Effects of Dietary Fiber on Nutrients Utilization and Gut Health of Poultry: A Review of Challenges and Opportunities



Simple Summary
Abstract
1. Introduction
2. Composition and Properties of DF
3. Antinutritive Effect of DF in Poultry
4. The Beneficial Effect of DF in Poultry Nutrition and Gut Health
4.1. Effects of Fibrous Diet on Nutrient Utilization and Ammonia Emission
4.2. Poultry Gut Microbiome and Its Modulation by DF
4.3. Microbial Fermentation of DF
4.4. Role of SCFA on Gut Health of Poultry
4.5. Effect of DF on the Gut Histomorphometry, Integrity, and Immune Response
5. Effects of Enzymatic Degradation and Processing on the Utilization of DF
6. Scope of Improvement in Poultry Productivity by Exogenous Fiber-Degrading Enzymes
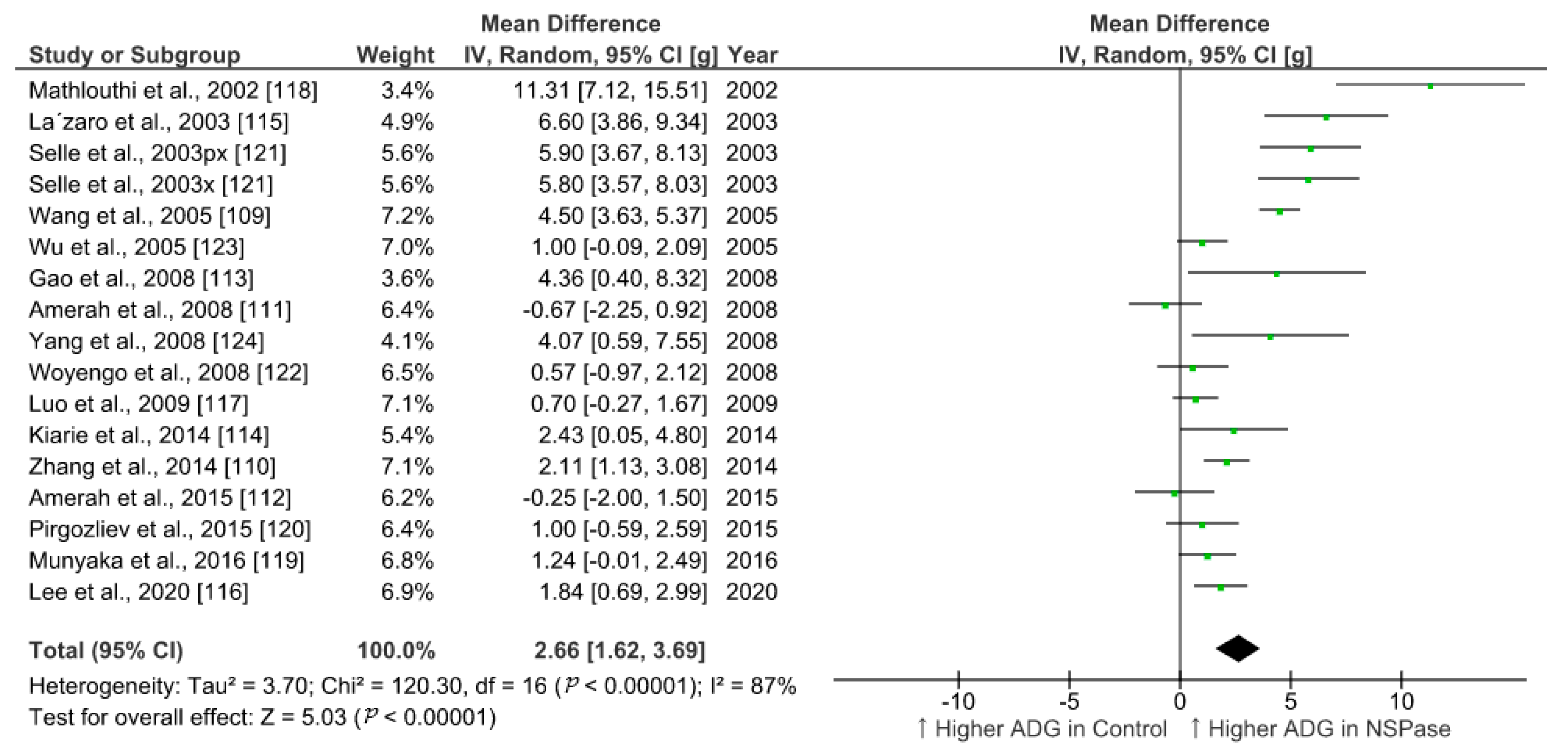
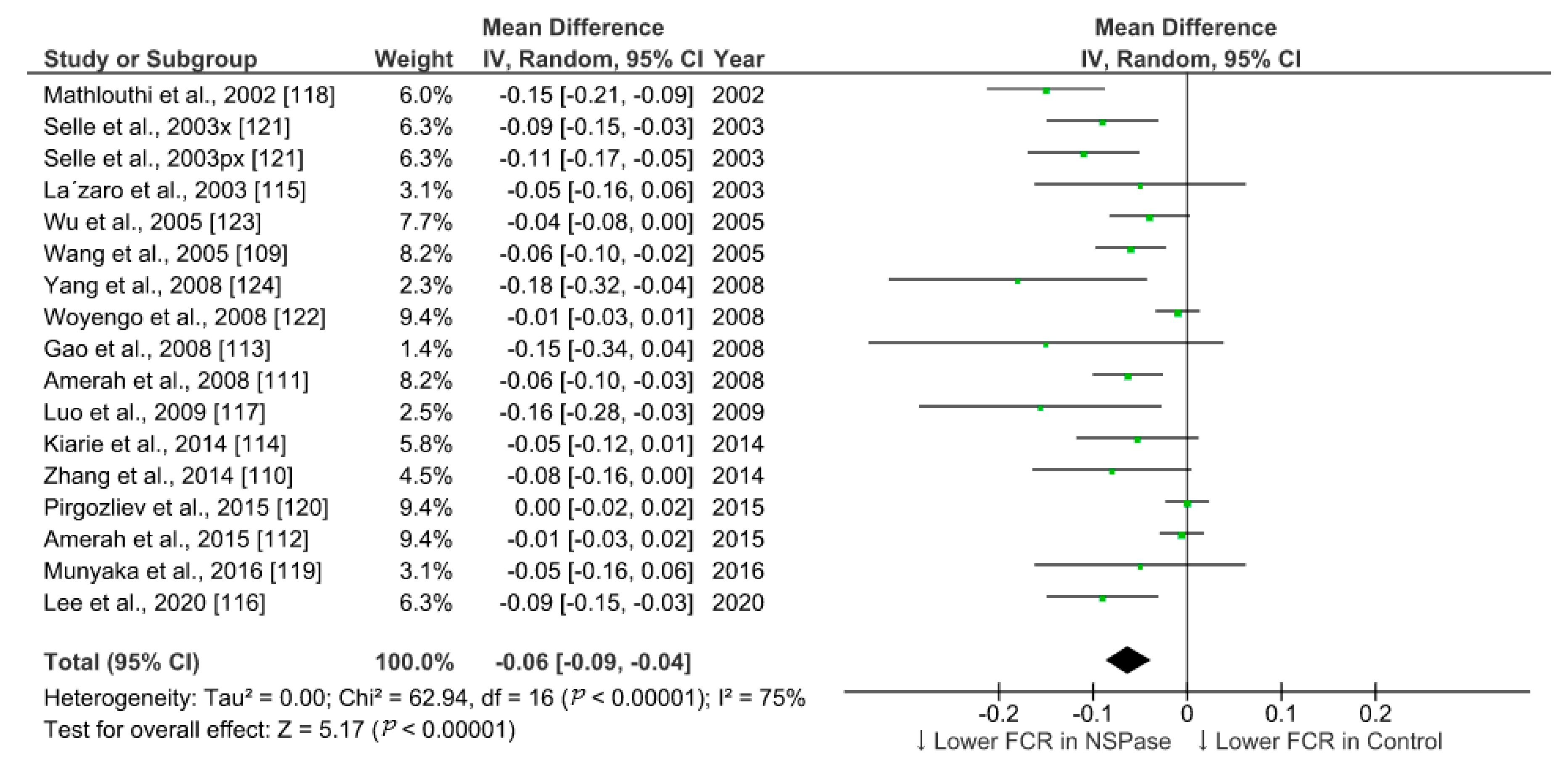
6.1. Meta-Analysis of the Effect of NSPase on Growth Performance of Broilers Fed Fibrous Diet

{kind=link}
{kind=link}
| ADG in NSPase | ADG in Control | FCR in NSPase | FCR in Control | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S.N. | Study | Days | Fiber Source | Enzyme | Rep (N) | Mean, g | ±SEM | Mean, g | ±SEM | Mean | ±SEM | Mean | ±SEM |
| 1 | Amerah et al. 2008 [111] | 21 | w 66.5% | xyl | 6 | 46.5 | 0.57 | 47.2 | 0.57 | 1.35 | 0.013 | 1.41 | 0.013 |
| 2 | Amerah et al. 2015 [112] | 21 | w > 60%, s > 5%, rp 0–6% | xyl + gluc | 8 | 49.6 | 0.63 | 49.9 | 0.63 | 1.34 | 0.008 | 1.35 | 0.008 |
| 3 | Gao et al. 2008 [113] | 21 | w 60% | xyl + gluc + cel + pec | 4 | 37.3 | 1.57 | 32.9 | 1.27 | 1.58 | 0.070 | 1.73 | 0.065 |
| 4 | Kiarie et al. 2014 [114] | 21 | w 60%, wb 9% | xyl + basal phy | 6 | 37.2 | 0.86 | 34.8 | 0.86 | 1.37 | 0.023 | 1.42 | 0.023 |
| 5 | La’zaro et al. 2003 [115] | 25 | r 50% | xyl + gluc | 7 | 38.3 | 0.99 | 31.7 | 0.99 | 1.66 | 0.040 | 1.71 | 0.040 |
| 6 | Lee et al. 2020 [116] | 20 | w 10–20% | xyl | 10 | 36.8 | 0.42 | 34.9 | 0.42 | 1.47 | 0.021 | 1.56 | 0.021 |
| 7 | Luo et al. 2009 [117] | 21 | w 40% | xyl | 5 | 27.9 | 0.35 | 27.2 | 0.35 | 1.56 | 0.046 | 1.71 | 0.046 |
| 8 | Mathlouthi et al. 2002 [118] | 21 | w 40%, b 20% | xyl + gluc | 12 | 38.7 | 1.21 | 27.4 | 1.77 | 1.50 | 0.009 | 1.65 | 0.030 |
| 9 | Munyaka et al. 2016 [119] | 21 | w 42%, b 5%, r 5%, wm 2% | xyl + gluc | 7 | 40.6 | 0.45 | 39.3 | 0.45 | 1.12 | 0.040 | 1.17 | 0.040 |
| 10 | Pirgozliev et al. 2015 [120] | 21 | w 63% | xyl | 6 | 39.4 | 0.57 | 38.4 | 0.57 | 1.35 | 0.008 | 1.35 | 0.008 |
| 11 | Selle et al. 2003 [121] | 24 | w 70% | xyl + phy | 8 | 44.3 | 0.81 | 38.4 | 0.81 | 1.46 | 0.021 | 1.57 | 0.021 |
| 12 | Selle et al. 2003 [121] | 24 | w 70% | xyl | 8 | 44.2 | 0.81 | 38.4 | 0.81 | 1.48 | 0.021 | 1.57 | 0.021 |
| 13 | Wang et al. 2005 [109] | 21 | w 70% | xyl + gluc | 6 | 41.4 | 0.32 | 36.9 | 0.32 | 1.53 | 0.013 | 1.59 | 0.013 |
| 14 | Woyengo et al. 2008 [122] | 21 | w 58% | xyl main effect | 8 | 44.3 | 0.56 | 43.7 | 0.56 | 1.26 | 0.008 | 1.27 | 0.008 |
| 15 | Wu et al. 2005 [123] | 21 | w 66% | xyl | 6 | 38.3 | 0.39 | 37.3 | 0.39 | 1.34 | 0.015 | 1.38 | 0.015 |
| 16 | Yang et al. 2008 [124] | 21 | w 62.4% | xyl | 8 | 56.9 | 1.26 | 52.9 | 1.26 | 1.65 | 0.049 | 1.83 | 0.049 |
| 17 | Zhang et al. 2014 [110] | 21 | w 60% | xyl | 6 | 37.7 | 0.35 | 35.6 | 0.35 | 1.52 | 0.030 | 1.60 | 0.030 |
6.2. Significance of Exogenous Fiber-Degrading Enzymes during Disease Challenge in Poultry
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muscat, A.; de Olde, E.M.; de Boer, I.J.M.; Ripoll-Bosch, R. The battle for biomass: A systematic review of food-feed-fuel competition. Glob. Food Secur. 2019. [Google Scholar] [CrossRef]
- Thebaudin, J.Y.; Lefebvre, A.C.; Harrington, M.; Bourgeois, C.M. Dietary fibers: Nutritional and technological interest. Trends Food Sci. Technol. 1997, 8, 41–48. [Google Scholar] [CrossRef]
- Choct, M.; Dersjant-Li, Y.; McLeish, J.; Peisker, M. Soy oligosaccharides and soluble non-starch polysaccharides: A review of digestion, nutritive and anti-nutritive effects in pigs and poultry. Asian Austral J. Anim. 2010, 23, 1386–1398. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Aftab, U.; Bedford, M.R. The use of NSP enzymes in poultry nutrition: Myths and realities. Worlds Poult. Sci. J. 2018, 74, 277–286. [Google Scholar] [CrossRef]
