Genetic Diversity and Population Structure of Polish Konik Horse Based on Individuals from All the Male Founder Lines and Microsatellite Markers

 , , , ,
, , , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
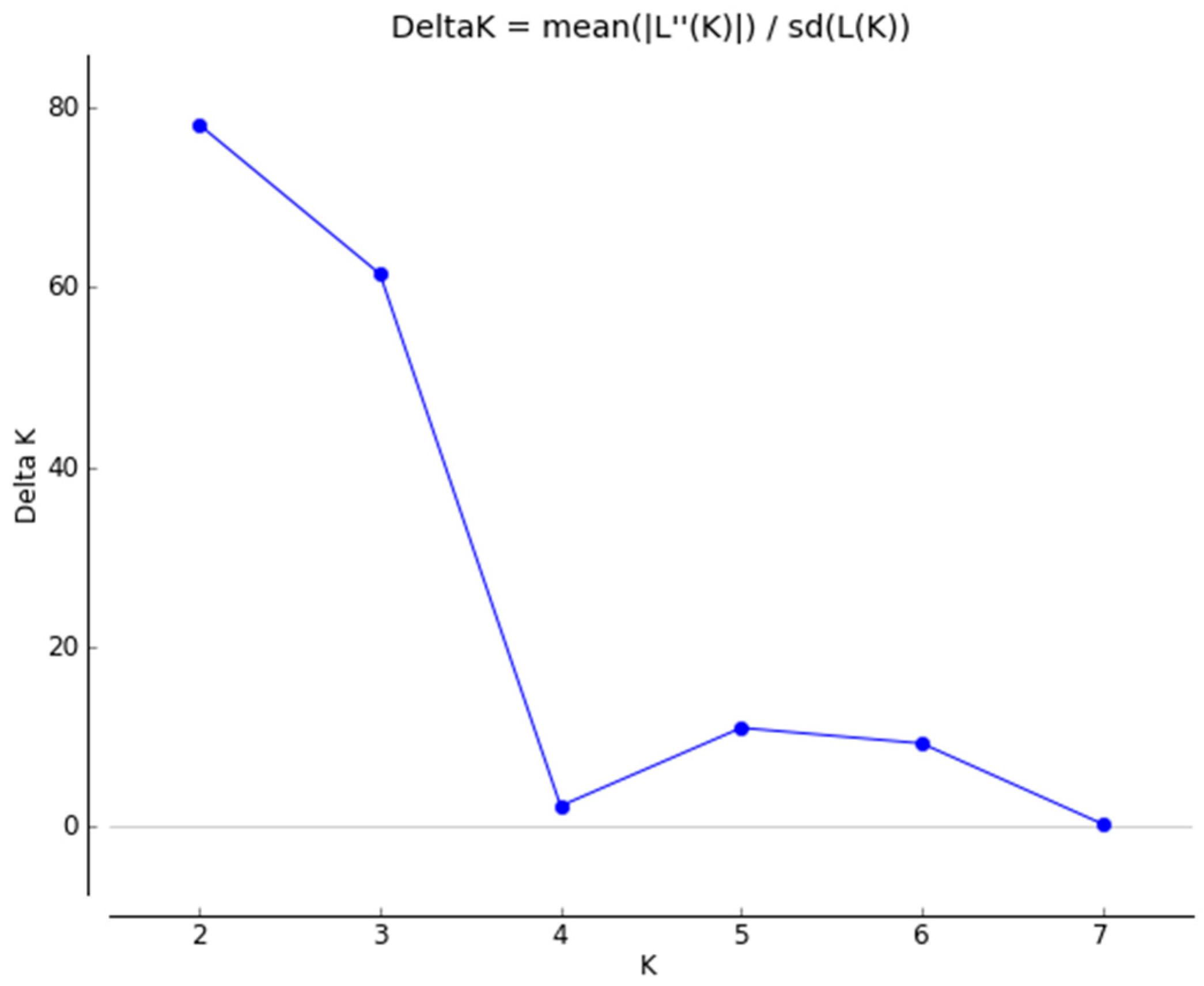
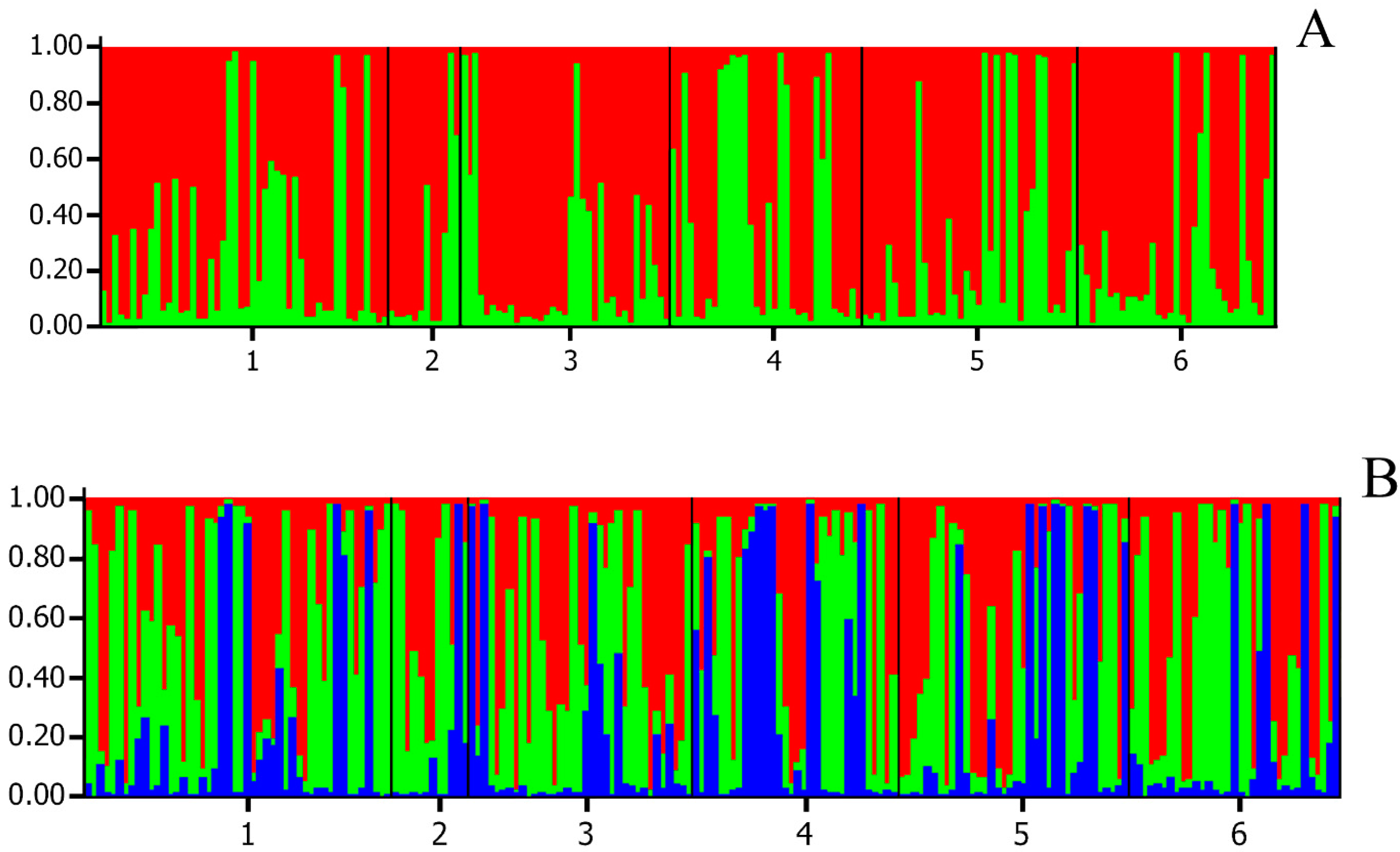
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gralak, B.; Niemczewski, C.; Jaworski, Z. Genetic polymorphism of 12 micorsatellite markers in Polish Primitive Horse. Anim. Sci. Pap. Rep. 2001, 19, 277–283. [Google Scholar]
- Doboszewski, P.; Doktor, D.; Jaworski, Z.; Kalski, R.; Kulakowska, G.; Lojek, J.; Plachocki, D.; Rys, A.; Tylkowska, A. Konik polski horses as a mean of biodiversity maintenance in post-agricultural and forest areas: An overview of Polish experiences. Anim. Sci. Pap. Rep. 2017, 35, 333–347. [Google Scholar]
- Janczarek, I.; Pluta, M.; Paszkowska, A. Pochodzenie, hodowla i użytkowanie koników polskich. Prz. Hod. 2017, 85, 6–10. [Google Scholar]
- Stefaniuk-Szmukier, M.; Ropka-Molik, K.; Piorkowska, K.; Szmatoła, T.; Długosz, B.; Pisarczyk, W.; Bugno-Poniewierska, M. Variation in TBX3 Gene Region in Dun Coat Color Polish Konik Horses. J. Equine Vet. Sci. 2017, 49, 60–62. [Google Scholar] [CrossRef]
- Komosa, M.; Purzyc, H. Konik and Hucul horses: A comparative study of exterior measurements. J. Anim. Sci. 2009, 87, 2245–2254. [Google Scholar] [CrossRef]
- Jodkowska, E.; Kuźniak, A.M.; Pikuła, R.; Smugała, M. Analysis of Polish Primitive Ponies Migration under Registration of the Global Positioning System. GSTF J. Vet. Sci. 2015, 1, 47–68. [Google Scholar] [CrossRef]
- Gurgul, A.; Jasielczuk, I.; Semik-Gurgul, E.; Pawlina-Tyszko, K.; Stefaniuk-Szmukier, M.; Szmatoła, T.; Polak, G.; Tomczyk-Wrona, I.; Bugno-Poniewierska, M. A genome-wide scan for diversifying selection signatures in selected horse breeds. PLoS ONE 2019, 14, 1–24. [Google Scholar] [CrossRef]
