Functional Analysis of Haplotypes in Bovine PSAP Gene and Their Relationship with Beef Cattle Production Traits

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples and Data Collection
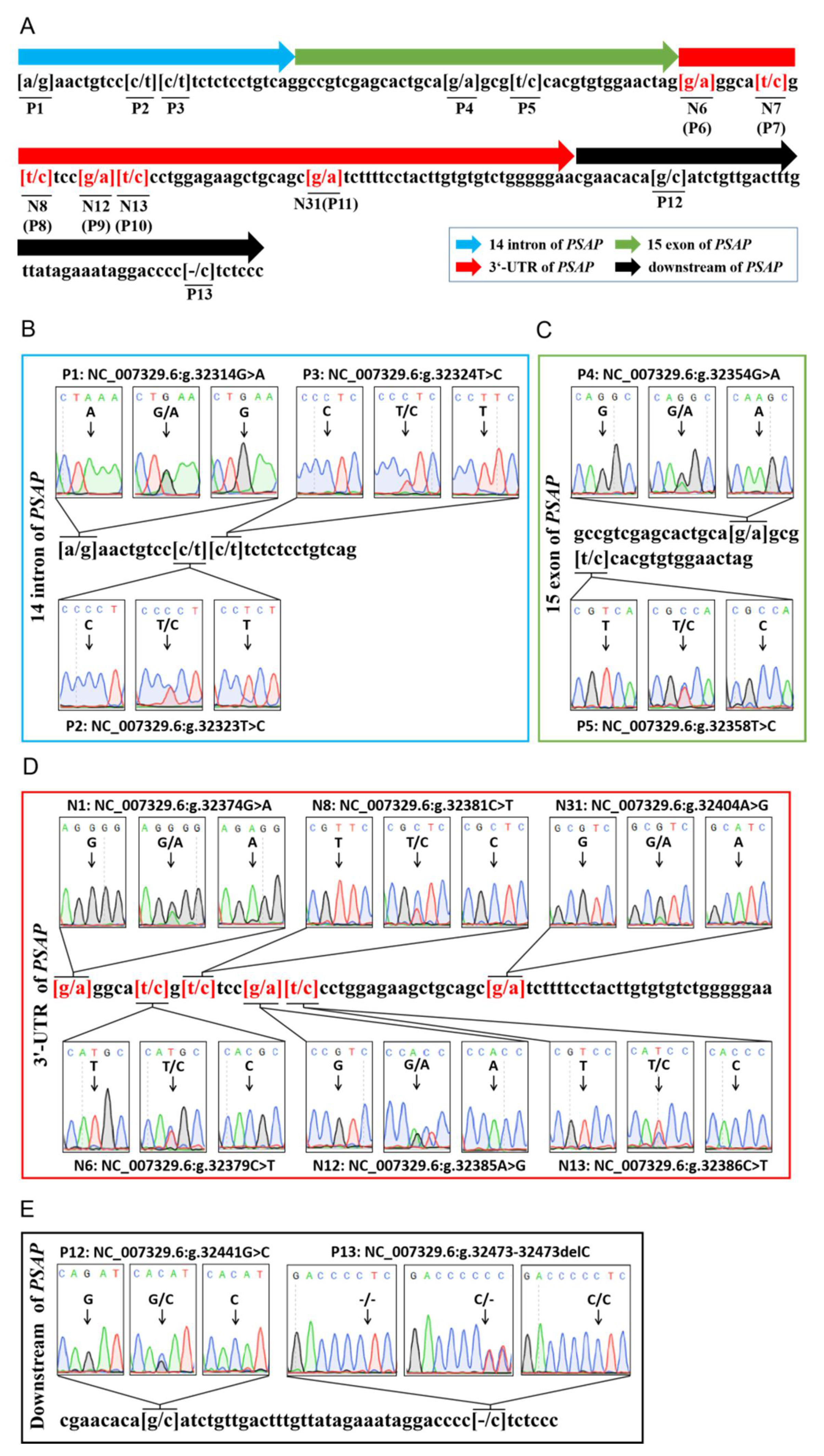
2.2. Primer Design and Variation Genotyping
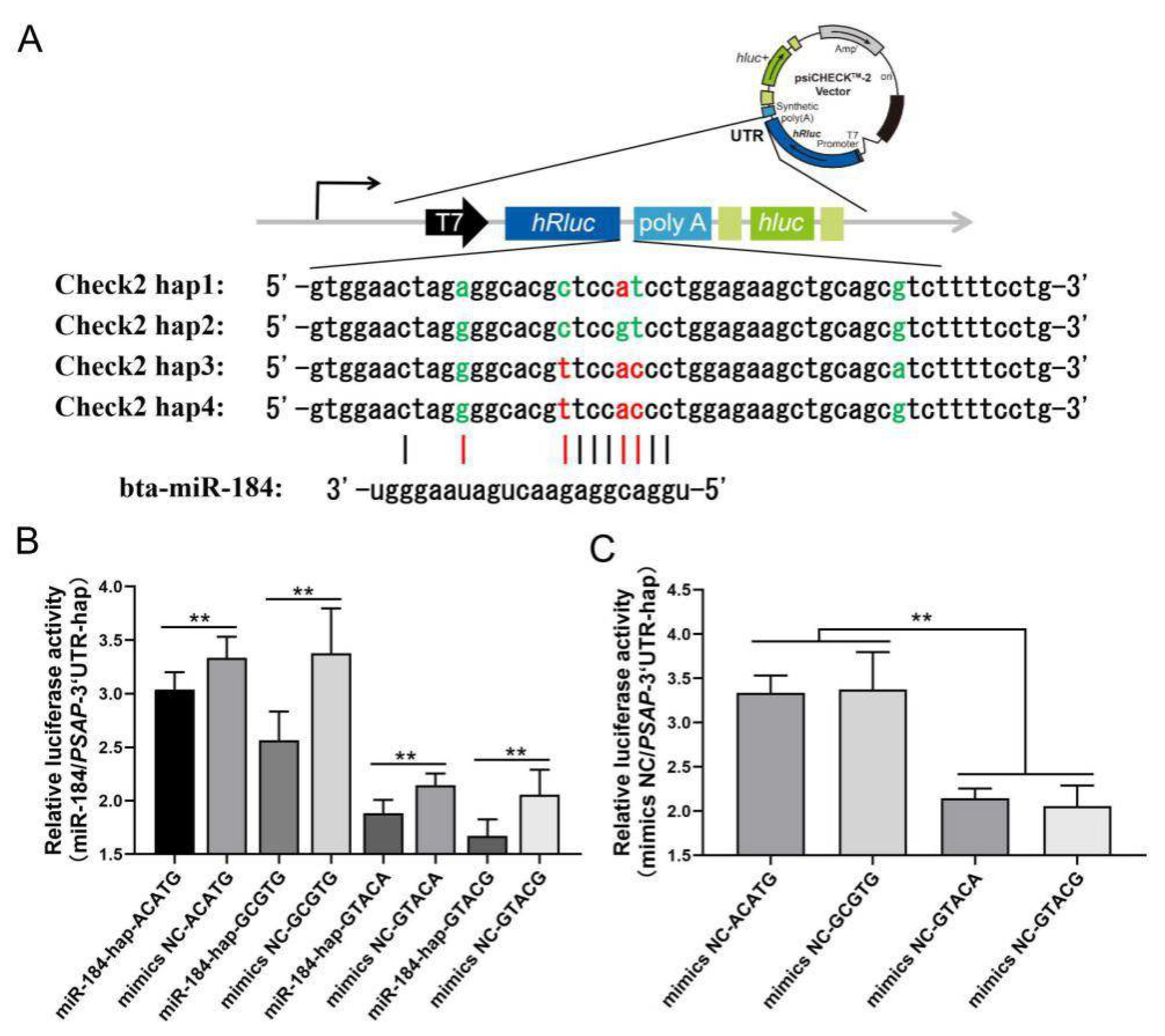
2.3. Dual-Luciferase Reporter Assay
2.4. Statistical Analysis
3. Results
3.1. Thirteen Variations Were Identified in Bovine PSAP
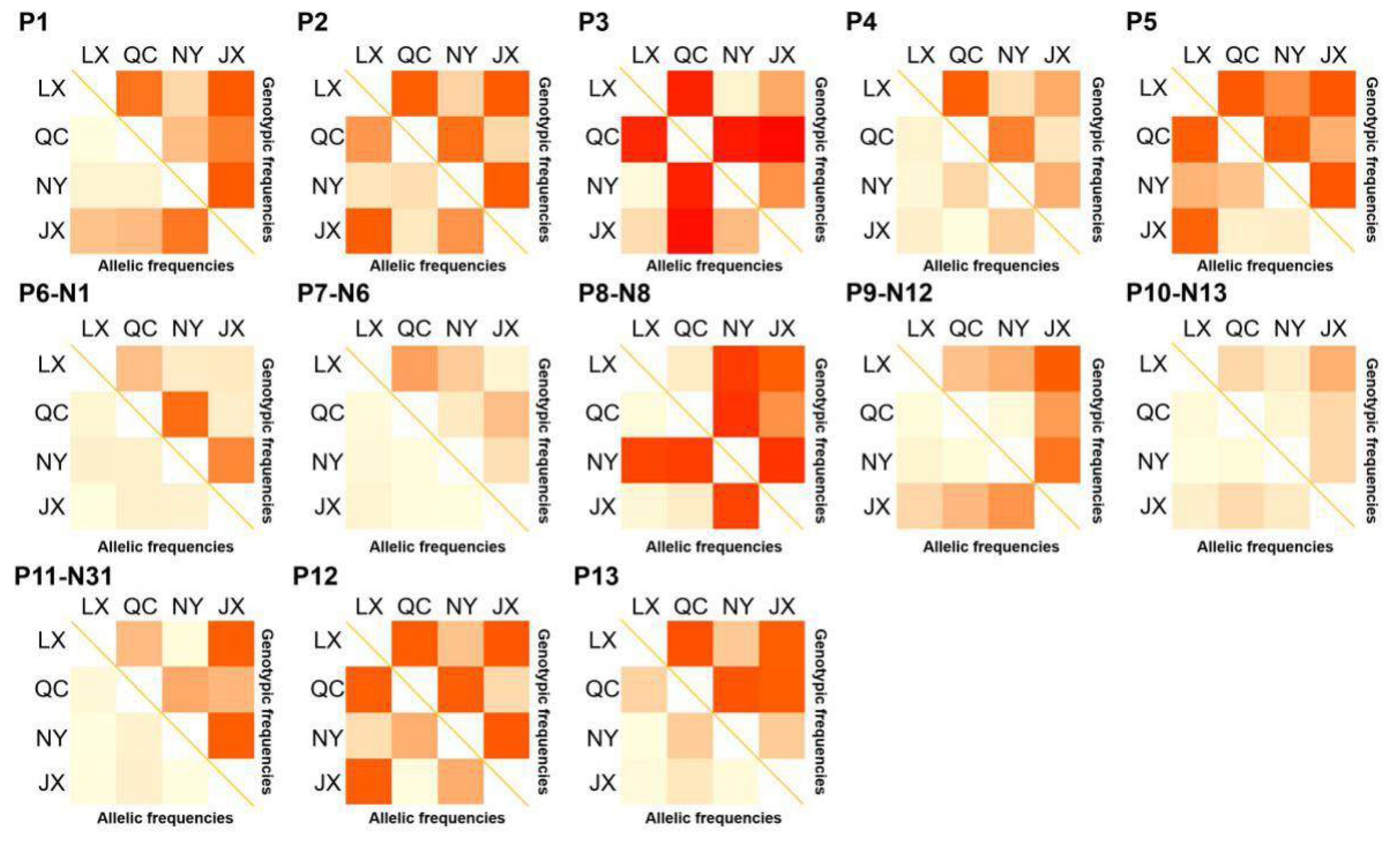
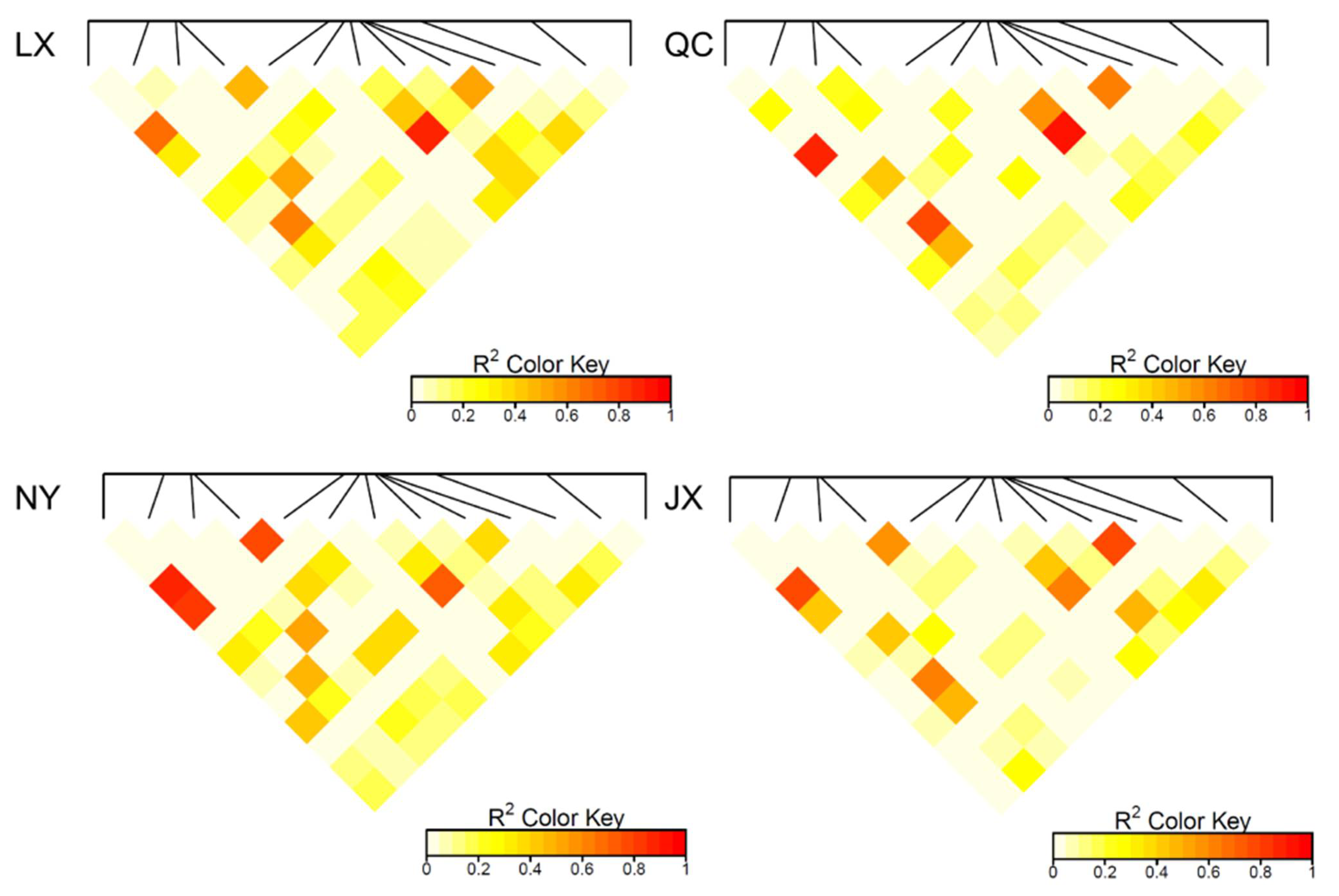
3.2. Population Parameters of 13 Variations in Four Cattle Breeds
3.3. Association between 13 Variations and Morphological Traits
3.4. Association of Four Haplotypes of PSAP 3’ UTR and Morphological Traits in NY Cattle
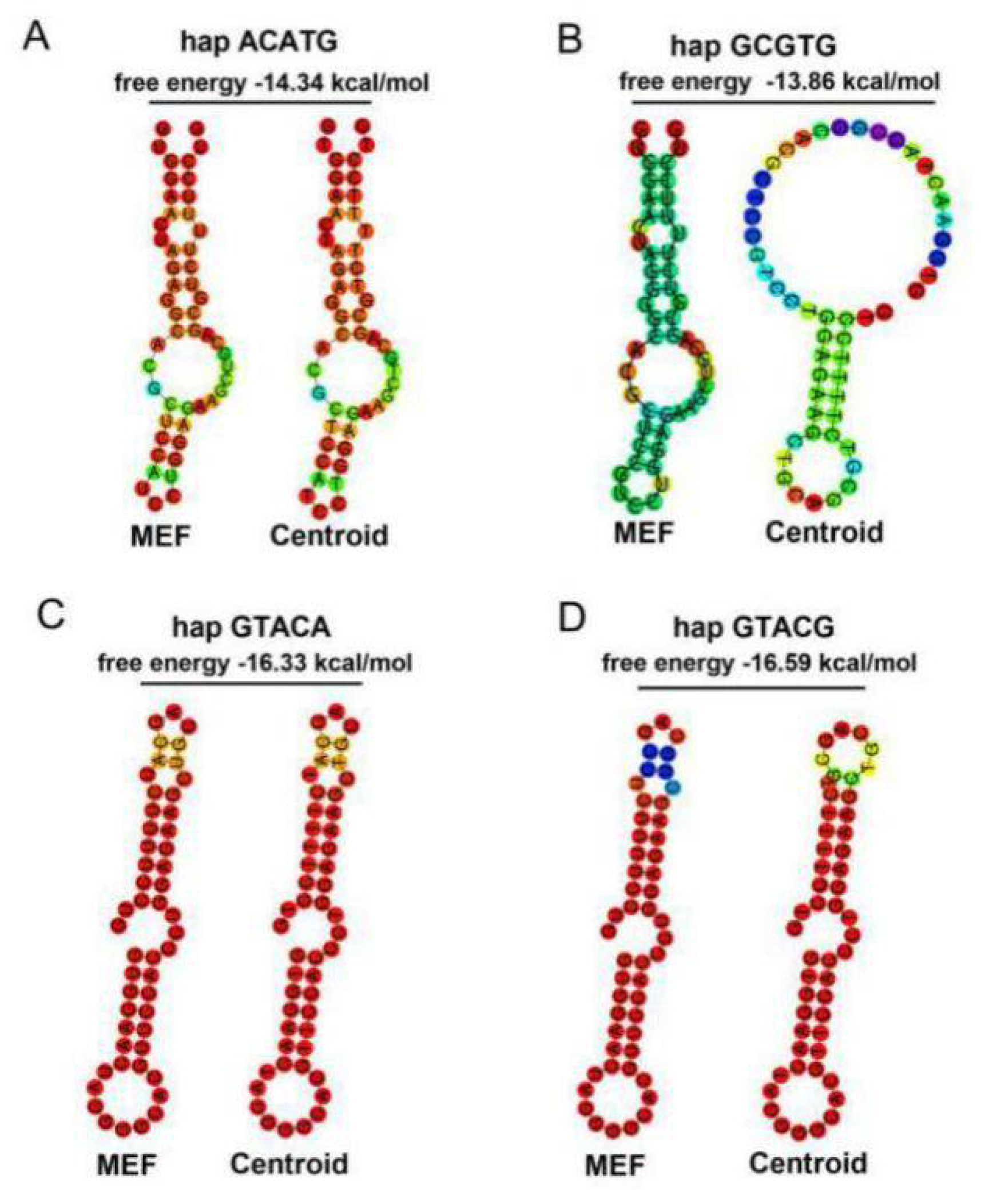
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, J.; Guo, Y.; Xu, J.; Li, H.; Fuller, A.; Tait, R.G., Jr.; Wu, X.L.; Bauck, S. Comparing SNP panels and statistical methods for estimating genomic breed composition of individual animals in ten cattle breeds. BMC Genom. 2018, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, Y.; Cho, E.; Cho, K.; Sa, S.; Kim, Y.; Choi, J.; Kim, J.; Hong, J.; Choi, T. Genomic Analysis Using Bayesian Methods under Different Genotyping Platforms in Korean Duroc Pigs. Animals 2020, 10, 752. [Google Scholar] [CrossRef] [PubMed]
- Akanno, E.C.; Abo-Ismail, M.K.; Chen, L.; Crowley, J.J.; Wang, Z.; Li, C.; Basarab, J.A.; MacNeil, M.D.; Plastow, G.S. Modeling heterotic effects in beef cattle using genome-wide SNP-marker genotypes. J. Anim. Sci. 2018, 96, 830–845. [Google Scholar] [CrossRef] [PubMed]
- Steri, M.; Idda, M.L.; Whalen, M.B.; Orrù, V. Genetic variants in mRNA untranslated regions. Wires RNA 2018, 9, e1474. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, S.; Du, M.; Chu, H.; Wang, M.; Zhang, Z. Novel CpG-SNPs in the gastric acid secretion pathway GNAI3 and susceptibility to gastric cancer. Gene 2020, 736, 144447. [Google Scholar] [CrossRef] [PubMed]
- Pegolo, S.; Cecchinato, A.; Savoia, S.; Di Stasio, L.; Pauciullo, A.; Brugiapaglia, A.; Bittante, G.; Albera, A. Genome-wide association and pathway analysis of carcass and meat quality traits in Piemontese young bulls. Animal 2020, 14, 243–252. [Google Scholar] [CrossRef]
- Kononoff, P.J.; Defoor, P.J.; Engler, M.J.; Swingle, R.S.; Gleghorn, J.F.; James, S.T.; Marquess, F.L. Impacts of a leptin SNP on growth performance and carcass characters in finishing steers studied over time. J. Anim. Sci. 2017, 95, 194–200. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, Y.; Shi, T.; Cai, H.; Lan, X.; Zhao, X.; Plath, M.; Chen, H. Whole-genome sequencing reveals mutational landscape underlying phenotypic differences between two widespread Chinese cattle breeds. PLoS ONE 2017, 12, e0183921. [Google Scholar] [CrossRef]