- Bederska-Łojewska, D.; Świątkiewicz, S.; Arczewska-Włosek, A.; Schwarz, T. Rye non-starch polysaccharides: Their impact on poultry intestinal physiology, nutrients digestibility and performance indices—A review. Ann. Anim. Sci. 2017, 17, 351–369. [Google Scholar] [CrossRef]
- Choct, M.; Annison, G. Anti-nutritive effect of wheat pentosans in broiler chickens: Roles of viscosity and gut microflora. Br. Poult. Sci. 1992, 33, 821–834. [Google Scholar] [CrossRef]
- Teng, P.Y.; Kim, W.K. Review: Roles of prebiotics in intestinal ecosystem of broilers. Front. Vet. Sci 2018, 5, 245. [Google Scholar] [CrossRef]
- Adhikari, P.; Cosby, D.E.; Cox, N.A.; Franca, M.S.; Williams, S.M.; Gogal, R.M., Jr.; Ritz, C.W.; Kim, W.K. Effect of dietary fructooligosaccharide supplementation on internal organs Salmonella colonization, immune response, ileal morphology, and ileal immunohistochemistry in laying hens challenged with Salmonella enteritidis. Poult. Sci. 2018, 97, 2525–2533. [Google Scholar] [CrossRef]
- Rezaei, S.; Faseleh Jahromi, M.; Liang, J.B.; Zulkifli, I.; Farjam, A.S.; Laudadio, V.; Tufarelli, V. Effect of oligosaccharides extract from palm kernel expeller on growth performance, gut microbiota and immune response in broiler chickens. Poult. Sci. 2015, 94, 2414–2420. [Google Scholar] [CrossRef]
- Patterson, J.A.; Burkholder, K.M. Application of prebiotics and probiotics in poultry production. Poult. Sci. 2003, 82, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Saminathan, M.; Sieo, C.C.; Kalavathy, R.; Abdullah, N.; Ho, Y.W. Effect of prebiotic oligosaccharides on growth of Lactobacillus strains used as a probiotic for chickens. Afr. J. Microbiol. Res. 2011, 5, 57–64. [Google Scholar]
- Schley, P.D.; Field, C.J. The immune-enhancing effects of dietary fibers and prebiotics. Br. J. Nutr. 2002, 87 (Suppl. 2), S221–S230. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2006. [Google Scholar]
- Van Soest, P.J. Use of detergents in the analysis of fibrous feeds. I. Preparation of fiber residues of low nitrogen content. J. Assoc. Off. Agric. Chem. 1963, 46, 825–829. [Google Scholar]
- McCleary, B.V.; DeVries, J.W.; Rader, J.I.; Cohen, G.; Prosky, L.; Mugford, D.C.; Champ, M.; Okuma, K. Determination of insoluble, soluble, and total dietary fiber (CODEX definition) by enzymatic-gravimetric method and liquid chromatography: Collaborative study. J. AOAC Int. 2012, 95, 824–844. [Google Scholar] [CrossRef]
- Englyst, H.N.; Quigley, M.E.; Hudson, G.J. Determination of dietary fibre as non-starch polysaccharides with gas–liquid chromatographic, high-performance liquid chromatographic or spectrophotometric measurement of constituent sugars. Analyst 1994, 119, 1497–1509. [Google Scholar] [CrossRef]
- Theander, O.; Åman, P.; Westerlund, E.; Andersson, R.; Pettersson, D. Total dietary fiber determined as neutral sugar residues, uronic acid residues, and Klason lignin (the Uppsala method): Collaborative study. J. AOAC Int. 1995, 78, 1030–1044. [Google Scholar] [CrossRef]
- Mudgil, D.; Barak, S. Composition, properties and health benefits of indigestible carbohydrate polymers as dietary fiber: A review. Int. J. Biol. Macromol. 2013, 61, 1–6. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Dikeman, C.L.; Fahey, G.C., Jr. Viscosity as related to dietary fiber: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 649–663. [Google Scholar] [CrossRef]
- Chutkan, R.; Fahey, G.; Wright, W.L.; McRorie, J. Viscous versus nonviscous soluble fiber supplements: Mechanisms and evidence for fiber-specific health benefits. J. Am. Acad. Nurse Pract. 2012, 24, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Mateos, G.G.; Jiménez-Moreno, E.; Serrano, M.P.; Lázaro, R.P. Poultry response to high levels of dietary fiber sources varying in physical and chemical characteristics. J. Appl. Poult. Res. 2012, 21, 156–174. [Google Scholar] [CrossRef]
- Topping, D. Cereal complex carbohydrates and their contribution to human health. J. Cereal Sci. 2007, 46, 220–229. [Google Scholar] [CrossRef]
- Montagne, L.; Pluske, J.R.; Hampson, D.J. A review of interactions between dietary fiber and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Kluth, H.; Rodehutscord, M. Effect of inclusion of cellulose in the diet on the inevitable endogenous amino acid losses in the ileum of broiler chicken. Poult. Sci. 2009, 88, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Angkanaporn, K.; Choct, M.; Bryden, W.L.; Annison, E.F.; Annison, G. Effects of wheat pentosans on endogenous amino acid losses in chickens. J. Sci. Food Agric. 1994, 66, 399–404. [Google Scholar] [CrossRef]
