Over-Winter Survival and Nest Site Selection of the West-European Hedgehog (Erinaceus europaeus) in Arable Dominated Landscapes

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Nesting Behaviour
2.2. Patterns of Survival
2.3. Body Mass Changes
2.4. Data Analysis
3. Results
3.1. Nesting Behaviour
3.2. Patterns of Survival
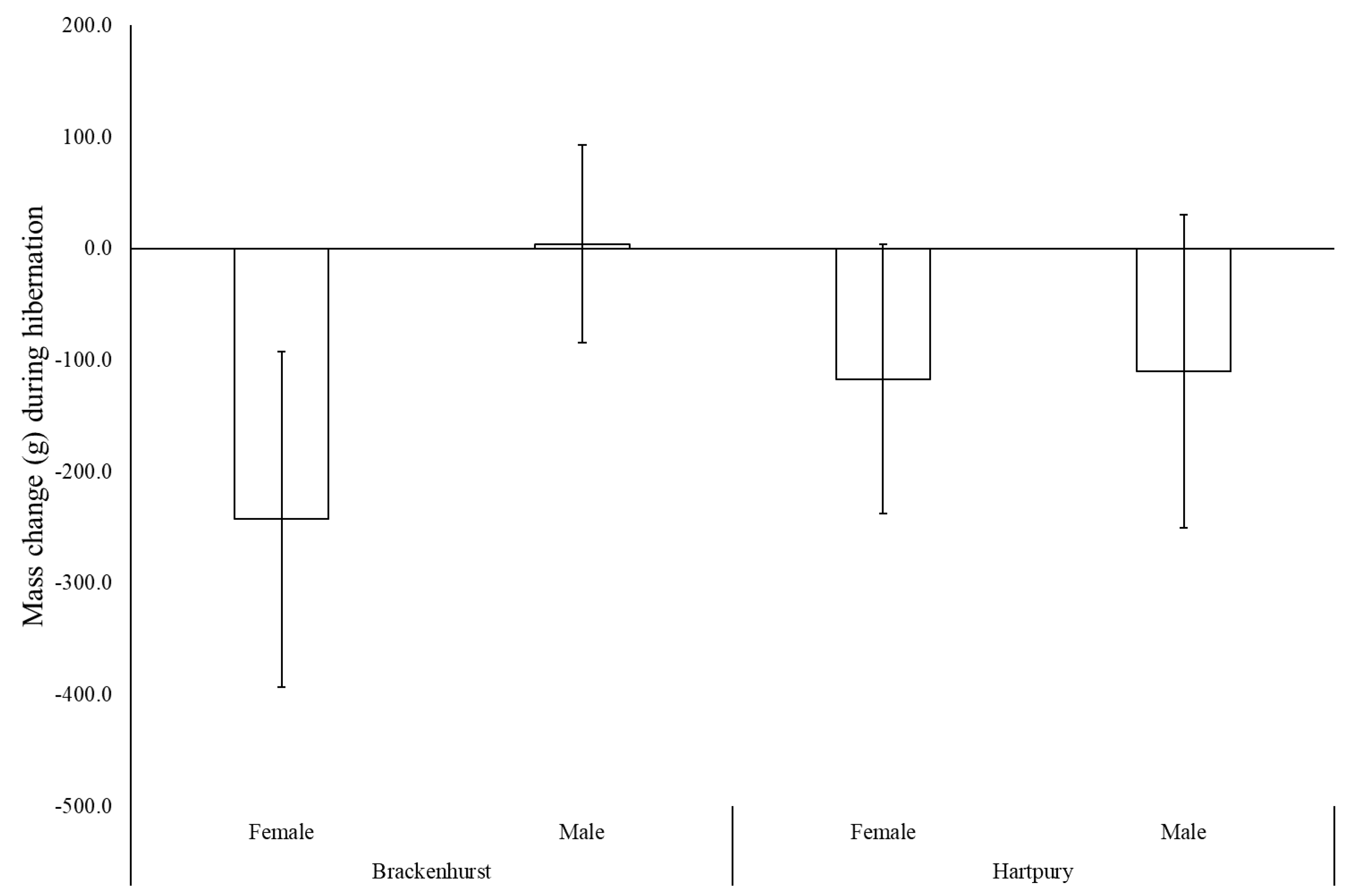
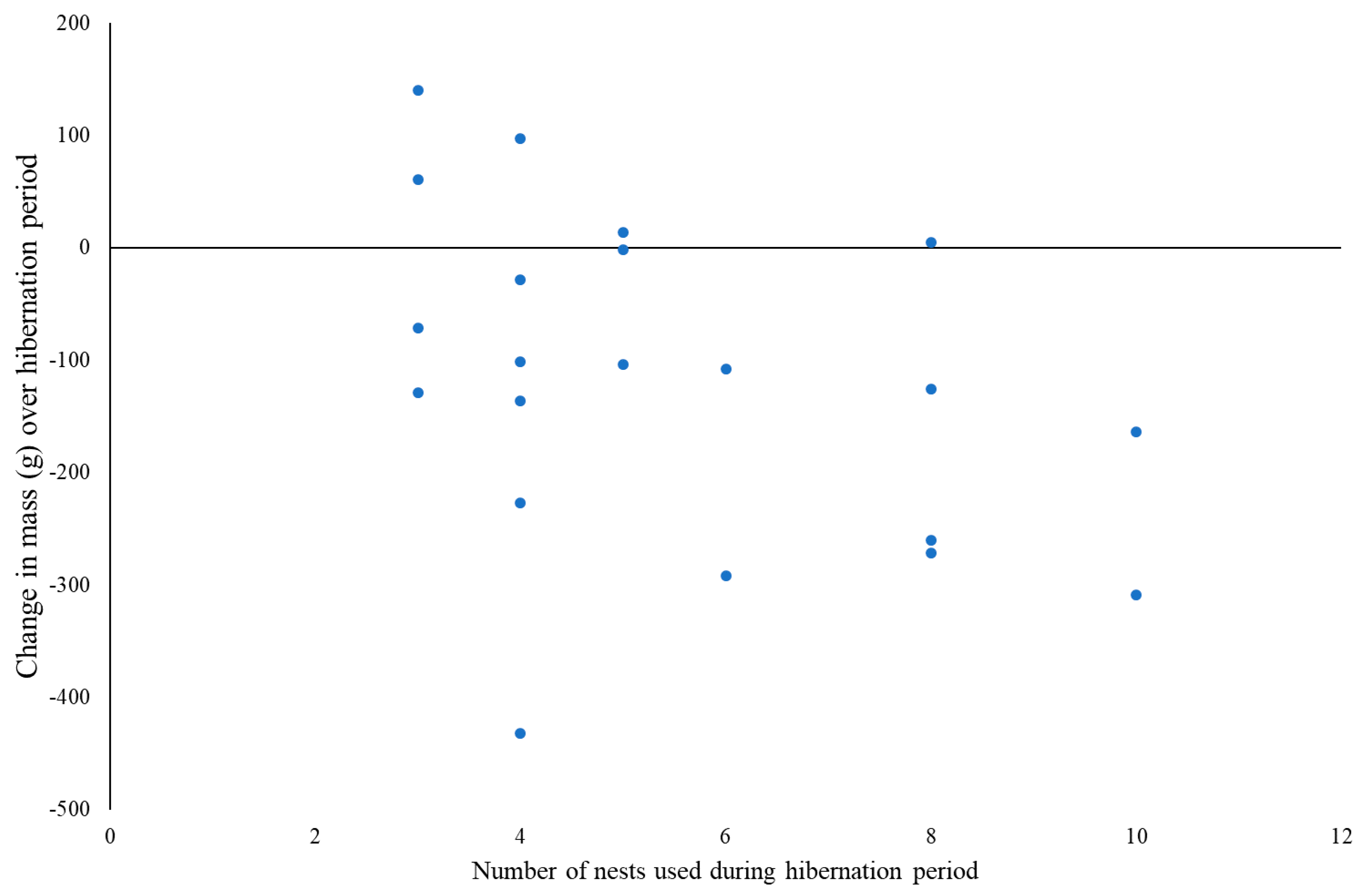
3.3. Body Mass Changes
4. Discussion
4.1. Change in Body Mass
4.2. Nesting Behaviour
4.3. Over-Winter Survival
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Firbank, L.G.; Petit, S.; Smart, S.; Blain, A.; Fuller, R.J. Assessing the impacts of agricultural intensification on biodiversity: A British perspective. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 777–787. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Tuck, S.L.; Winqvist, C.; Mota, F.; Ahnström, J.; Turnbull, L.A.; Bengtsson, J. Land-use intensity and the effects of organic farming on biodiversity: A hierarchical meta-analysis. J. Appl. Ecol. 2014, 51, 746–755. [Google Scholar] [CrossRef]
- Veach, V.; Moilanen, A.; Di Minin, E. Threats from urban expansion, agricultural transformation and forest loss on global conservation priority areas. PLoS ONE 2017, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zabel, F.; Delzeit, R.; Schneider, J.M.; Seppelt, R.; Mauser, W.; Václavík, T. Global impacts of future cropland expansion and intensification on agricultural markets and biodiversity. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Firbank, L.G.; Attwood, S.; Eory, V.; Gadanaski, Y.; Lynch, J.M.; Sonnino, R.; Takahashi, T. Grand challenges in sustainable intensification and ecosystem services. Front. Sustain. Food Syst. 2018, 2, 7. [Google Scholar] [CrossRef]
- Ellis, E.C.; Goldewijk, K.K.; Siebert, S.; Lightman, D.; Ramankutty, N. Anthropogenic transformation of the biomes, 1700 to 2000. Glob. Ecol. Biogeogr. 2010, 19, 589–606. [Google Scholar] [CrossRef]
- Crooks, K.R.; Burdett, C.L.; Theobald, D.M.; Rondinini, C.; Boitani, L. Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2642–2651. [Google Scholar] [CrossRef]
- Robinson, T.P.; Wint, G.R.; Conchedda, G.; Van Boeckel, T.P.; Ercoli, V.; Palamara, E.; Cinardi, G.; D’Aietti, L.; Hay, S.I.; Gilbert, M. Mapping the global distribution of livestock. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Wiethoelter, A.K.; Beltrán-Alcrudo, D.; Kock, R.; Mor, S.M. Global trends in infectious diseases at the wildlife-livestock interface. Proc. Natl. Acad. Sci. USA 2015, 112, 9662–9667. [Google Scholar] [CrossRef] [PubMed]
- Gordon, I.J. Review: Livestock production increasingly influences wildlife across the globe. Animal 2018, 12, S372–S382. [Google Scholar] [CrossRef] [PubMed]
- Howell, H.J.; Mothes, C.C.; Clements, S.L.; Catania, S.V.; Rothermel, B.B.; Searcy, C.A. Amphibian responses to livestock use of wetlands: New empirical data and a global review. Ecol. Appl. 2019, 29, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Öllerer, K.; Varga, A.; Kirby, K.; Demeter, L.; Biró, M.; Bölöni, J.; Molnár, Z. Beyond the obvious impact of domestic livestock grazing on temperate forest vegetation—A global review. Biol. Conserv. 2019, 237, 209–219. [Google Scholar] [CrossRef]