- Pluta, M.; Osinski, Z.; Cieśla, A.; Kolstrung, R. Genetic and Phenotypic Characteristics of Polish Konik Horses Maintained in the Reserve and Stable System in Central-Eastern Poland. Acta Sci. Pol. Zootech. 2016, 15, 59–76. [Google Scholar] [CrossRef]
- Pasicka, E. Polish konik horse-characteristics and historical background of native descendants of tarpan. Acta Sci. Pol. Med. Vet. 2013, 12, 25–38. [Google Scholar]
- Siemieniuch, M.J.; Jaworski, Z. 70-lecie hodowli konikow polskich w Popielnie. Hod. Jezdziec. 2020, 1, 28–30. [Google Scholar]
- Cieslak, J.; Wodas, L.; Borowska, A.; Cothran, E.G.; Khanshour, A.M.; Mackowski, M. Characterization of the Polish Primitive Horse (Konik) maternal lines using mitochondrial D-loop sequence variation. PeerJ 2017, 5, e3714. [Google Scholar] [CrossRef]
- Kaminski, S.; Hering, D.M.; Jaworski, Z.; Zabolewicz, T.; Rusc, A.; Kaminski, S.; Hering, D.M.; Jaworski, Z.; Zabolewicz, T.; Rusc, A. Assessment of genomic inbreeding in Polish Konik horses. Pol. J. Vet. Sci. 2017, 20, 603–605. [Google Scholar] [CrossRef]
- Jaworski, Z.; Tomczyk-Wrona, I. Program. Ochrony Zasobów Genetycznych Koni Rasy Konik Polski; Krajowy Ośrodek Koordynacyjny ds. Zasobów Genetycznych Zwierząt, Instytut Zootechniki-PIB w Krakowie: Krakow, Poland, 2009; pp. 1–11. [Google Scholar]
- Tomczyk-Wrona, I. Koniki Polskie-Modelowa populacja ochrony bioróżnorodności. Wiadomości Zootech. 2017, 5, 98–103. [Google Scholar]
- Jaworski, Z.; Siemieniuch, M.; Jastrzębska, E.; Wojciechowska, P. Zróżnicowanie Genetyczne Populacji Koników Polskich. In Proceedings of the LXXXIV Zjazd Naukowy Polskiego Towarzystwa Zootechnicznego w Szczecinie “Osiągniecia i perspektywy zootechniki w aspekcie zrównoważonego rolnictwa i ochrony środowiska”, Szczecin, Poland, 18–20 September 2019; p. 123. [Google Scholar]
- Chodkiewicz, A. Advantages and disadvantages of Polish primitive horse grazing on valuable nature areas—A review. Glob. Ecol. Conserv. 2020, 21, e00879. [Google Scholar] [CrossRef]
- Siemieniuch, M.J.; Jaworski, Z. A 17-year survey of reproductive efficiency in Polish Konik horses. Acta Sci. Pol. Zootech. 2018, 17, 31–40. [Google Scholar] [CrossRef]