- Jamroz, D.; Jakobsen, K.; Bach Knudsen, K.E.; Wiliczkiewicz, A.; Orda, J. Digestibility and energy value of non-starch polysaccharides in young chickens, ducks and geese, fed diets containing high amounts of barley. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 131, 657–668. [Google Scholar] [CrossRef]
- Jørgensen, H.; Zhao, X.-Q.; Bach Knudsen, K.E.; Eggum, B.O. The influence of dietary fibre source and level on the development of the gastrointestinal tract, digestibility and energy metabolism in broiler chickens. Br. J. Nutr. 1996, 75, 379–395. [Google Scholar] [CrossRef]
- Rodriguez, M.L.; Rebole, A.; Velasco, S.; Ortiz, L.T.; Trevino, J.; Alzueta, C. Wheat- and barley-based diets with or without additives influence broiler chicken performance, nutrient digestibility and intestinal microflora. J. Sci. Food Agric. 2012, 92, 184–190. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.-B. Coarse particle inclusion and lignocellulose-rich fiber addition in feed benefit performance and health of broiler chickens. Poult. Sci. 2017, 96, 3272–3281. [Google Scholar] [CrossRef]
- Capuano, E. The behavior of dietary fiber in the gastrointestinal tract determines its physiological effect. Crit. Rev. Food Sci. Nutr. 2017, 57, 3543–3564. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, G.; van Milgen, J.; Noblet, J. Influence of dietary fibre on digestive utilization and rate of passage in growing pigs, finishing pigs and adult sows. Anim. Sci. 2002, 74, 503–515. [Google Scholar] [CrossRef]
- Wilfart, A.; Montagne, L.; Simmins, H.; Noblet, J.; van Milgen, J. Digesta transit in different segments of the gastrointestinal tract of pigs as affected by insoluble fibre supplied by wheat bran. Br. J. Nutr. 2007, 98, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Choct, M.; Kocher, A. Non-Starch Carbohydrates: Digestion and Its Secondary Effects in Monogastrics; Nutrition Society of Australia: Crows Nest, NSW, Australia, 2000; pp. 31–38. [Google Scholar]
- Liu, Z.; Wang, L.; Beasley, D.; Oviedo, E. Effect of moisture content on ammonia emissions from broiler litter: A laboratory study. J. Atmos. Chem. 2007, 58, 41–53. [Google Scholar] [CrossRef]
- Shepherd, E.M.; Fairchild, B.D. Footpad dermatitis in poultry. Poult. Sci. 2010, 89, 2043–2051. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Acamovic, T.; Bedford, M.R. The effects of phytase and phytic acid on the loss of endogenous amino acids and minerals from broiler chickens. Br. Poult. Sci. 2004, 45, 101–108. [Google Scholar] [CrossRef]
- Kumar, V.; Sinha, A.K.; Makkar, H.P.S.; Becker, K. Dietary roles of phytate and phytase in human nutrition: A review. Food Chem. 2010, 120, 945–959. [Google Scholar] [CrossRef]
- Oatway, L.; Vasanthan, T.; Helm, J.H. Phytic acid. Food Rev. Int. 2001, 17, 419–431. [Google Scholar] [CrossRef]
- Latshaw, J.D.; Zhao, L. Dietary protein effects on hen performance and nitrogen excretion. Poult. Sci. 2011, 90, 99–106. [Google Scholar] [CrossRef]
- Kim, W.K.; Patterson, P.H. Effect of minerals on activity of microbial uricase to reduce ammonia volatilization in poultry manure. Poult. Sci. 2003, 82, 223–231. [Google Scholar] [CrossRef]
- Piva, A.; Panciroli, A.; Meola, E.; Formigoni, A. Lactitol enhances short-chain fatty acid and gas production by swine cecal microflora to a greater extent when fermenting low rather than high fiber diets. J. Nutr. 1996, 126, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Leterme, P. Feed ingredients differing in fermentable fiber and indigestible protein content affect fermentation metabolites and faecal nitrogen excretion in growing pigs. Animal 2012, 6, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Berrocoso, J.F.D. Dietary fiber and protein fermentation in the intestine of swine and their interactive effects on gut health and on the environment: A review. Anim. Feed Sci. Technol. 2016, 212, 18–26. [Google Scholar] [CrossRef]
- Metges, C.C. Contribution of microbial amino acids to amino acid homeostasis of the host. J. Nutr. 2000, 130, 1857S–1864S. [Google Scholar] [CrossRef]