- Han, B.; Yuan, Y.; Shi, L.; Li, Y.; Liu, L.; Sun, D. Identification of single nucleotide polymorphisms of PIK3R1 and DUSP1 genes and their genetic associations with milk production traits in dairy cows. J. Anim. Sci. Biotechnol. 2019, 10, 81. [Google Scholar] [CrossRef]
- Smith, J.L.; Wilson, M.L.; Nilson, S.M.; Rowan, T.N.; Oldeschulte, D.L.; Schnabel, R.D.; Decker, J.E.; Seabury, C.M. Genome-wide association and genotype by environment interactions for growth traits in U.S. Gelbvieh cattle. BMC Genom. 2019, 20, 926. [Google Scholar] [CrossRef]
- Xu, Z.Q.; He, J.; Ji, C.L.; Zhang, Y.; Nie, Q.H.; Zhang, D.X.; Zhang, X.Q. Polymorphisms in the 5’-flanking regions of the GH, PRL, and Pit-1 genes with Muscovy duck egg production. J. Anim. Sci. 2015, 93, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Si, Y.; He, F.; Wen, H.; Li, J.; Zhao, J.; Ren, Y.; Zhao, M.; Ji, L.; Huang, Z.; Zhang, M.; et al. Genetic polymorphisms and DNA methylation in exon 1 CpG-rich regions of PACAP gene and its effect on mRNA expression and growth traits in half smooth tongue sole (Cynoglossus semilaevis). Fish. Physiol. Biochem. 2016, 42, 407–421. [Google Scholar] [CrossRef]
- Cai, X.; Yang, Z.H.; Li, H.J.; Xiao, X.; Li, M.; Chang, H. A Human-Specific Schizophrenia Risk Tandem Repeat Affects Alternative Splicing of a Human-Unique Isoform AS3MTd2d3 and Mushroom Dendritic Spine Density. Schizophr. Bull 2020, sbaa098. [Google Scholar] [CrossRef] [PubMed]
- Kolnikova, M.; Jungova, P.; Skopkova, M.; Foltan, T.; Gasperikova, D.; Mattosova, S.; Chandoga, J. Late Infantile Metachromatic Leukodystrophy Due to Novel Pathogenic Variants in the PSAP Gene. J. Mol. Neurosci. 2019, 67, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Khan, M.; Nabeka, H.; Shimokawa, T.; Yamamiya, K.; Matsuda, S. Age- and sex-associated changes in prosaposin and its receptors in the lacrimal glands of rats. Histol. Histopathol. 2020, 35, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wu, M.; Wang, S.; Yu, X.; Sun, X. Identification of a novel 24 bp insertion–deletion (indel) of the androgen receptor gene and its association with growth traits in four indigenous cattle breeds. Arch. Anim. Breed. 2018, 61, 71–78. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, J.; Hou, D.; Luo, P.; Gao, H.; Ma, Y.; Chen, Y.S.; Li, L.; Zou, D.; Zhang, H.; et al. Prosaposin is a biomarker of mesenchymal glioblastoma and regulates mesenchymal transition through the TGF-β1/Smad signaling pathway. J. Pathol. 2019, 249, 26–38. [Google Scholar] [CrossRef]
