Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection, Library Construction, and miRNA Sequencing
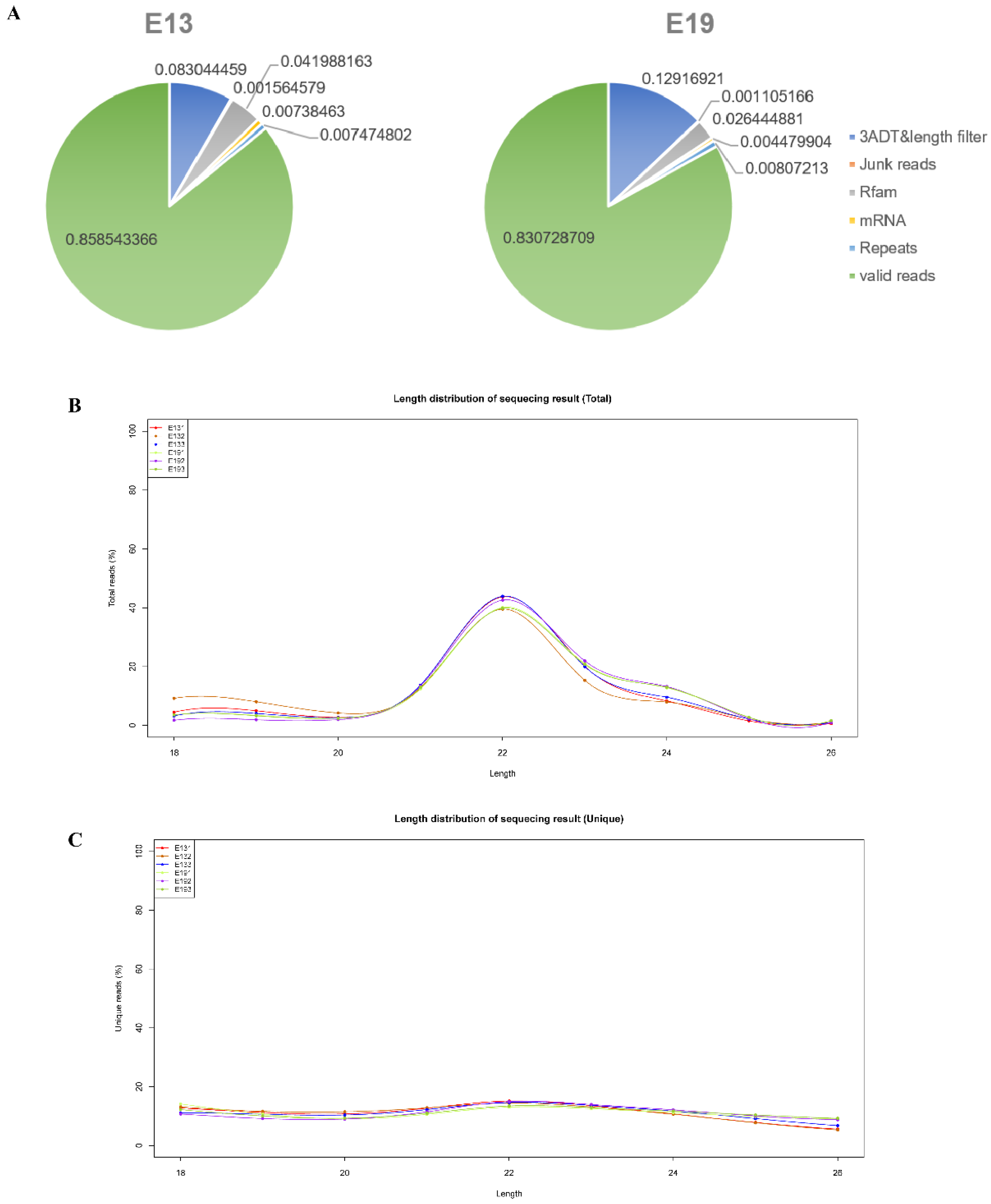
2.3. Sequencing Data Analysis