- Roberts, S.A.; Xin, H.; Kerr, B.J.; Russell, J.R.; Bregendahl, K. Effects of dietary fiber and reduced crude protein on ammonia emission from laying-hen manure. Poult. Sci. 2007, 86, 1625–1632. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Honda, K. Intestinal commensal microbes as immune modulators. Cell Host Microbe 2012, 12, 496–508. [Google Scholar] [CrossRef]
- Littman, D.R.; Pamer, E.G. Role of the commensal microbiota in normal and pathogenic host immune responses. Cell Host Microbe 2011, 10, 311–323. [Google Scholar] [CrossRef]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Apajalahti, J.; Kettunen, A.; Graham, H. Characteristics of the gastrointestinal microbial communities, with special reference to the chicken. Worlds Poult. Sci. J. 2004, 60, 223–232. [Google Scholar] [CrossRef]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci 2018, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Pokorna, A.; Cizek, A.; et al. Contact with adult hen affects development of caecal microbiota in newly hatched chicks. PLoS ONE 2019, 14, e0212446. [Google Scholar] [CrossRef] [PubMed]
- Rinttilä, T.; Apajalahti, J. Intestinal microbiota and metabolites—Implications for broiler chicken health and performance. J. Appl. Poult. Res. 2013, 22, 647–658. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Gibson, G.R. Carbohydrate fermentation, energy transduction and gas metabolism in the human large intestine. In Gastrointestinal Microbiology: Gastrointestinal Ecosystems and Fermentations; Mackie, R.I., White, B.A., Eds.; Springer: Boston, MA, USA, 1997; Volume 1, pp. 269–318. [Google Scholar]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and environmental factors affecting the intestinal microbiota in chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Janardhana, V.; Broadway, M.M.; Bruce, M.P.; Lowenthal, J.W.; Geier, M.S.; Hughes, R.J.; Bean, A.G. Prebiotics modulate immune responses in the gut-associated lymphoid tissue of chickens. J. Nutr. 2009, 139, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Shoaf, K.; Mulvey, G.L.; Armstrong, G.D.; Hutkins, R.W. Prebiotic galactooligosaccharides reduce adherence of enteropathogenic Escherichia coli to tissue culture cells. Infect. Immun. 2006, 74, 6920–6928. [Google Scholar] [CrossRef] [PubMed]
- Spring, P.; Wenk, C.; Dawson, K.A.; Newman, K.E. The effects of dietary mannaoligosaccharides on cecal parameters and the concentrations of enteric bacteria in the ceca of salmonella-challenged broiler chicks. Poult. Sci. 2000, 79, 205–211. [Google Scholar] [CrossRef]
- Jung, S.J.; Houde, R.; Baurhoo, B.; Zhao, X.; Lee, B.H. Effects of galacto-oligosaccharides and a Bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens. Poult. Sci. 2008, 87, 1694–1699. [Google Scholar] [CrossRef]
- Baurhoo, B.; Phillip, L.; Ruiz-Feria, C.A. Effects of purified lignin and mannan oligosaccharides on intestinal integrity and microbial populations in the ceca and litter of broiler chickens. Poult. Sci. 2007, 86, 1070–1078. [Google Scholar] [CrossRef]
- Snel, J.; Harmsen, H.J.M.; Van der Wielen, P.W.J.J.; Williams, B.A. Dietary strategies to influence the gastrointestinal microflora of young animals, and its potential to improve intestinal health. In Nutrition and Health on the Gastrointestinal Tract; Blok, M.C., Vahl, H.A., de Lange, L., van de Braak, A.E., Hemke, G., Hessing, M., Eds.; Wageningen Academic Publishers: Wageningen, NL, USA, 2002; pp. 37–69. [Google Scholar]
- Singh, A.K. In Ovo and Post-Hatch Nutritional Programming to Improve Broiler Performance and Gut Health. Ph.D. Thesis, University of Hawai’i at Manoa, Ann Arbor, MI, USA, 2019. Available online: https://scholarspace.manoa.hawaii.edu/bitstream/10125/63507/1/Singh_hawii_0085A_10392.pdf (accessed on 1 December 2020).
- Dunkley, K.D.; Dunkley, C.S.; Njongmeta, N.L.; Callaway, T.R.; Hume, M.E.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Comparison of in vitro fermentation and molecular microbial profiles of high-fiber feed substrates incubated with chicken cecal inocula. Poult. Sci. 2007, 86, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, D.; Kaczmarek, S.; Rutkowski, A.; Józefiak, A.; Jensen, B.B.; Engberg, R.M. Fermentation in broiler chicken gastrointestinal tract as affected by high dietary inclusion of barley and β-glucanase supplementation. J. Anim. Feed Sci. 2005, 14, 695. [Google Scholar] [CrossRef]