- Guo, P.; Zhao, Z.; Yan, S.; Li, J.; Yang, R. PSAP gene variants and haplotypes reveal significant effects on carcass and meat quality traits in chinese simmental-cross cattle. Arch. Anim. Breed. 2016, 59, 461–468. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, H.; Wu, M.; Yi, X.; Chen, P.; Liu, S.; Pan, Y.; Li, Q.; Tang, X.; Sun, X. Exploring of InDel in bovine PSAP gene and their association with growth traits in different development stages. Anim. Biotechnol. 2020, 1–12. [Google Scholar] [CrossRef]
- Zhao, H.; He, S.; Zhu, Y.; Cao, X.; Luo, R.; Cai, Y.; Xu, H.; Sun, X. A novel 29 bp insertion/deletion (indel) variant of the lhx3 gene and its influence on growth traits in four sheep breeds of various fecundity. Arch. Anim. Breed. 2017, 60, 79–85. [Google Scholar] [CrossRef]
- Kukučková, V.; Moravčíková, N.; Curik, I.; Simčič, M.; Mészáros, G.; Kasarda, R. Genetic diversity of local cattle. Acta Biochim. Pol. 2018, 65, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Cai, Y.; Chen, Q.; Li, R.; Wang, K.; Huang, Y.; Hu, S.; Huang, S.; Zhang, H.; Zheng, Z.; et al. Whole-genome resequencing reveals world-wide ancestry and adaptive introgression events of domesticated cattle in East Asia. Nat. Commun. 2018, 9, 2337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Gao, X.; Zhang, Y.; Zhao, Y.; Zhang, J.; Jia, Y.; Zhu, B.; Xu, L.; Zhang, L.; Gao, H.; et al. Genome-wide assessment of genetic diversity and population structure insights into admixture and introgression in Chinese indigenous cattle. BMC Genet. 2018, 19, 114. [Google Scholar] [CrossRef] [PubMed]
- Camara, Y.; Moula, N.; Sow, F.; Sissokho, M.M.; Antoine-Moussiaux, N. Analysing innovations among cattle smallholders to evaluate the adequacy of breeding programs. Animal 2019, 13, 417–426. [Google Scholar] [CrossRef]
- Tong, B.; Zhang, L.; Li, G.P. Progress in the molecular and genetic modification breeding of beef cattle in China. Hereditas 2017, 39, 984–1015. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, L.; Wang, Z.; Xu, L.; Chen, Y.; Zhang, L.; Xu, L.; Gao, X.; Gao, H.; Zhu, B.; et al. Genomic Prediction and Association Analysis with Models Including Dominance Effects for Important Traits in Chinese Simmental Beef Cattle. Animals 2019, 9, 1055. [Google Scholar] [CrossRef]
- Pirooz, H.J.; Jafari, N.; Rastegari, M.; Fathi-Roudsari, M.; Tasharrofi, N.; Shokri, G.; Tamadon, M.; Sazegar, H.; Kouhkan, F. Functional SNP in microRNA-491-5p binding site of MMP9 3’-UTR affects cancer susceptibility. J. Cell Biochem. 2018, 119, 5126–5134. [Google Scholar] [CrossRef]