- Jaworska, J.; Jaworski, Z.; McDonnell, S.M.; Gorecka-Bruzda, A. Harem stallion changes are not associated with diminished reproductive performance of females in semi-feral Konik polski horses (Equus caballus). Theriogenol. J. 2020, 151, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Zabek, T.; Nogaj, A.; Radko, A.; Nogaj, J.; Słota, E. Genetic variation of Polish endangered Biłgoraj horses and two common horse breeds in microsatellite loci. J. Appl. Genet. 2005, 46, 299–305. [Google Scholar] [PubMed]
- Vostry, L.; Vostra-Vydrova, H.; Hofmanova, B.; Vesela, Z.; Majzlik, I. Genetic diversity in Czech Haflinger horses. Poljoprivreda/Agriculture 2015, 21, 163–165. [Google Scholar] [CrossRef]
- Mackowski, M.; Mucha, S.; Cholewinski, G.; Cieslak, J. Genetic diversity in Hucul and Polish primitive horse breeds. Arch. Anim. Breed. 2015, 58, 23–31. [Google Scholar] [CrossRef]
- Castaneda, C.; Juras, R.; Khanshour, A.; Randlaht, I.; Wallner, B.; Rigler, D.; Lindgren, G.; Raudsepp, T.; Cothran, E.G. Population Genetic Analysis of the Estonian Native Horse Suggests Diverse and Distinct Genetics, Ancient Origin and Contribution from Unique Patrilines. Genes 2019, 10, 629. [Google Scholar] [CrossRef]
- Fornal, A.; Radko, A. Ocena polimorfizmu markerów mikrosatelitarnych DNA u koników polskich. Episteme Czas. Nauk. 2012, 14, 287–292. [Google Scholar]
- Solis, A.; Jugo, B.M.; Mériaux, J.C.; Iriondo, M.; Mazón, L.I.; Aguirre, A.I.; Vicario, A.; Estomba, A. Genetic diversity within and among four South European native horse breeds based on microsatellite DNA analysis: Implications for conservation. J. Hered. 2005, 96, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT, a program to estimate and test gene diversities and fixation indices (version 2.9. 3). Department of Ecology and Evolution, University of Lausanne, Switzerland. J. Hered. 2001, 149, 507–526. [Google Scholar]