- Baker, P.J.; Boitani, L.; Harris, S.; Saunders, G.; White, P.C.L. Terrestrial carnivores and human food production: Impact and management. Mamm. Rev. 2008, 38, 123–166. [Google Scholar] [CrossRef]
- Eklund, A.; López-Bao, J.V.; Tourani, M.; Chapron, G.; Frank, J. Limited evidence on the effectiveness of interventions to reduce livestock predation by large carnivores. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- van Eeden, L.M.; Eklund, A.; Miller, J.R.B.; López-Bao, J.V.; Chapron, G.; Cejtin, M.R.; Crowther, M.S.; Dickman, C.R.; Frank, J.; Krofel, M.; et al. Carnivore conservation needs evidence-based livestock protection. PLoS Biol. 2018, 16, 1–8. [Google Scholar] [CrossRef]
- van Eeden, L.M.; Crowther, M.S.; Dickman, C.R.; Macdonald, D.W.; Ripple, W.J.; Ritchie, E.G.; Newsome, T.M. Managing conflict between large carnivores and livestock. Conserv. Biol. 2018, 32, 26–34. [Google Scholar] [CrossRef]
- Garcês, A.; Pires, I.; Rodrigues, P. Teratological effects of pesticides in vertebrates: A review. J. Environ. Sci. Heal.Part. B Pestic. Food Contam. Agric. Wastes 2020, 55, 75–89. [Google Scholar] [CrossRef]
- Brown, D.D.; Kays, R.; Wikelski, M.; Wilson, R.; Klimley, A. Observing the unwatchable through acceleration logging of animal behavior. Anim. Biotelemetry 2013, 1. [Google Scholar] [CrossRef]
- Kharouba, H.M.; Ehrlén, J.; Gelman, A.; Bolmgren, K.; Allen, J.M.; Travers, S.E.; Wolkovich, E.M. Global shifts in the phenological synchrony of species interactions over recent decades. Proc. Natl. Acad. Sci. USA 2018, 115, 5211–5216. [Google Scholar] [CrossRef] [PubMed]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of recent declines. Science 2010, 1164, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The impact of conservation on the status of the world’s vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Di Marco, M.; Boitani, L.; Mallon, D.; Hoffmann, M.; Iacucci, A.; Meijaard, E.; Visconti, P.; Schipper, J.; Rondinini, C. A Retrospective evaluation of the global decline of carnivores and ungulates. Conserv. Biol. 2014, 28, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, S.L.; Fuller, R.A.; Brooks, T.M.; Watson, J.E.M. Biodiversity: The ravages of guns, nets and bulldozers. Nature 2016, 536, 143–145. [Google Scholar] [CrossRef]
- Dudley, N.; Alexander, S. Agriculture and biodiversity: A review. Biodiversity 2017, 18, 45–49. [Google Scholar] [CrossRef]
- Stanton, R.L.; Morrissey, C.A.; Clark, R.G. Analysis of trends and agricultural drivers of farmland bird declines in North America: A review. Agric. Ecosyst. Environ. 2018, 254, 244–254. [Google Scholar] [CrossRef]
- Clout, M.N.; Russell, J.C. The invasion ecology of mammals: A global perspective. Wildl. Res. 2007, 35, 180–184. [Google Scholar] [CrossRef]