- Lin, X.; Xia, W.; Ji, W.; Xie, F.F.; He, P.; Zhu, X.W.; Zhang, Y.H.; Deng, F.Y.; Lei, S.F. Genome-wide integrative analysis identified SNP-miRNA-mRNA interaction networks in peripheral blood mononuclear cells. Epigenomics 2017, 9, 1287–1298. [Google Scholar] [CrossRef]
- Jiang, Q.; Zhao, H.; Li, R.; Zhang, Y.; Liu, Y.; Wang, J.; Wang, X.; Ju, Z.; Liu, W.; Hou, M.; et al. In silico genome-wide miRNA-QTL-SNPs analyses identify a functional SNP associated with mastitis in Holsteins. BMC Genet. 2019, 20, 46. [Google Scholar] [CrossRef]
- Ceolin, L.; Romitti, M.; Siqueira, D.R.; Vaz Ferreira, C.; Oliboni Scapineli, J.; Assis-Brazil, B.; Vieira Maximiano, R.; Dias Amarante, T.; de Souza Nunes, M.C.; Weber, G.; et al. Effect of 3’UTR RET Variants on RET mRNA Secondary Structure and Disease Presentation in Medullary Thyroid Carcinoma. PLoS ONE 2016, 11, e0147840. [Google Scholar] [CrossRef]
- Walsh, I.M.; Bowman, M.A.; Soto Santarriaga, I.F.; Rodriguez, A.; Clark, P.L. Synonymous codon substitutions perturb cotranslational protein folding in vivo and impair cell fitness. Proc. Natl. Acad. Sci. USA 2020, 117, 3528–3534. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, Y.; Li, H.P.; Han, L.; Yan, X.M.; Li, H.B.; Du, W.; Zhang, J.S.; Yu, Q.L. Differential expression of mRNA-miRNAs related to intramuscular fat content in the longissimus dorsi in Xinjiang brown cattle. PLoS ONE 2018, 13, e0206757. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, M.; Rong, Y.; Liu, Z.; Yang, S.; Zhang, W.; Li, J.; Cai, Y. A Novel SNPs in Alpha-Lactalbumin Gene Effects on Lactation Traits in Chinese Holstein Dairy Cows. Animals 2019, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tang, J.; He, X.; Di, R.; Chu, M. Mutations in NLRP5 and NLRP9 are Associated with Litter Size in Small Tail Han Sheep. Animals 2020, 10, 689. [Google Scholar] [CrossRef]
- Martignani, E.; Miretti, S.; Vincenti, L.; Baratta, M. Correlation between estrogen plasma level and miRNAs in muscle of Piedmontese cattle. Domest. Anim. Endocrinol. 2019, 67, 37–41. [Google Scholar] [CrossRef]
- Abo-Ismail, M.K.; Lansink, N.; Akanno, E.; Karisa, B.K.; Crowley, J.J.; Moore, S.S.; Bork, E.; Stothard, P.; Basarab, J.A.; Plastow, G.S. Development and validation of a small SNP panel for feed efficiency in beef cattle. J. Anim. Sci. 2018, 96, 375–397. [Google Scholar] [CrossRef]
- Li, M.; Gao, Q.; Wang, M.; Liang, Y.; Sun, Y.; Chen, Z.; Zhang, H.; Karrow, N.A.; Yang, Z.; Mao, Y. Polymorphisms in Fatty Acid Desaturase 2 Gene are Associated with Milk Production Traits in Chinese Holstein Cows. Animals 2020, 10, 671. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Sampling Location | Population |
|---|---|---|
| QC (Qinchuan cattle) | Yangling, Shaanxi | 123 |
| NY (Nanyang cattle) | Nanyang, Henan | 137 |