- Walugembe, M.; Hsieh, J.C.F.; Koszewski, N.J.; Lamont, S.J.; Persia, M.E.; Rothschild, M.F. Effects of dietary fiber on cecal short-chain fatty acid and cecal microbiota of broiler and laying-hen chicks. Poult. Sci. 2015, 94, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Ricke, S.C.; Woodward, C.L.; Kwon, Y.M.; Kubena, L.F.; Nisbet, D.J. Limiting avian gastrointestinal tract Salmonella colonization by cecal anaerobic bacteria, and a potential role for methanogens. In Preharvest and Postharvest Food Safety: Contemporary Issues and Future Directions; Beier, R.C., Pillai, S.D., Phillip, T.D., Eds.; Blackwell Publishing: Ames, IA, USA, 2004; p. 141. [Google Scholar]
- Meimandipour, A.; Shuhaimi, M.; Soleimani, A.F.; Azhar, K.; Hair-Bejo, M.; Kabeir, B.M.; Javanmard, A.; Muhammad Anas, O.; Yazid, A.M. Selected microbial groups and short-chain fatty acids profile in a simulated chicken cecum supplemented with two strains of Lactobacillus. Poult. Sci. 2010, 89, 470–476. [Google Scholar] [CrossRef]
- Reichardt, N.; Vollmer, M.; Holtrop, G.; Farquharson, F.M.; Wefers, D.; Bunzel, M.; Duncan, S.H.; Drew, J.E.; Williams, L.M.; Milligan, G.; et al. Specific substrate-driven changes in human faecal microbiota composition contrast with functional redundancy in short-chain fatty acid production. ISME J. 2018, 12, 610–622. [Google Scholar] [CrossRef]
- Peng, Q.; Zeng, X.F.; Zhu, J.L.; Wang, S.; Liu, X.T.; Hou, C.L.; Thacker, P.A.; Qiao, S.Y. Effects of dietary Lactobacillus plantarum B1 on growth performance, intestinal microbiota, and short chain fatty acid profiles in broiler chickens. Poult. Sci. 2016, 95, 893–900. [Google Scholar] [CrossRef]
- Lee, S.A.; Apajalahti, J.; Vienola, K.; González-Ortiz, G.; Fontes, C.M.G.A.; Bedford, M.R. Age and dietary xylanase supplementation affects ileal sugar residues and short chain fatty acid concentration in the ileum and caecum of broiler chickens. Anim. Feed Sci. Technol. 2017, 234, 29–42. [Google Scholar] [CrossRef]
- Rehman, H.; Hellweg, P.; Taras, D.; Zentek, J. Effects of dietary inulin on the intestinal short chain fatty acids and microbial ecology in broiler chickens as revealed by denaturing gradient gel electrophoresis. Poult. Sci. 2008, 87, 783–789. [Google Scholar] [CrossRef]
- Scheppach, W. Effects of short chain fatty acids on gut morphology and function. Gut 1994, 35, S35–S38. [Google Scholar] [CrossRef]
- Yan, H.; Ajuwon, K.M. Butyrate modifies intestinal barrier function in IPEC-J2 cells through a selective upregulation of tight junction proteins and activation of the Akt signaling pathway. PLoS ONE 2017, 12, e0179586. [Google Scholar] [CrossRef] [PubMed]
- Kamath, P.S.; Hoepfner, M.T.; Phillips, S.F. Short-chain fatty acids stimulate motility of the canine ileum. Am. J. Physiol. Gastrointest. Liver Physiol. 1987, 253, G427–G433. [Google Scholar] [CrossRef] [PubMed]
- Binder, H.J.; Mehta, P. Short-chain fatty acids stimulate active sodium and chloride absorption in vitro in the rat distal colon. Gastroenterology 1989, 96, 989–996. [Google Scholar] [CrossRef]
- Fukunaga, T.; Sasaki, M.; Araki, Y.; Okamoto, T.; Yasuoka, T.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T. Effects of the soluble fiber pectin on intestinal cell proliferation, fecal short chain fatty acid production and microbial population. Digestion 2003, 67, 42–49. [Google Scholar] [CrossRef]
- Fernández-Rubio, C.; Ordonez, C.; Abad-González, J.; Garcia-Gallego, A.; Honrubia, M.P.; Mallo, J.J.; Balana-Fouce, R. Butyric acid-based feed additives help protect broiler chickens from Salmonella Enteritidis infection. Poult. Sci. 2009, 88, 943–948. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Fievez, V.; de Buck, J.; Pasmans, F.; Martel, A.; Haesebrouck, F.; Ducatelle, R. Microencapsulated short-chain fatty acids in feed modify colonization and invasion early after infection with Salmonella enteritidis in young chickens. Poult. Sci. 2004, 83, 69–74. [Google Scholar] [CrossRef]
- Leeson, S.; Namkung, H.; Antongiovanni, M.; Lee, E.H. Effect of butyric acid on the performance and carcass yield of broiler chickens. Poult. Sci. 2005, 84, 1418–1422. [Google Scholar] [CrossRef]