- Long, J.L. Introduced Mammals of the World: Their History, Distribution and Influence; CSIRO Publishing: Victoria, Australia, 2003. [Google Scholar]
- Inouye, D.W.; Barr, B.; Armitage, K.B.; Inouye, B.D. Climate change is affecting altitudinal migrants and hibernating species. Proc. Natl. Acad. Sci. USA 2000, 97, 1630–1633. [Google Scholar] [CrossRef]
- Lane, J.E. Evolutionary Ecology of Mammalian Hibernation Phenology. In Living in a Seasonal World; Ruf, T., Bieber, C., Arnold, W., Millesi, E., Eds.; Springer: Berlin, Germany, 2012; pp. 51–61. ISBN 9783642286780. [Google Scholar]
- Lane, J.E.; Kruuk, L.E.B.; Charmantier, A.; Murie, J.O.; Dobson, F.S. Delayed phenology and reduced fitness associated with climate change in a wild hibernator. Nature 2012, 489, 554–557. [Google Scholar] [CrossRef]
- Geiser, F. Hibernation. Curr. Biol. 2013, 23, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Geiser, F. Hibernation: Endotherms. eLS 2011, 1. [Google Scholar] [CrossRef]
- Staples, J.F. Metabolic suppression in mammalian hibernation: The role of mitochondria. J. Exp. Biol. 2014, 217, 2032–2036. [Google Scholar] [CrossRef] [PubMed]
- Turbill, C.; Bieber, C.; Ruf, T. Hibernation is associated with increased survival and the evolution of slow life histories among mammals. Proc. R. Soc. B Biol. Sci. 2011, 278, 3355–3363. [Google Scholar] [CrossRef] [PubMed]
- Morris, P.A.; Reeve, N.J. Hedgehog Erinaceus europaeus. In Mammals of the British Isles: Handbook; Harris, S.J., Yalden, D.W., Eds.; The Mammal Society: Southampton, UK, 2008; pp. 241–249. [Google Scholar]
- Burton, M. The Hedgehog; Andre Deutsch: London, UK, 1969. [Google Scholar]
- Tapper, S.S. Game Heritage: An Ecological Review from Shooting and Gamekeeping Records; Game Conservancy Ltd.: Fordingbridge, Hampshire, UK, 1992. [Google Scholar]
- Arnold, H.R. Atlas of Mammals of Britain; HMSO: London, UK, 1993. [Google Scholar]
- Lovegrove, R. Silent Fields: The Long Decline of a Nation’s Wildlife; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Hof, A.R.; Bright, P.W. Quantifying the long-term decline of the West European hedgehog in England by subsampling citizen-science datasets. Eur. J. Wildl. Res. 2016, 62, 407–413. [Google Scholar] [CrossRef]
- Mathews, F.; Kubasiewicz, L.M.; Gurnell, J.; Harrower, C.A.; McDonald, R.A.; Shore, R.F. A Review of the Population and Conservation Status of British Mammals. A report by the Mammal. Society under contract to Natural England, Natural Resources Wales and Scottish Natural Heritage; Natural England: Peterborough, UK, 2018; ISBN 9781783544943. [Google Scholar]
- Williams, B.M.; Baker, P.J.; Thomas, E.; Wilson, G.J.; Judge, J.; Yarnell, R.W. Reduced occupancy of hedgehogs (Erinaceus europaeus) in rural England and Wales: The influence of habitat and an asymmetric intra-guild predator. Sci. Rep. 2018, 8, 12156. [Google Scholar] [CrossRef]
- Young, R.P.; Davison, J.; Trewby, I.D.; Wilson, G.J.; Delahay, R.J.; Doncaster, C.P. Abundance of hedgehogs (Erinaceus europaeus) in relation to the density and distribution of badgers (Meles meles). J. Zool. 2006, 269, 349–356. [Google Scholar] [CrossRef]