| JX (Jiaxian red cattle) | Nanyang, Henan | 137 |
| LX (Luxi cattle) | Jining, Shandong | 104 |
| Primers | Sequence | Notes |
|---|---|---|
| PSAP-HAP1-F1 | TCGAGgtggaactagaggcacgctccatcctggagaagctgcagcgtcttttcctgGC | Vector construction |
| PSAP-HAP1-R1 | GGCCGCcaggaaaagacgctgcagcttctccaggatggagcgtgcctctagttccacC | |
| PSAP-HAP2-F2 | TCGAGgtggaactaggggcacgctccgtcctggagaagctgcagcgtcttttcctgGC | Vector construction |
| PSAP-HAP2-R2 | GGCCGCcaggaaaagacgctgcagcttctccaggacggagcgtgcccctagttccacC | |
| PSAP-HAP3-F3 | TCGAGgtggaactaggggcacgttccaccctggagaagctgcagcatcttttcctgGC | Vector construction |
| PSAP-HAP3-R3 | GGCCGCcaggaaaagatgctgcagcttctccagggtggaacgtgcccctagttccacC | |
| PSAP-HAP4-F4 | TCGAGgtggaactaggggcacgttccaccctggagaagctgcagcgtcttttcctgGC | Vector construction |
| PSAP-HAP4-R4 | GGCCGCcaggaaaagacgctgcagcttctccagggtggaacgtgcccctagttccacC | |
| PSAP-F | GGTGTCGGGTCCTCTTTCTG | Variations screening |
| PSAP-R | GCGTGTCGGCATCTGTCTAG |
| Locus | Breed | Size | Genotype Frequency | Allele Frequency | HWE | Population Parameters | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | GG | GA | AA | G | A | pValue | Ho | He | Ne | PIC | ||
| P1 | LX | 104 | 11 | 6 | 87 | 0.135 | 0.865 | p < 0.05 | 0.767 | 0.233 | 1.304 | 0.206 |
| QC | 123 | 6 | 21 | 96 | 0.134 | 0.866 | p < 0.05 | 0.768 | 0.232 | 1.303 | 0.205 | |
| NY | 137 | 8 | 12 | 117 | 0.102 | 0.898 | p < 0.05 | 0.817 | 0.183 | 1.225 | 0.167 | |
| JX | 137 | 12 | 40 | 85 | 0.234 | 0.766 | p < 0.05 | 0.642 | 0.358 | 1.558 | 0.294 | |
| N | CC | TC | TT | C | T | pValue | Ho | He | Ne | PIC | ||
| P2 | LX | 104 | 65 | 19 | 20 | 0.716 | 0.284 | p < 0.05 | 0.594 | 0.406 | 1.685 | 0.324 |
| QC | 123 | 93 | 24 | 6 | 0.854 | 0.146 | p < 0.05 | 0.750 | 0.250 | 1.333 | 0.219 | |
| NY | 137 | 99 | 17 | 21 | 0.785 | 0.215 | p < 0.05 | 0.662 | 0.338 | 1.510 | 0.281 | |
| JX | 137 | 113 | 21 | 3 | 0.901 | 0.099 | p < 0.05 | 0.822 | 0.178 | 1.216 | 0.162 | |
| N | CC | TC | TT | C | T | pValue | Ho | He | Ne | PIC | ||
| P3 | LX | 104 | 88 | 7 | 9 | 0.880 | 0.120 | p < 0.05 | 0.789 | 0.211 | 1.268 | 0.189 |
| QC | 123 | 14 | 6 | 103 | 0.138 | 0.862 | p < 0.05 | 0.762 | 0.238 | 1.313 | 0.210 | |
| NY | 137 | 111 | 13 | 13 | 0.858 | 0.142 | p < 0.05 | 0.756 | 0.244 | 1.323 | 0.214 | |
| JX | 137 | 122 | 12 | 3 | 0.934 | 0.066 | p < 0.05 | 0.877 | 0.123 | 1.140 | 0.115 | |
| N | GG | GA | AA | G | A | pValue | Ho | He | Ne | PIC | ||
| P4 | LX | 104 | 91 | 5 | 8 | 0.899 | 0.101 | p < 0.05 | 0.818 | 0.182 | 1.222 | 0.165 |
| QC | 123 | 107 | 15 | 1 | 0.930 | 0.070 | p < 0.05 | 0.871 | 0.129 | 1.148 | 0.120 | |
| NY | 137 | 113 | 13 | 11 | 0.872 | 0.128 | p < 0.05 | 0.777 | 0.223 | 1.287 | 0.198 | |