- van Der Wielen, P.W.; Biesterveld, S.; Notermans, S.; Hofstra, H.; Urlings, B.A.; van Knapen, F. Role of volatile fatty acids in development of the cecal microflora in broiler chickens during growth. Appl. Environ. Microbiol. 2000, 66, 2536–2540. [Google Scholar] [CrossRef]
- Shyer, A.E.; Tallinen, T.; Nerurkar, N.L.; Wei, Z.; Gil, E.S.; Kaplan, D.L.; Tabin, C.J.; Mahadevan, L. Villification: How the gut gets its villi. Science 2013, 342, 212–218. [Google Scholar] [CrossRef]
- Andoh, A.; Bamba, T.; Sasaki, M. Physiological and anti-inflammatory roles of dietary fiber and butyrate in intestinal functions. JPEN J. Parenter. Enter. Nutr. 1999, 23, S70–S73. [Google Scholar] [CrossRef]
- Shang, Y.; Regassa, A.; Kim, J.H.; Kim, W.K. The effect of dietary fructooligosaccharide supplementation on growth performance, intestinal morphology, and immune responses in broiler chickens challenged with Salmonella Enteritidis lipopolysaccharides. Poult. Sci. 2015, 94, 2887–2897. [Google Scholar] [CrossRef]
- Ashraf, S.; Zaneb, H.; Yousaf, M.S.; Ijaz, A.; Sohail, M.U.; Muti, S.; Usman, M.M.; Ijaz, S.; Rehman, H. Effect of dietary supplementation of prebiotics and probiotics on intestinal microarchitecture in broilers reared under cyclic heat stress. J. Anim. Physiol. Anim. Nutr. (Berl.) 2013, 97 (Suppl. 1), 68–73. [Google Scholar] [CrossRef]
- Amat, C.; Planas, J.M.; Moreto, M. Kinetics of hexose uptake by the small and large intestine of the chicken. Am. J. Physiol. 1996, 271, R1085–R1089. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564. [Google Scholar] [CrossRef]
- Teng, P.-Y.; Yadav, S.; de Souza Castro, F.L.; Tompkins, Y.H.; Fuller, A.L.; Kim, W.K. Graded Eimeria challenge linearly regulated growth performance, dynamic change of gastrointestinal permeability, apparent ileal digestibility, intestinal morphology, and tight junctions of broiler chickens. Poult. Sci. 2020, 99, 4203–4216. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Guo, S.; Guo, Y. Xylanase supplementation to a wheat-based diet alleviated the intestinal mucosal barrier impairment of broiler chickens challenged by Clostridium perfringens. Avian Pathol. 2012, 41, 291–298. [Google Scholar] [CrossRef]
- Wang, H.B.; Wang, P.Y.; Wang, X.; Wan, Y.L.; Liu, Y.C. Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein claudin-1 transcription. Dig. Dis. Sci. 2012, 57, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Wu, D.; Lewis, E.D.; Pae, M.; Meydani, S.N. Nutritional modulation of immune function: Analysis of evidence, mechanisms, and clinical relevance. Front. Immunol. 2019, 9, 3160. [Google Scholar] [CrossRef] [PubMed]
- Cunningham-Rundles, S.; Lin, H.; Ho-Lin, D.; Dnistrian, A.; Cassileth, B.R.; Perlman, J.M. Role of nutrients in the development of neonatal immune response. Nutr. Rev. 2009, 67, S152–S163. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Verduzco, G.; Cortes-Cuevas, A.; López-Coello, C.; Ávila-González, E.; Nava, G.M. Dietary supplementation of mannan-oligosaccharide enhances neonatal immune responses in chickens during natural exposure to Eimeria spp. Acta Vet. Scand. 2009, 51, 11. [Google Scholar] [CrossRef]
- Min, Y.N.; Li, L.L.; Liu, S.K.; Zhang, J.; Gao, Y.P.; Liu, F.Z. Effects of dietary distillers dried grains with solubles (DDGS) on growth performance, oxidative stress, and immune function in broiler chickens. J. Appl. Poult. Res. 2015, 24, 23–29. [Google Scholar] [CrossRef]
- Sadeghi, A.; Toghyani, M.; Gheisari, A. Effect of various fiber types and choice feeding of fiber on performance, gut development, humoral immunity, and fiber preference in broiler chicks. Poult. Sci. 2015, 94, 2734–2743. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of heat stress on poultry production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Vicuña, E.A.; Kuttappan, V.A.; Galarza-Seeber, R.; Latorre, J.D.; Faulkner, O.B.; Hargis, B.M.; Tellez, G.; Bielke, L.R. Effect of dexamethasone in feed on intestinal permeability, differential white blood cell counts, and immune organs in broiler chicks. Poult. Sci. 2015, 94, 2075–2080. [Google Scholar] [CrossRef]
- Jia, W.; Slominski, B.A.; Bruce, H.L.; Blank, G.; Crow, G.; Jones, O. Effects of diet type and enzyme addition on growth performance and gut health of broiler chickens during subclinical Clostridium perfringens challenge. Poult. Sci. 2009, 88, 132–140. [Google Scholar] [CrossRef]