2.4. Naming and Classification of miRNAs
2.5. Target Gene Prediction and Enrichment Analysis of the Differentially Expressed miRNAs
2.6. Oligo Synthesis and qRT-PCR Validation
2.7. Cell Culture and Tranfection
2.8. Dual-Luciferase Reporter Assay
3. Results
3.1. Overview of Sequencing Data
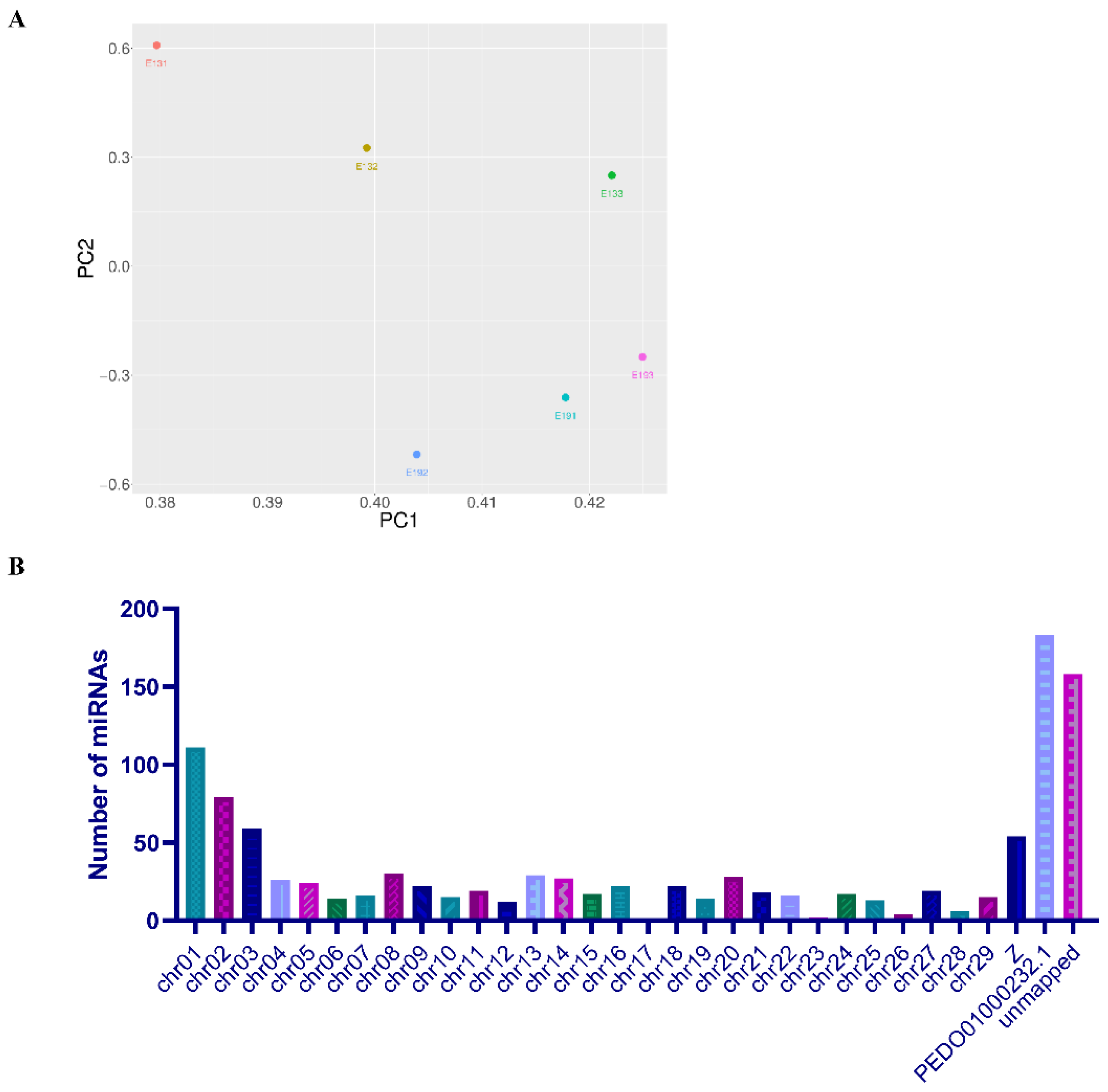
3.2. Characterization of Known miRNAs and Novel miRNAs