| JX | 137 | 122 | 12 | 3 | 0.934 | 0.066 | p < 0.05 | 0.877 | 0.123 | 1.140 | 0.115 | |
| N | CC | TC | TT | C | T | pValue | Ho | He | Ne | PIC | ||
| P5 | LX | 104 | 33 | 21 | 50 | 0.418 | 0.582 | p < 0.05 | 0.513 | 0.487 | 1.948 | 0.368 |
| QC | 123 | 9 | 27 | 87 | 0.183 | 0.817 | p < 0.05 | 0.701 | 0.299 | 1.426 | 0.254 | |
| NY | 137 | 30 | 18 | 89 | 0.285 | 0.715 | p < 0.05 | 0.593 | 0.407 | 1.687 | 0.324 | |
| JX | 137 | 8 | 47 | 82 | 0.230 | 0.770 | p < 0.05 | 0.646 | 0.354 | 1.548 | 0.291 | |
| N | GG | GC | CC | G | C | pValue | Ho | He | Ne | PIC | ||
| P12 | LX | 104 | 48 | 28 | 28 | 0.596 | 0.404 | p < 0.05 | 0.518 | 0.482 | 1.929 | 0.366 |
| QC | 123 | 24 | 42 | 57 | 0.366 | 0.634 | p < 0.05 | 0.536 | 0.464 | 1.866 | 0.356 | |
| NY | 137 | 54 | 30 | 53 | 0.504 | 0.496 | p < 0.05 | 0.500 | 0.500 | 2.000 | 0.375 | |
| JX | 137 | 21 | 59 | 57 | 0.369 | 0.631 | p > 0.05 | 0.535 | 0.465 | 1.871 | 0.357 | |
| N | CC | C- | -- | C | - | pValue | Ho | He | Ne | PIC | ||
| P13 | LX | 104 | 32 | 14 | 58 | 0.375 | 0.625 | p < 0.05 | 0.531 | 0.496 | 1.882 | 0.359 |
| QC | 123 | 26 | 67 | 30 | 0.484 | 0.516 | p > 0.05 | 0.501 | 0.499 | 1.998 | 0.375 | |
| NY | 137 | 36 | 31 | 70 | 0.376 | 0.624 | p < 0.05 | 0.531 | 0.469 | 1.884 | 0.359 | |
| JX | 137 | 34 | 44 | 59 | 0.409 | 0.591 | p < 0.05 | 0.517 | 0.483 | 1.936 | 0.367 | |
| Locus | Breed | Size | Genotype Frequency | Allele Frequency | HWE | Population Parameters | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | GG | GA | AA | G | A | pValue | Ho | He | Ne | PIC | ||
| P6 (N1) | LX | 104 | 91 | 8 | 5 | 0.913 | 0.087 | p < 0.05 | 0.842 | 0.158 | 1.188 | 0.146 |
| QC | 123 | 109 | 13 | 1 | 0.939 | 0.061 | p > 0.05 | 0.885 | 0.115 | 1.129 | 0.108 | |
| NY | 137 | 115 | 10 | 12 | 0.876 | 0.124 | p < 0.05 | 0.783 | 0.217 | 1.278 | 0.194 | |
| JX | 137 | 122 | 12 | 3 | 0.908 | 0.092 | p < 0.05 | 0.877 | 0.123 | 1.140 | 0.115 | |
| N | CC | TC | TT | C | T | pValue | Ho | He | Ne | PIC | ||
| P7 (N6) | LX | 104 | 101 | 1 | 2 | 0.976 | 0.024 | p < 0.05 | 0.953 | 0.046 | 1.049 | 0.046 |
| QC | 123 | 113 | 9 | 1 | 0.955 | 0.045 | p > 0.05 | 0.914 | 0.085 | 1.093 | 0.082 | |
| NY | 137 | 127 | 7 | 3 | 0.953 | 0.047 | p < 0.05 | 0.910 | 0.090 | 1.100 | 0.086 | |
| JX | 137 | 132 | 3 | 2 | 0.947 | 0.053 | p < 0.05 | 0.950 | 0.050 | 1.052 | 0.049 | |
| N | TT | TC | CC | T | C | pValue | Ho | He | Ne | PIC | ||
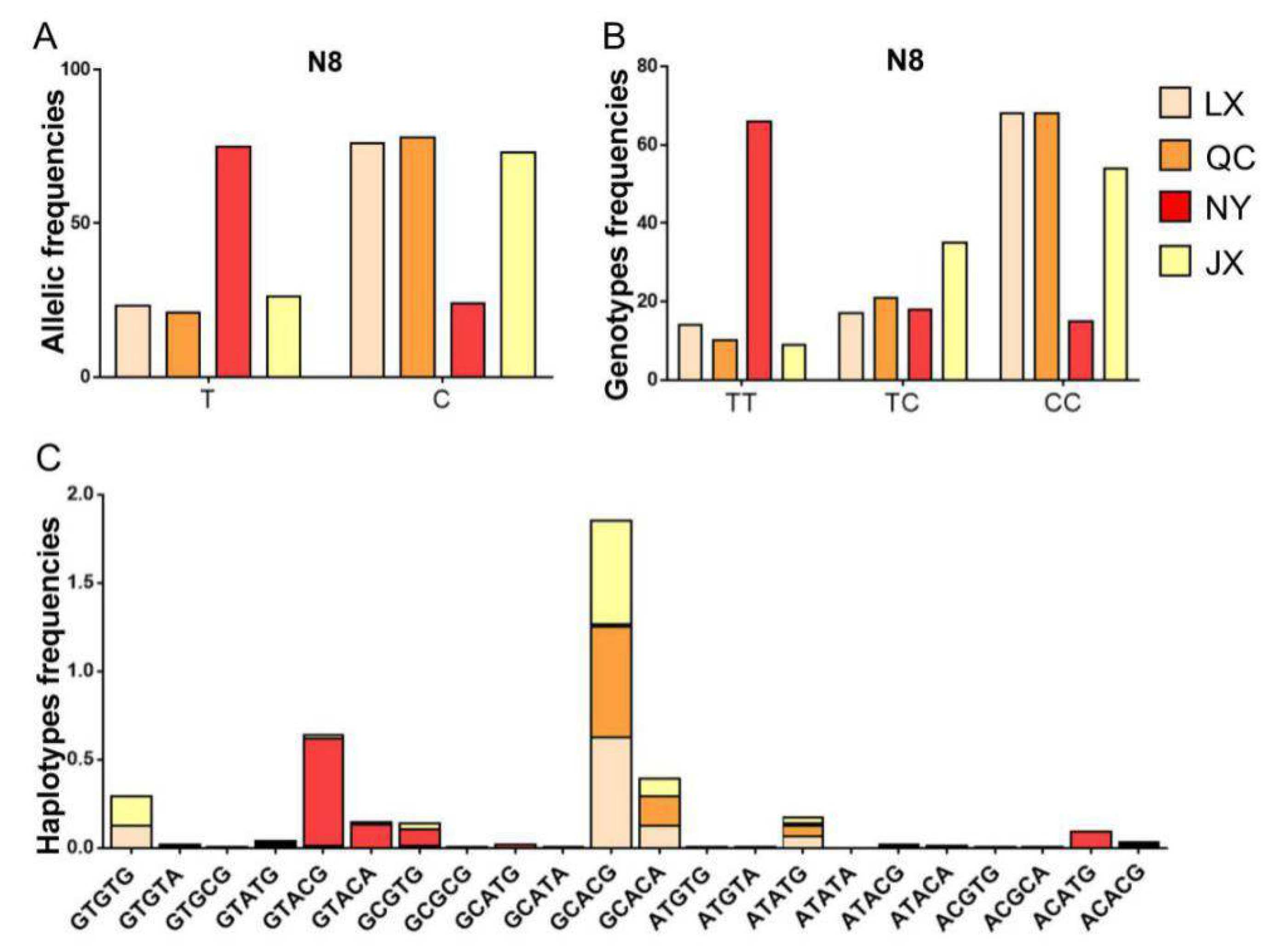
| P8 (N8) | LX | 104 | 15 | 18 | 71 | 0.231 | 0.769 | p < 0.05 | 0.645 | 0.355 | 1.550 | 0.292 |
| QC | 123 | 13 | 26 | 84 | 0.211 | 0.789 | p < 0.05 | 0.667 | 0.333 | 1.500 | 0.279 | |