- Parrott, D.; Etherington, T.R.; Dendy, J. A geographically extensive survey of hedgehogs (Erinaceus europaeus) in England. Eur. J. Wildl. Res. 2014, 60, 399–403. [Google Scholar] [CrossRef]
- Pettett, C.E.; Moorhouse, T.P.; Johnson, P.J.; Macdonald, D.W. Factors affecting hedgehog (Erinaceus europaeus) attraction to rural villages in arable landscapes. Eur. J. Wildl. Res. 2017, 63, 54. [Google Scholar] [CrossRef]
- Hubert, P.; Julliard, R.; Biagianti, S.; Poulle, M.L. Ecological factors driving the higher hedgehog (Erinaceus europeaus) density in an urban area compared to the adjacent rural area. Landsc. Urban. Plan. 2011, 103, 34–43. [Google Scholar] [CrossRef]
- Van de Poel, J.L.; Dekker, J.; Langevelde, F.V. Dutch hedgehogs Erinaceus europaeus are nowadays mainly found in urban areas, possibly due to the negative effects of badgers Meles meles. Wildl. Biol. 2015, 21, 51–55. [Google Scholar] [CrossRef]
- Robinson, R.A.; Sutherland, W.J. Post-war changes in arable farming and biodiversity in Great Britain. J. Appl. Ecol. 2002, 39, 157–176. [Google Scholar] [CrossRef]
- Carey, P.D.; Wallis, S.M.; Emmett, B.; Maskell, L.C.; Murphy, J.; Norton, L.R.; Simpson, I.C.; Smart, S.M. Countryside Survey: UK Headline Messages from 2007; Centre for Ecology & Hydrology: Wallingford, UK, 2008. [Google Scholar]
- Wright, J. A Natural History of the Hedgerow and Ditches, Dykes and Dry Stone Walls; Profile Books Ltd.: London, UK, 2016. [Google Scholar]
- Dover, J.W. The Ecology of Hedgerows and Field Margins; Routledge: Abingdon, UK, 2019. [Google Scholar]
- Hof, A.R.; Bright, P.W. The value of agri-environment schemes for macro-invertebrate feeders: Hedgehogs on arable farms in Britain. Anim. Conserv. 2010, 13, 467–473. [Google Scholar] [CrossRef]
- Moorhouse, T.P.; Palmer, S.C.F.; Travis, J.M.J.; Macdonald, D.W. Hugging the hedges: Might agri-environment manipulations affect landscape permeability for hedgehogs? Biol. Conserv. 2014, 176, 109–116. [Google Scholar] [CrossRef]
- Hof, A.R.; Snellenberg, J.; Bright, P.W. Food or fear? Predation risk mediates edge refuging in an insectivorous mammal. Anim. Behav. 2012, 83, 1099–1106. [Google Scholar] [CrossRef]
- Judge, J.; Wilson, G.J.; Macarthur, R.; Delahay, R.J.; McDonald, R.A. Density and abundance of badger social groups in England and Wales in 2011-2013. Sci. Rep. 2014, 4, 3809. [Google Scholar] [CrossRef]
- Judge, J.; Wilson, G.J.; Macarthur, R.; McDonald, R.A.; Delahay, R.J. Abundance of badgers (Meles meles) in England and Wales. Sci. Rep. 2017, 7, 276. [Google Scholar] [CrossRef]
- Trewby, I.D.; Young, R.P.; McDonald, R.A.; Wilson, G.J.; Davison, J.; Walker, N.; Robertson, P.A.; Doncaster, C.P.; Delahay, R.J. Impacts of removing badgers on localised counts of hedgehogs. PLoS ONE 2014, 9, 2–5. [Google Scholar] [CrossRef]
- Morris, P.A. Hedgehog; HarperCollins: London, UK, 2018. [Google Scholar]
- Morris, P.A. Winter nests of the hedgehog. Oecologia 1973, 11, 299–313. [Google Scholar] [CrossRef]