- Pritchard, J.K.; Stephens, P.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Earl, D.A.; VonHoldt, B.M. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Paradis, E. Pegas: An R package for population genetics with an integrated-modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef]
- Huston, K.A. Statistical Analysis of STR Data. Profiles DNA 1998, 1, 14–15. [Google Scholar]
- Nei, M. Molecular Evolutionary Genetics; New York Columbia University Press: New York, NY, USA, 1987; p. 512. [Google Scholar]
- Ling, Y.; Cheng, Y.; Wang, Y.; Guan, W.; Han, J.; Fu, B.; He, X.; Pu, Y. Genetic diversity of 23 Chinese indigenous horse breeds revealed by microsatellite markers. Biodivers. Sci. 2009, 17, 240–247. [Google Scholar]
- Nei, M.; Roychoudhury, A.K. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- May-Davis, S.; Brown, W.Y.; Shorter, K.; Vermeulen, Z.; Butler, R.; Koekkoek, M. A novel non-invasive selection criterion for the preservation of primitive dutch konik horses. Animals 2018, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Brem, G.; Müller, M.; Achmann, R. Fixed nucleotide differences on the Y chromosome indicate clear divergence between Equus przewalskii and Equus caballus. Anim. Genet. 2003, 34, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Vogl, C.; Shukla, P.; Burgstaller, J.P.; Druml, T.; Brem, G. Identification of Genetic Variation on the Horse Y Chromosome and the Tracing of Male Founder Lineages in Modern Breeds. PLoS ONE 2013, 8, e60015. [Google Scholar] [CrossRef]
- Rendo, F.; Iriondo, M.; Manzano, C.; Estonba, A. Effectes of a 10-years conservation programme on the genetic diversity of the Pottoka pony-new clues regarding their origin. J. Anim. Breed Genet. 2012, 129, 234–243. [Google Scholar] [CrossRef]
- Lopes, M.S.; Mendonca, D.; Rojer, H.; Cabral, V.; Bettencourt, S.X.; da Câmara Machado, A. Morphological and genetic characterization of an emerging Azorean horse breed: The Terceira Pony. Front. Genet. 2015, 5, 1–7. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Paternal Lineage | N | Na | Na Freq. ≥ 5% | Ae | I | No. Private Alleles | He |
