toyonensis B. bifidum | 5 × 108 6 × 108 CFU/mL | Orally (0.5–1 mL/kg added to basal diet) | Japanese quail | 42 days |
| n.s | [105] |
| L. casei L. acidophilus Bifidobacterium | >5 × 109 CFU/g | Orally (1%; 10 mL/L of distilled drinking water) | Broiler chickens | 42 days |
| n.s | [106] |
| L. casei L. lactis L. plantarum Carnobacterium divergens S. cerevisiae | 107, 108, 109 CFU/g * (3 different doses of the mixture) | Orally (500 g/1000 kg of feed) | Meat-type female turkey | 16 weeks |
| No effect on body weight, bone mass, and bone mineral concentration | [107] |
| Multistrain | Cell Count | Mode of Administration/Dose | Host | Duration | Effect | No Effect | Ref. |
|---|---|---|---|---|---|---|---|
| L. acidophilus B. subtilis S. cerevisiae | 1 × 107 1 × 107 1 × 107 CFU/g | Orally (0.1% and 0.2% mixed with basal diet) | Finishing pigs | 10 weeks |
| No effect on meat quality parameters | [117] |
| Product A: L. plantarum L21 L. plantarum L80 L. paraplantarum L103 Product B: B. subtilis L. acidophilus S. cerevisiae | 1 × 109 1 × 109 1 × 109 1 × 1012 1.5 × 107 1 × 109 CFU/mL | Oral gavage (0.25 g/day) | Weaned pigs | 28 days |
| n.s | [120] |
| B.coagulans B. licheniformis B. subtilis C. butyricum | 1 × 109 5 × 108 1 × 109 1 × 108 CFU/g | Orally (0.1 or 0.2 g/kg mixed with basal diet) | Growing-finishing pigs | 16 weeks |
| No effect on average daily feed intake and meat color | [121] |
| L. amylovorus L. reuteri LAB 26 L. reuteri LAB 49 L. johnsonii L. salivarius L. mucosae | 1.7 × 1019 CFU/mL | Orally (1 mL mixed with PBS and 13% glycerol, aliquots added to feed) | Piglets | 3 weeks |
| No effect on the population of lactobacilli and bacteria in the large intestine digesta and growth enhancement | [122] |
| B. subtilitis B. licheniformis | 1 × 109 CFU/g | Orally (0.1 and 0.2% inoculated into limestone and maltodextrin as carriers) | Lactating sow and their suckling piglets | 28 days |
| No effect on reproductive performance, H2S concentration, and fecal score in sows | [123] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambo, M.T.; Chang, X.; Liu, D. The Recent Trend in the Use of Multistrain Probiotics in Livestock Production: An Overview. Animals 2021, 11, 2805. https://doi.org/10.3390/ani11102805
Lambo MT, Chang X, Liu D. The Recent Trend in the Use of Multistrain Probiotics in Livestock Production: An Overview. Animals. 2021; 11(10):2805. https://doi.org/10.3390/ani11102805
Chicago/Turabian StyleLambo, Modinat Tolani, Xiaofeng Chang, and Dasen Liu. 2021. "The Recent Trend in the Use of Multistrain Probiotics in Livestock Production: An Overview" Animals 11, no. 10: 2805. https://doi.org/10.3390/ani11102805
APA StyleLambo, M. T., Chang, X., & Liu, D. (2021). The Recent Trend in the Use of Multistrain Probiotics in Livestock Production: An Overview. Animals, 11(10), 2805. https://doi.org/10.3390/ani11102805