- Bedford, M.R.; Apajalahti, J. Microbial interactions in response to Exogenous enzyme utilization. In Enzymes in Farm Animal Nutrition; Bedford, M.R., Partrige, G.G., Eds.; CAB International: Wallingford, UK, 2001; pp. 299–314. [Google Scholar]
- De Vries, S.; Pustjens, A.M.; Schols, H.A.; Hendriks, W.H.; Gerrits, W.J.J. Improving digestive utilization of fiber-rich feedstuffs in pigs and poultry by processing and enzyme technologies: A review. Anim. Feed Sci. Technol. 2012, 178, 123–138. [Google Scholar] [CrossRef]
- Teitge, D.A.; Campbell, G.L.; Classen, H.L.; Thacker, P.A. Heat pretreatment as a means of improving the response to dietary pentosanase in chicks fed rye. Can. J. Anim. Sci. 1991, 71, 507–513. [Google Scholar] [CrossRef]
- Review Manager (RevMan). Computer Program, Version 5.4; The Cochrane Collaboration: London, UK; The Nordic Cochrane Center: Copenhagen, Denmark, 2020. [Google Scholar]
- Wang, Z.R.; Qiao, S.Y.; Lu, W.Q.; Li, D.F. Effects of enzyme supplementation on performance, nutrient digestibility, gastrointestinal morphology, and volatile fatty acid profiles in the hindgut of broilers fed wheat-based diets. Poult. Sci. 2005, 84, 875–881. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, J.; Lei, L.; Jiang, Y.; Gao, F.; Zhou, G.H. Effects of xylanase supplementation on growth performance, nutrient digestibility and non-starch polysaccharide degradation in different sections of the gastrointestinal tract of broilers fed wheat-based diets. Asian Austral. J. Anim 2014, 27, 855. [Google Scholar] [CrossRef] [PubMed]
- Amerah, A.M.; Ravindran, V.; Lentle, R.G. Influence of wheat hardness and xylanase supplementation on the performance, energy utilisation, digestive tract development and digesta parameters of broiler starters. Anim. Prod. Sci. 2008, 49, 71–78. [Google Scholar] [CrossRef]
- Amerah, A.M.; Van de Belt, K.; van Der Klis, J.D. Effect of different levels of rapeseed meal and sunflower meal and enzyme combination on the performance, digesta viscosity and carcass traits of broiler chickens fed wheat-based diets. Animal 2015, 9, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Jiang, Y.; Zhou, G.H.; Han, Z.K. The effects of xylanase supplementation on performance, characteristics of the gastrointestinal tract, blood parameters and gut microflora in broilers fed on wheat-based diets. Anim. Feed Sci. Technol. 2008, 142, 173–184. [Google Scholar] [CrossRef]
- Kiarie, E.; Romero, L.F.; Ravindran, V. Growth performance, nutrient utilization, and digesta characteristics in broiler chickens fed corn or wheat diets without or with supplemental xylanase. Poult. Sci. 2014, 93, 1186–1196. [Google Scholar] [CrossRef]
- Lazaro, R.; Garcia, M.; Medel, P.; Mateos, G.G. Influence of enzymes on performance and digestive parameters of broilers fed rye-based diets. Poult. Sci. 2003, 82, 132–140. [Google Scholar] [CrossRef]
- Lee, S.H.; Hosseindoust, A.; Laxman Ingale, S.; Rathi, P.C.; Yoon, S.Y.; Choi, J.W.; Kim, J.S. Thermostable xylanase derived from Trichoderma citrinoviride increases growth performance and non-starch polysaccharide degradation in broiler chickens. Br. Poult. Sci. 2020, 61, 57–62. [Google Scholar] [CrossRef]
- Luo, D.; Yang, F.; Yang, X.; Yao, J.; Shi, B.; Zhou, Z. Effects of xylanase on performance, blood parameters, intestinal morphology, microflora and digestive enzyme activities of broilers fed wheat-based diets. Asian Austral. J. Anim. 2009, 22, 1288–1295. [Google Scholar] [CrossRef]
- Mathlouthi, N.; Mallet, S.; Saulnier, L.; Quemener, B.; Larbier, M. Effects of xylanase and β-glucanase addition on performance, nutrient digestibility, and physico-chemical conditions in the small intestine contents and caecal microflora of broiler chickens fed a wheat and barley-based diet. Anim. Res. 2002, 51, 395–406. [Google Scholar] [CrossRef]