| NY | 137 | 91 | 25 | 21 | 0.755 | 0.245 | p < 0.05 | 0.630 | 0.369 | 1.586 | 0.301 | |
| JX | 137 | 13 | 49 | 75 | 0.266 | 0.734 | p > 0.05 | 0.602 | 0.398 | 1.660 | 0.319 | |
| N | AA | GA | GG | A | G | pValue | Ho | He | Ne | PIC | ||
| P9 (N12) | LX | 104 | 85 | 7 | 12 | 0.851 | 0.149 | p < 0.05 | 0.746 | 0.254 | 1.340 | 0.221 |
| QC | 123 | 99 | 16 | 8 | 0.870 | 0.130 | p < 0.05 | 0.774 | 0.226 | 1.293 | 0.201 | |
| NY | 137 | 113 | 17 | 7 | 0.887 | 0.113 | p < 0.05 | 0.799 | 0.201 | 1.251 | 0.181 | |
| JX | 137 | 92 | 33 | 12 | 0.770 | 0.230 | p < 0.05 | 0.670 | 0.330 | 1.491 | 0.275 | |
| N | CC | TC | TT | C | T | pValue | Ho | He | Ne | PIC | ||
| P10 (N13) | LX | 104 | 72 | 17 | 15 | 0.774 | 0.226 | P < 0.05 | 0.650 | 0.350 | 1.538 | 0.289 |
| QC | 123 | 85 | 27 | 11 | 0.801 | 0.199 | p < 0.05 | 0.681 | 0.319 | 1.468 | 0.268 | |
| NY | 137 | 92 | 29 | 16 | 0.777 | 0.223 | p < 0.05 | 0.654 | 0.346 | 1.529 | 0.286 | |
| JX | 137 | 82 | 39 | 16 | 0.720 | 0.280 | p < 0.05 | 0.616 | 0.384 | 1.623 | 0.310 | |
| N | AA | GA | GG | A | G | pValue | Ho | He | Ne | PIC | ||
| P11 (N31) | LX | 104 | 11 | 7 | 86 | 0.139 | 0.861 | p < 0.05 | 0.760 | 0.240 | 1.316 | 0.211 |
| QC | 123 | 11 | 19 | 93 | 0.167 | 0.833 | p < 0.05 | 0.722 | 0.278 | 1.385 | 0.239 | |
| NY | 137 | 13 | 9 | 115 | 0.128 | 0.872 | p < 0.05 | 0.777 | 0.223 | 1.287 | 0.198 | |
| JX | 137 | 4 | 27 | 106 | 0.124 | 0.876 | p > 0.05 | 0.777 | 0.223 | 1.287 | 0.198 | |
| Locus | Breed | Morphological Trait | Observed Genotypes (Mean a ± SE) | |||
|---|---|---|---|---|---|---|
| GG | GA | AA | p Value | |||
| P1 | JX | Waist width (cm) | 41.38 b ± 2.60 (n = 8) | 42.13 ab ± 1.43 (n = 24) | 45.58 a ± 0.61 (n = 69) | 0.015 |
| P1 | JX | Hucklebone width (cm) | 24.17 b ± 2.08 (n = 12) | 23.74 b ± 1.04 (n = 39) | 27.91 a ± 0.50 (n = 85) | 0.000 |
| P1 | NY | Body length (cm) | 146.38 a ± 5.63 (n = 8) | 145.08 a ± 5.41 (n = 12) | 138.13 b ± 1.00 (n =115) | 0.038 |
| TT | TC | CC | pValue | |||
| P3 | LX | Abdominal circumference (cm) | 183.78 c ± 5.89 (n = 9) | 214.71 a ± 12.81 (n = 7) | 198.06 b ± 1.69 (n = 88) | 0.003 |
| GG | GA | AA | pValue | |||
| P4 | NY | Waist width (cm) | 44.59 b ± 0.84 (n = 45) | 49.83 ab ± 0.17 (n = 3) | 51.83 a ± 5.09 (n = 3) | 0.045 |
| P4 | LX | Abdominal circumference (cm) | 197.79 b ± 1.68 (n = 89) | 216.14 a ± 12.43 (n = 7) | 183.75 c ± 6.62 (n = 8) | 0.002 |
| GG | GA | AA | pValue | |||
| N1 | NY | Abdominal circumference (cm) | 174.11 a ± 1.09 (n = 114) | 160.90 b ± 6.72 (n = 10) | 177.77 a ± 6.95 (n = 11) | 0.009 |
| GG | GA | AA | pValue | |||
| N12 | JX | Hucklebone width (cm) | 24.08 b ± 6.36 (n = 12) | 24.72 ab ± 6.61 (n = 32) | 27.27 a ± 5.18 (n = 92) | 0.033 |
| GG | GA | AA | pValue | |||