- Kristiansson, H. Population variables and causes of mortality in a hedgehog (Erinaceus europaeus) population in Southern Sweden. J. Zool. 1990, 220, 391–404. [Google Scholar] [CrossRef]
- Morris, P.A. An estimate of the minimum body weight necessary for hedgehogs (Erinaceus europaeus) to survive hibernation. J. Zool. 1984, 203, 291–294. [Google Scholar]
- Yarnell, R.W.; Surgey, J.; Grogan, A.; Thompson, R.; Davies, K.; Kimbrough, C.; Scott, D.M. Should rehabilitated hedgehogs be released in winter? A comparison of survival, nest use and weight change in wild and rescued animals. Eur. J. Wildl. Res. 2019, 65. [Google Scholar] [CrossRef]
- Natural England Entry Level Stewardship; Natural England: Sheffield, UK, 2013; ISBN 978-1-84754-239-7.
- Schaus, J.; Uzal, A.; Gentle, L.K.; Baker, P.J.; Bearman-Brown, L.; Bullion, S.; Gazzard, A.; Lockwood, H.; North, A.; Reader, T.; et al. Application of the Random Encounter Model in citizen science projects to monitor animal densities. Remote Sens. Ecol. Conserv. 2020. [Google Scholar] [CrossRef]
- Haigh, A.; O’Riordan, R.M.; Butler, F. Nesting behaviour and seasonal body mass changes in a rural Irish population of the Western hedgehog (Erinaceus europaeus). Acta Theriol. 2012, 57, 321–331. [Google Scholar] [CrossRef]
- Rautio, A.; Valtonen, A.; Auttila, M.; Kunnasranta, M. Nesting patterns of European hedgehogs (Erinaceus europaeus) under northern conditions. Acta Theriol. 2014, 59, 173–181. [Google Scholar] [CrossRef]
- Morris, P.A. Hedgehogs, 4th ed.; Whittet Books: Stansted, UK, 2014. [Google Scholar]
- McClean, S.A.; Rumble, M.A.; King, R.M.; Baker, W.L. Evaluation of Resource Selection Methods with Different Definitions of Availability. J. Wildl. Manag. 1998, 62, 793. [Google Scholar] [CrossRef]
- Uzal, A.; Walls, S.; Stillman, R.A.; Diaz, A. Sika deer distribution and habitat selection: The influence of the availability and distribution of food, cover, and threats. Eur. J. Wildl. Res. 2013, 59, 563–572. [Google Scholar] [CrossRef]
- Environmental Systems Resource Institute. ArcGIS 2015; ESRI: Redlands, CA, USA, 2015. [Google Scholar]
- Manly, B.F.L.; McDonald, L.; Thomas, D.L.; McDonald, T.L.; Erickson, W.P. Resource Selection by Animals: Statistical Design and Analysis for Field Studies; Springer: London, UK, 2007. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Multi-Model Inference: A Practical Information-Theoretic Approach; Springer: New York City, NY, USA, 2002. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: London, UK, 2009. [Google Scholar]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457. [Google Scholar] [CrossRef]
- Pollock, K.H.; Winterstein, S.R.; Bunck, C.M.; Curtis, P.D. Survival analysis in telemetry studies: The staggered entry design. J. Wildl. Manage. 1989, 53, 7–15. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team R: Vienna, Austria, 2016. [Google Scholar]
- Bates, D.; Machler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. Package “MuMIn”; R Package Version 1(6); CRAN: 2019. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 19 August 2020).