|---|---|---|---|---|---|---|---|
| Wicek | 48 | 6.824 | 4.647 | 3.824 | 1.476 | 0.412 | 0.693 |
| Myszak | 12 | 5.059 | 4.000 | 3.306 | 1.302 | 0.000 | 0.657 |
| Glejt I | 35 | 6.294 | 4.588 | 3.875 | 1.453 | 0.059 | 0.700 |
| Goraj | 32 | 6.118 | 4.176 | 3.627 | 1.396 | 0.000 | 0.678 |
| Chochlik | 36 | 6.471 | 4.765 | 3.730 | 1.449 | 0.000 | 0.689 |
| Liliput | 33 | 6.176 | 4.588 | 3.715 | 1.429 | 0.059 | 0.688 |
| FIT | FST | FIS | |
|---|---|---|---|
| AHT4 | −0.0096 | 0.0213 | −0.0316 |
| AHT5 | 0.0162 | 0.0373 | −0.0219 |
| ASB2 | −0.0100 | 0.0113 | −0.0215 |
| HMS2 | −0.0191 | 0.0053 | −0.0245 |
| HMS3 | 0.0231 | 0.0357 | −0.0130 |
| HMS6 | −0.0185 | 0.0269 | −0.0466 |
| HMS7 | 0.0613 | 0.0673 | −0.0065 |
| HTG10 | −0.0061 | 0.0211 | −0.0278 |
| HTG4 | 0.0342 | 0.0099 | 0.0246 |
| HTG6 | 0.0000 | 0.0526 | −0.0555 |
| HTG7 | 0.0105 | 0.0244 | −0.0143 |
| VHL20 | 0.0261 | 0.0438 | −0.0185 |
| ASB17 | 0.0142 | 0.0166 | −0.0024 |
| ASB23 | 0.0449 | 0.0262 | 0.0193 |
| CA425 | 0.1052 | 0.0756 | 0.0319 |
| HMS1 | 0.0305 | 0.0362 | −0.0059 |
| LEX3 | 0.9562 | 0.0079 | 0.9559 |
| Mean | 0.0741 | 0.0305 | −0.0134 |
| Ho | He | PD | PE | PIC | PE1 | PE2 | PID | |
|---|---|---|---|---|---|---|---|---|
| AHT4 | 0.7602 | 0.7482 | 0.8914 | 0.5274 | 0.7088 | 0.3486 | 0.5261 | 0.1028 |
| AHT5 | 0.7908 | 0.7964 | 0.9280 | 0.5821 | 0.7672 | 0.4240 | 0.6020 | 0.0706 |
| ASB2 | 0.8010 | 0.7895 | 0.9303 | 0.6009 | 0.7670 | 0.4333 | 0.6129 | 0.0669 |
| HMS2 | 0.7398 | 0.7234 | 0.8819 | 0.4925 | 0.6857 | 0.3207 | 0.5013 | 0.1142 |
| HMS3 | 0.7653 | 0.7763 | 0.9101 | 0.5363 | 0.7431 | 0.3902 | 0.5694 | 0.0832 |
| HMS6 | 0.8163 | 0.7956 | 0.9222 | 0.6297 | 0.7638 | 0.4146 | 0.5929 | 0.0735 |
| HMS7 | 0.6684 | 0.7015 | 0.8654 | 0.3810 | 0.6519 | 0.2914 | 0.4610 | 0.1387 |
| HTG10 | 0.8112 | 0.8012 | 0.9326 | 0.6200 | 0.7747 | 0.4393 | 0.6160 | 0.0661 |
| HTG4 | 0.4541 | 0.4681 | 0.6885 | 0.1504 | 0.4421 | 0.1198 | 0.2785 | 0.3090 |
| HTG6 | 0.1684 | 0.1663 | 0.3011 | 0.0217 | 0.1593 | 0.0138 | 0.0843 | 0.7020 |