- Munyaka, P.M.; Nandha, N.K.; Kiarie, E.; Nyachoti, C.M.; Khafipour, E. Impact of combined beta-glucanase and xylanase enzymes on growth performance, nutrients utilization and gut microbiota in broiler chickens fed corn or wheat-based diets. Poult. Sci. 2016, 95, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Pirgozliev, V.; Rose, S.P.; Pellny, T.; Amerah, A.M.; Wickramasinghe, M.; Ulker, M.; Rakszegi, M.; Bedo, Z.; Shewry, P.R.; Lovegrove, A. Energy utilization and growth performance of chickens fed novel wheat inbred lines selected for different pentosan levels with and without xylanase supplementation. Poult. Sci. 2015, 94, 232–239. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Selle, P.; Ravindran, V.; Ravindran, G.; Pittolo, P.; Bryden, W. Influence of phytase and xylanase supplementation on growth performance and nutrient utilisation of broilers offered wheat-based diets. Asian-australas. J. Anim. Sci. 2003, 16, 394–402. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Guenter, W.; Sands, J.S.; Nyachoti, C.M.; Mirza, M.A. Nutrient utilisation and performance responses of broilers fed a wheat-based diet supplemented with phytase and xylanase alone or in combination. Anim. Feed Sci. Technol. 2008, 146, 113–123. [Google Scholar] [CrossRef]
- Wu, Y.; Lai, C.; Qiao, S.; Gong, L.; Lu, W.; Li, D. Properties of Aspergillar xylanase and the effects of xylanase supplementation in wheat-based diets on growth performance and the blood biochemical values in broilers. Asian Austral. J. Anim. 2005, 18, 66–74. [Google Scholar] [CrossRef]
- Yang, Y.; Iji, P.A.; Kocher, A.; Mikkelsen, L.L.; Choct, M. Effects of xylanase on growth and gut development of broiler chickens given a wheat-based diet. Asian Austral. J. Anim. 2008, 21, 1659–1664. [Google Scholar] [CrossRef]
- Amerah, A.M.; Mathis, G.; Hofacre, C.L. Effect of xylanase and a blend of essential oils on performance and Salmonella colonization of broiler chickens challenged with Salmonella Heidelberg. Poult. Sci. 2012, 91, 943–947. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, D.; Guo, S.; Chen, Y.; Guo, Y. Effects of dietary essential oil and enzyme supplementation on growth performance and gut health of broilers challenged by Clostridium perfringens. Anim. Feed Sci. Technol. 2015, 207, 234–244. [Google Scholar] [CrossRef]
- Jia, W.; Slominski, B.A.; Bruce, H.L.; Nyachoti, C.M.; Jones, R.O. Enzyme addition facilitates the post-disease compensatory growth of broiler chickens challenged with Clostridium perfringens. Can. J. Anim. Sci. 2009, 89, 369–381. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Gibbs, K.; Awati, A.; Klasing, K.C. The effects of enzymes and direct fed microbial combination on performance and immune response of broilers under a coccidia challenge. J. Appl. Anim. Nutr. 2016, 4. [Google Scholar] [CrossRef]
- Jackson, M.E.; Anderson, D.M.; Hsiao, H.Y.; Mathis, G.F.; Fodge, D.W. Beneficial effect of β-mannanase feed enzyme on performance of chicks challenged with Eimeria sp. and Clostridium perfringens. Avian Dis. 2003, 47, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Choct, M.; Sinlae, M.; Al-Jassim, R.A.M.; Pettersson, D. Effects of xylanase supplementation on between-bird variation in energy metabolism and the number of Clostridium perfringens in broilers fed a wheat-based diet. Aust. J. Agric. Res. 2006, 57, 1017–1021. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Scapini, L.B.; Ribeiro, M.V.; Pivetta, M.R.; Buzim, R.; Fernandes, J.I.M. Effects of β-mannanase supplementation on the intestinal microbiota composition of broiler chickens challenged with a coccidiosis vaccine. Livest. Sci. 2019, 228, 187–194. [Google Scholar] [CrossRef]
- Khattak, F.M.; Pasha, T.N.; Hayat, Z.; Mahmud, A. Enzymes in poultry nutrition. J. Anim. Plant Sci. 2006, 16, 1–7. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.K.; Kim, W.K. Effects of Dietary Fiber on Nutrients Utilization and Gut Health of Poultry: A Review of Challenges and Opportunities. Animals 2021, 11, 181. https://doi.org/10.3390/ani11010181
Singh AK, Kim WK. Effects of Dietary Fiber on Nutrients Utilization and Gut Health of Poultry: A Review of Challenges and Opportunities. Animals. 2021; 11(1):181. https://doi.org/10.3390/ani11010181
Chicago/Turabian StyleSingh, Amit Kumar, and Woo Kyun Kim. 2021. "Effects of Dietary Fiber on Nutrients Utilization and Gut Health of Poultry: A Review of Challenges and Opportunities" Animals 11, no. 1: 181. https://doi.org/10.3390/ani11010181
APA StyleSingh, A. K., & Kim, W. K. (2021). Effects of Dietary Fiber on Nutrients Utilization and Gut Health of Poultry: A Review of Challenges and Opportunities. Animals, 11(1), 181. https://doi.org/10.3390/ani11010181