- RSPCA. RSPCA Wildlife Rehabilitation Protocol: Hedgehogs; RSPCA: Southwater, UK, 2013. [Google Scholar]
- Jensen, A.B. Overwintering of European hedgehogs Erinaceus europaeus in a Danish rural area. Acta Theriol. 2004, 49, 145–155. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Berg, T.B.; Dabelsteen, T.; Jones, O.R. The ecology of suburban juvenile European hedgehogs (Erinaceus europaeus) in Denmark. Ecol. Evol. 2019, 9, 13174–13187. [Google Scholar] [CrossRef]
- Tähti, H.; Soivio, A. Respiratory and circulatory differences between induced and spontaneous arousals in hibernating hedgehogs (Erinaceus europaeus L.). Ann. Zool. Fenn. 1977, 14, 198–203. [Google Scholar]
- Walhovd, H. Partial arousals from hibernation in hedgehogs in outdoor hibernacula. Oecologia 1979, 153, 141–153. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. The impact of grassy field margins on macro-invertebrate abundance in adjacent arable fields. Agric. Ecosyst. Environ. 2010, 139, 280–283. [Google Scholar] [CrossRef]
- Pettett, C.E.; Johnson, P.J.; Moorhouse, T.P.; Hambly, C.; Speakman, J.R.; Macdonald, D.W. Daily energy expenditure in the face of predation: Hedgehog energetics in rural landscapes. J. Exp. Biol. 2017, 220, 460–468. [Google Scholar] [CrossRef]
- Riber, A. Habitat use and behaviour of European hedgehog Erinaceus europaeus in a Danish rural area. Acta Theriol. 2006, 51, 363–371. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Armstrong-Brown, S.; Armsworth, P.R.; Brereton, T.; Brickland, J.; Campbell, C.D.; Chamberlain, D.E.; Cooke, A.I.; Dulvy, N.K.; Dusic, N.R.; et al. The identification of 100 ecological questions of high policy relevance in the UK. J. Appl. Ecol. 2006, 43, 617–627. [Google Scholar] [CrossRef]
- Cornulier, T.; Robinson, R.A.; Elston, D.; Lambin, X.; Sutherland, W.J.; Benton, T.G. Bayesian reconstitution of environmental change from disparate historical records: Hedgerow loss and farmland bird declines. Methods Ecol. Evol. 2011, 2, 86–94. [Google Scholar] [CrossRef]
- Harris, S.J.; Morris, P.A.; Wray, S.; Yalden, D.W. A Review of British Mammals: Population Estimates and Conservation Status of British Mammals Other than Cetaceans; JNCC: Peterborough, UK, 1995; ISBN 1-873701-68-3. [Google Scholar]
- Bearman-Brown, L.E.; Wilson, L.E.; Evans, L.; Baker, P.J. Comparing non-invasive surveying techniques for elusive, nocturnal mammals: A case study of the West European hedgehog (Erinaceus europaeus). J. Vertebr. Biol. in press.