| HTG7 | 0.6173 | 0.6195 | 0.7915 | 0.3122 | 0.5522 | 0.1994 | 0.3475 | 0.2121 |
| VHL20 | 0.7806 | 0.7931 | 0.9267 | 0.5636 | 0.7683 | 0.4344 | 0.6121 | 0.0676 |
| ASB17 | 0.7857 | 0.7926 | 0.9197 | 0.5728 | 0.7662 | 0.4311 | 0.6081 | 0.0694 |
| ASB23 | 0.7092 | 0.7371 | 0.8827 | 0.4426 | 0.6954 | 0.3333 | 0.5094 | 0.1108 |
| CA425 | 0.6480 | 0.7122 | 0.8793 | 0.3524 | 0.6813 | 0.3198 | 0.5046 | 0.1137 |
| HMS1 | 0.7296 | 0.7457 | 0.8956 | 0.4755 | 0.7124 | 0.3541 | 0.5358 | 0.0979 |
| LEX3 | 0.0357 | 0.8110 | 0.8230 | 0.0012 | 0.7858 | 0.4552 | 0.6307 | 0.0609 |
| 0.6519 * | 0.7046 * | 0.9999 ** | 0.9999 ** | 0.6721 * | 0.9992 ** | 0.9999 ** | 5.04 × 10−17 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fornal, A.; Kowalska, K.; Zabek, T.; Piestrzynska-Kajtoch, A.; Musiał, A.D.; Ropka-Molik, K. Genetic Diversity and Population Structure of Polish Konik Horse Based on Individuals from All the Male Founder Lines and Microsatellite Markers. Animals 2020, 10, 1569. https://doi.org/10.3390/ani10091569
Fornal A, Kowalska K, Zabek T, Piestrzynska-Kajtoch A, Musiał AD, Ropka-Molik K. Genetic Diversity and Population Structure of Polish Konik Horse Based on Individuals from All the Male Founder Lines and Microsatellite Markers. Animals. 2020; 10(9):1569. https://doi.org/10.3390/ani10091569
Chicago/Turabian StyleFornal, Agnieszka, Katarzyna Kowalska, Tomasz Zabek, Agata Piestrzynska-Kajtoch, Adrianna D. Musiał, and Katarzyna Ropka-Molik. 2020. "Genetic Diversity and Population Structure of Polish Konik Horse Based on Individuals from All the Male Founder Lines and Microsatellite Markers" Animals 10, no. 9: 1569. https://doi.org/10.3390/ani10091569
APA StyleFornal, A., Kowalska, K., Zabek, T., Piestrzynska-Kajtoch, A., Musiał, A. D., & Ropka-Molik, K. (2020). Genetic Diversity and Population Structure of Polish Konik Horse Based on Individuals from All the Male Founder Lines and Microsatellite Markers. Animals, 10(9), 1569. https://doi.org/10.3390/ani10091569