| N31 | NY | Abdominal circumference (cm) | 219.65 a ± 3.80 (n = 42) | 175.67 b ± 12.39 (n = 3) | 211.17 a ± 6.16 (n = 6) | 0.011 |
| N31 | JX | Body length (cm) | 145.83 a ± 0.89 (n = 106) | 139.27 b ± 1.76 (n = 26) | 142.00 ab ± 5.40 (n = 4) | 0.005 |
| N31 | JX | Waist width (cm) | 45.45 a ± 0.50 (n = 83) | 38.00 b ± 2.29 (n = 15) | 48.33 a ± 2.33 (n = 3) | 0.000 |
| N31 | JX | Hucklebone width (cm) | 27.28 a ± 0.49 (n = 106) | 22.65 b ± 1.38 (n = 26) | 27.00 ab ± 2.65 (n = 4) | 0.001 |
| CC | GC | GG | pValue | |||
| P12 | NY | Abdominal circumference (cm) | 228.03 a ± 3.51 (n = 18) | 217.00 ab ± 8.18 (n = 13) | 204.70 b ± 5.89 (n = 20) | 0.016 |
| P12 | LX | Waist width (cm) | 48.11 a ± 0.80 (n = 28) | 45.50 ab ± 1.14 (n = 28) | 44.23 b ± 0.78 (n = 48) | 0.011 |
| P12 | LX | Body weight (kg) | 426.57 a ± 12.39 (n = 28) | 409.04 ab ± 20.17 (n = 28) | 367.69 b ± 10.27 (n = 48) | 0.029 |
| Morphological Trait | Observed Genotypes (LSMa ± SE) | ||||
|---|---|---|---|---|---|
| AA-CC-AA-TT-GG | GG-CC-GG-TT-GG | GG-TT-AA-CC-AA | GG-TT-AA-CC-GG | p Value | |
| Body height (cm) | 134.28 ± 1.34 a (n = 7) | 137.00 ± 1.96 a (n = 4) | 129.58 ± 1.40 b (n = 12) | 128.02 ± 0.59 b (n = 66) | 0.001 |
| Body length (cm) | 146.00 ± 2.85 a (n = 7) | 152.00 ± 3.14 a (n = 4) | 140.42 ± 1.52 b (n = 12) | 139.11 ± 1.07 b (n = 66) | 0.007 |
| Chest circumference (cm) | 184.57 ± 2.72 a (n = 7) | 189.50 ± 2.60 a (n = 4) | 176.17 ± 2.62 b (n = 12) | 174.16 ± 1.07 b (n = 66) | 0.001 |
| Body weight (kg) | 440.86 ± 11.53 a (n = 7) | 440.25 ± 5.56 a (n = 4) | 398.42 ± 11.41 b (n = 12) | 395.06 ± 6.30 b (n = 66) | 0.040 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Wu, M.; Yi, X.; Tang, X.; Chen, P.; Wang, S.; Sun, X. Functional Analysis of Haplotypes in Bovine PSAP Gene and Their Relationship with Beef Cattle Production Traits. Animals 2021, 11, 49. https://doi.org/10.3390/ani11010049
Zhao H, Wu M, Yi X, Tang X, Chen P, Wang S, Sun X. Functional Analysis of Haplotypes in Bovine PSAP Gene and Their Relationship with Beef Cattle Production Traits. Animals. 2021; 11(1):49. https://doi.org/10.3390/ani11010049
Chicago/Turabian StyleZhao, Haidong, Mingli Wu, Xiaohua Yi, Xiaoqin Tang, Pingbo Chen, Shuhui Wang, and Xiuzhu Sun. 2021. "Functional Analysis of Haplotypes in Bovine PSAP Gene and Their Relationship with Beef Cattle Production Traits" Animals 11, no. 1: 49. https://doi.org/10.3390/ani11010049
APA StyleZhao, H., Wu, M., Yi, X., Tang, X., Chen, P., Wang, S., & Sun, X. (2021). Functional Analysis of Haplotypes in Bovine PSAP Gene and Their Relationship with Beef Cattle Production Traits. Animals, 11(1), 49. https://doi.org/10.3390/ani11010049