- Wilson, G.; Harris, S.J.; McLaren, G. Changes in the British Badger Population, 1988 to 1997; Kerenza J Ltd.: London, UK, 1997. [Google Scholar]
- Roper, T.J. Badger; HarperCollins: London, UK, 2010. [Google Scholar]
- Hopkins, J.J.; Kirby, K.J. Ecological change in British broadleaved woodland since 1947. Ibis 2007, 149, 29–40. [Google Scholar] [CrossRef]
- Amar, A.; Smith, K.W.; Butler, S.; Lindsell, J.A.; Hewson, C.M.; Fuller, R.J.; Charman, E.C. Recent patterns of change in vegetation structure and tree composition of British broadleaved woodland: Evidence from large-scale surveys. Forestry 2010, 83, 345–356. [Google Scholar] [CrossRef]
- Rautio, A.; Valtonen, A.; Kunnasranta, M. The effects of sex and season on home range in European hedgehogs at the northern edge of the species range. Ann. Zool. Fenn. 2013, 50, 107–123. [Google Scholar] [CrossRef]
- Glasby, L.; Yarnell, R.W. Evaluation of the performance and accuracy of Global Positioning System bug transmitters deployed on a small mammal. Eur. J. Wildl. Res. 2013, 59, 915–919. [Google Scholar] [CrossRef]
- Chmura, H.E.; Glass, T.W.; Williams, C.T. Biologging physiological and ecological responses to climatic variation: New tools for the climate change era. Front. Ecol. Evol. 2018, 6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brackenhurst | Hartpury | Total | |||
|---|---|---|---|---|---|
| 2015–2016 | 2016–2017 | 2015–2016 | 2016–2017 | ||
| No. captured & marked | 7 (4♀:3♂) | 3 (2♀:1♂) | 22 (12♀:10♂) | 8 (3♀: 5♂) | 40 (21♀:19♂) |
| No. radio-tagged | 7 (4♀:3♂) | 3 (2♀:1♂) | 18 (9♀:9♂) | 5 (3♀:2♂) | 33 (18♀:15♂) |
| No. tracked during hibernation | 7 (4♀:3♂) | 3 (2♀:1♂) | 7 (4♀:3♂) | 4 (2♀:2♂) | 21 (12♀:9♂) |
| Total no. of nests recorded (% accessible for recording composition) | 54 (59%) | 12 (100%) | 50 (66%) | 16 (75%) | 138 (65%) |
| No. of nocturnal locations recorded | 103 | 74 | 210 | 61 | 448 |
| No. of diurnal locations recorded | 408 | 114 | 360 | 146 | 1028 |
| Brackenhurst Model A | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Models (N = 64) | |||||||||
| Amenity grassland | Buildings | Hedgerows | Pastures | Roads | Woodland | Arable | AIC | ΔAIC | AICw |
| ✓ | Not included | ✓ | ✓ | ✓ | ✓ | 357.5 | 0.00 | 0.38 | |
| ✓ | Not included | ✓ | ✓ | ✓ | ✓ | ✓ | 358.2 | 0.75 | 0.26 |
| ✓ | Not included | ✓ | ✓ | ✓ | ✓ | 359.4 | 1.94 | 0.14 | |
| ✓ | Not included | ✓ | ✓ | ✓ | ✓ | 360.8 | 3.33 | 0.07 | |
| ✓ | Not included | ✓ | ✓ | ✓ | ✓ | 362.2 | 4.67 | 0.04 | |
| NULL | 491.2 | 134.00 | <0.01 | ||||||
| Brackenhurst Model B | |||||||||
| Models (N = 64) | |||||||||
| Amenity grassland | Buildings | Hedgerows | Pastures | Roads | Woodland | Arable | AIC | ΔAIC | AICw |
| Not included | ✓ | ✓ | ✓ | ✓ | ✓ | 350.2 | 0.00 | 0.41 | |
| Not included | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 351.1 | 0.90 | 0.26 |
| Not included | ✓ | ✓ | ✓ | ✓ | 352.1 | 1.89 | 0.16 | ||
| Not included | ✓ | ✓ | ✓ | ✓ | ✓ | 352.6 | 3.44 | 0.07 | |
| Not included | ✓ | ✓ | ✓ | ✓ | ✓ | 354.3 | 4.09 | 0.05 | |
| NULL | 491.2 | 141.00 | <0.01 | ||||||
| Hartpury | |||||||||
| Models (N = 128) | |||||||||
| Amenity grassland | Buildings | Hedgerows | Pastures | Roads | Woodland | Arable | AIC | ΔAIC | AICw |
| ✓ | ✓ | ✓ | ✓ | ✓ | 395.6 | 0.00 | 0.49 | ||
| ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 397.4 | 1.80 | 0.20 | |
| ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 397.6 | 2.04 | 0.18 | |
| ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 399.4 | 3.84 | 0.07 |
| ✓ | ✓ | ✓ | ✓ | 401.2 | 5.61 | 0.03 | |||
| NULL | 464.4 | 68.8 | <0.01 | ||||||
| Variable | Brackenhurst Model A (3 Best a-priori Models) | Brackenhurst Model B (3 Best a-priori Models) | Hartpury (2 Best a-priori Models) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SE | z | p-Value | Estimate | SE | z | p-Value | Estimate | SE | z | p-Value | |
| (Intercept) | −0.281 | 0.439 | 0.640 | 0.522 | −0.113 | 0.432 | 0.261 | 0.794 | −2.514 | 0.515 | 4.879 | <0.001 |
| Hedgerows | −0.013 | 0.006 | 2.000 | <0.05 | −0.013 | 0.006 | 2.000 | <0.05 | −0.008 | 0.003 | 3.204 | <0.01 |
| Pasture | 0.017 | 0.006 | 2.942 | <0.01 | 0.017 | 0.006 | 2.877 | <0.01 | 0.010 | 0.003 | 3.748 | <0.001 |
| Roads | −0.012 | 0.005 | 2.544 | <0.05 | −0.010 | 0.004 | 2.443 | <0.05 | −0.016 | 0.006 | 2.590 | <0.01 |
| Woodland | −0.020 | 0.003 | 5.919 | <0.001 | −0.020 | 0.003 | 5.607 | <0.001 | −0.013 | 0.003 | 3.774 | <0.001 |
| Arable | 0.002 | 0.002 | 1.127 | 0.260 | 0.002 | 0.002 | 1.062 | 0.288 | 0.005 | 0.001 | 3.436 | <0.001 |
| Buildings | Not included | −0.01 | 0.003 | 3.412 | <0.001 | 0.001 | 0.001 | 0.488 | 0.626 | |||
| Amenity grassland | −0.008 | 0.003 | 2.527 | <0.05 | Not included | Not included | ||||||
| Country | Habitat | Years Studied | Sample Size& Composition | Mass Loss Recordedover Winter | Minimum Weightto SurviveHibernation | Reference |
|---|---|---|---|---|---|---|
| England | Urban parkland | 1963–1968 | 105 | 25% | Recommends 450 g (550 g in more northern areas) | [63] |
| Denmark | Rural | 2001–2002 | 10 (5♀:5♂); (3A:7J) | 30.2 ± 7.1% (A) 22.1 ± 10.1% (J) | 513 g | [82] |
| Ireland | Rural | 2008–2009 | 8 (7A:1J) | 301 ± 3.9g (♀) (range: 15–38%) 108 ± 2.6g (♂) (range: 3–6%) | 475 g in Nov | [67] |
| Denmark | Suburban | 2014–2015 | 8 (8J) | 16 ± 2.9% (J) | - | [83] |
| England | Various | 2010–2014 | 55 (19♀:30♂:16?); (20A:35J) | 98.6 ± 35.6 g (♀) 160.8 ± 40.5 g (♂) 111.4 ± 33.0 g (A) 162.2 ± 43.3 g (J) 14.1 ± 3.1% (All animals) | Recommends >600 g for release, but one individual weighing 391 g survived release and hibernation | [64] |
| England | Various | 2015–2017 | 21 (11♀:10♂) | Site 1: 240 ± 150 g (25 ± 13%) (♀) Site 1: −4 ± 89 g (1 ± 9%) (♂) Site 2: 117 ± 121 g (14 ± 16%) (♀) Site 2: 110 ± 141 g (15± 19%) (♂) | - | Present study |
| Country | Habitat | Years Studied | Sample Size& Composition 1 | Duration of Hibernation(Days) | Number of Nests Used | Reference |
|---|---|---|---|---|---|---|
| England | Urban park | 1963–1967 | 167 nests | Not recorded | Mean occupation time = 1.4 months (range 0–6 months) | [61] |
| Denmark | Rural | 2001–2002 | 10 (3A:7J) | 197.7 ± 2.2 (A) 178.8 ± 13.1 (J) | 2.2 (range: 1–4) | [82] |
| Ireland | Rural | 2008–2010 | 8 (7A:1J) | 167.3 ± 10.5 (♀) 148.6 ± 10.2 (♂) 155.4 ± 9.0 (A) 157 (J) | 2.0 ± 0.6 (♀) 3.2 ± 0.6 (♂) 2.4 ± 0.7 (A) 5.0 (J) | [67] |
| Finland | Urban | 2004–2006 | 11 (11A) (5♀:6♂) | 223 ± 2.5 (♀) 224 ± 4.8 (♂) | 1.0 (♀) 1.0 (♂) | [68] |
| Denmark | Urban | 2014–2015 | 8 (8J) | 138.0 ± 5.6 (J) | 1.8 ± 0.14 (J) | [83] |
| England | Various | 2010–2014 | 55 (20A:35J); (19♀:30♂:16?) | Not recorded | 2.2 ± 0.5 (♀) 2 1.7 ± 0.4 (♂) 2 1.8 ± 0.4 (A) 2 2.6 ± 0.6 (J) 2 | [64] |
| England | Arable | 2015–2017 | 21A (12♀:9♂) | Not recorded | 5.8 ± 2.6 (♀) 5.0 ± 1.9 (♂) | Present study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bearman-Brown, L.E.; Baker, P.J.; Scott, D.; Uzal, A.; Evans, L.; Yarnell, R.W. Over-Winter Survival and Nest Site Selection of the West-European Hedgehog (Erinaceus europaeus) in Arable Dominated Landscapes. Animals 2020, 10, 1449. https://doi.org/10.3390/ani10091449
Bearman-Brown LE, Baker PJ, Scott D, Uzal A, Evans L, Yarnell RW. Over-Winter Survival and Nest Site Selection of the West-European Hedgehog (Erinaceus europaeus) in Arable Dominated Landscapes. Animals. 2020; 10(9):1449. https://doi.org/10.3390/ani10091449
Chicago/Turabian StyleBearman-Brown, Lucy E., Philip J. Baker, Dawn Scott, Antonio Uzal, Luke Evans, and Richard W. Yarnell. 2020. "Over-Winter Survival and Nest Site Selection of the West-European Hedgehog (Erinaceus europaeus) in Arable Dominated Landscapes" Animals 10, no. 9: 1449. https://doi.org/10.3390/ani10091449
APA StyleBearman-Brown, L. E., Baker, P. J., Scott, D., Uzal, A., Evans, L., & Yarnell, R. W. (2020). Over-Winter Survival and Nest Site Selection of the West-European Hedgehog (Erinaceus europaeus) in Arable Dominated Landscapes. Animals, 10(9), 1449. https://doi.org/10.3390/ani10091449