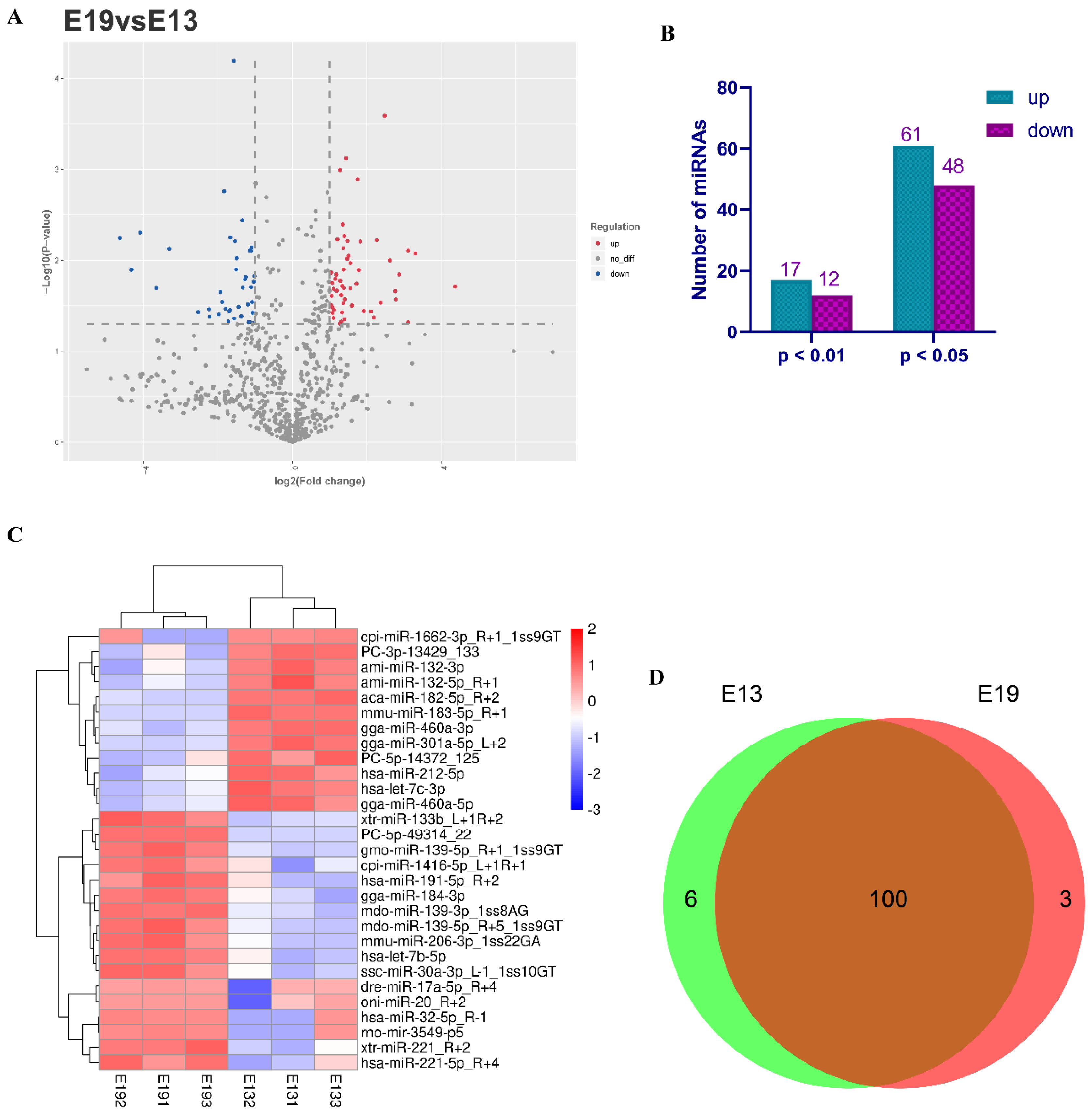
3.3. Identification of Differentially Expressed miRNAs
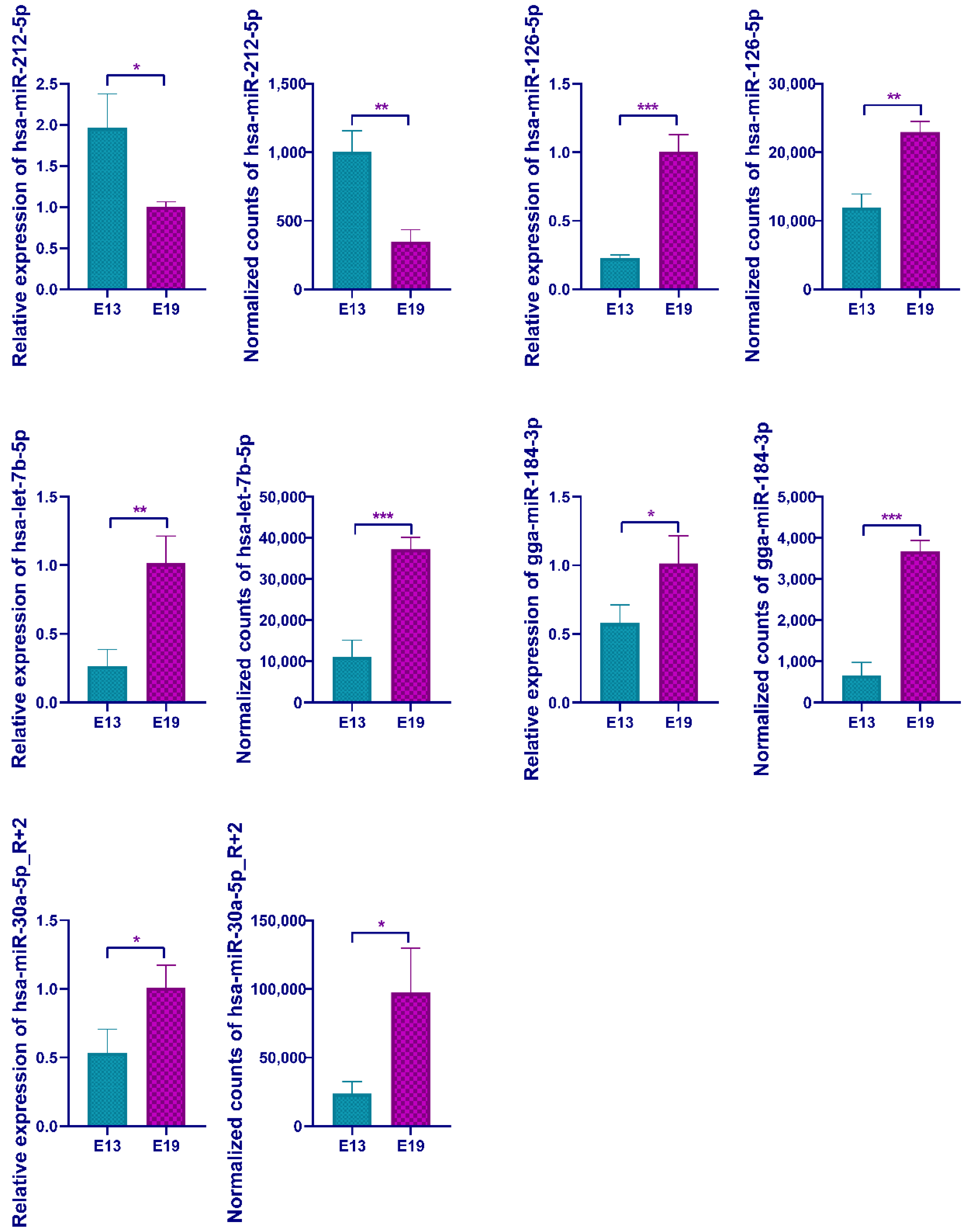
3.4. qPCR Validation of the Differentially Expressed miRNAs
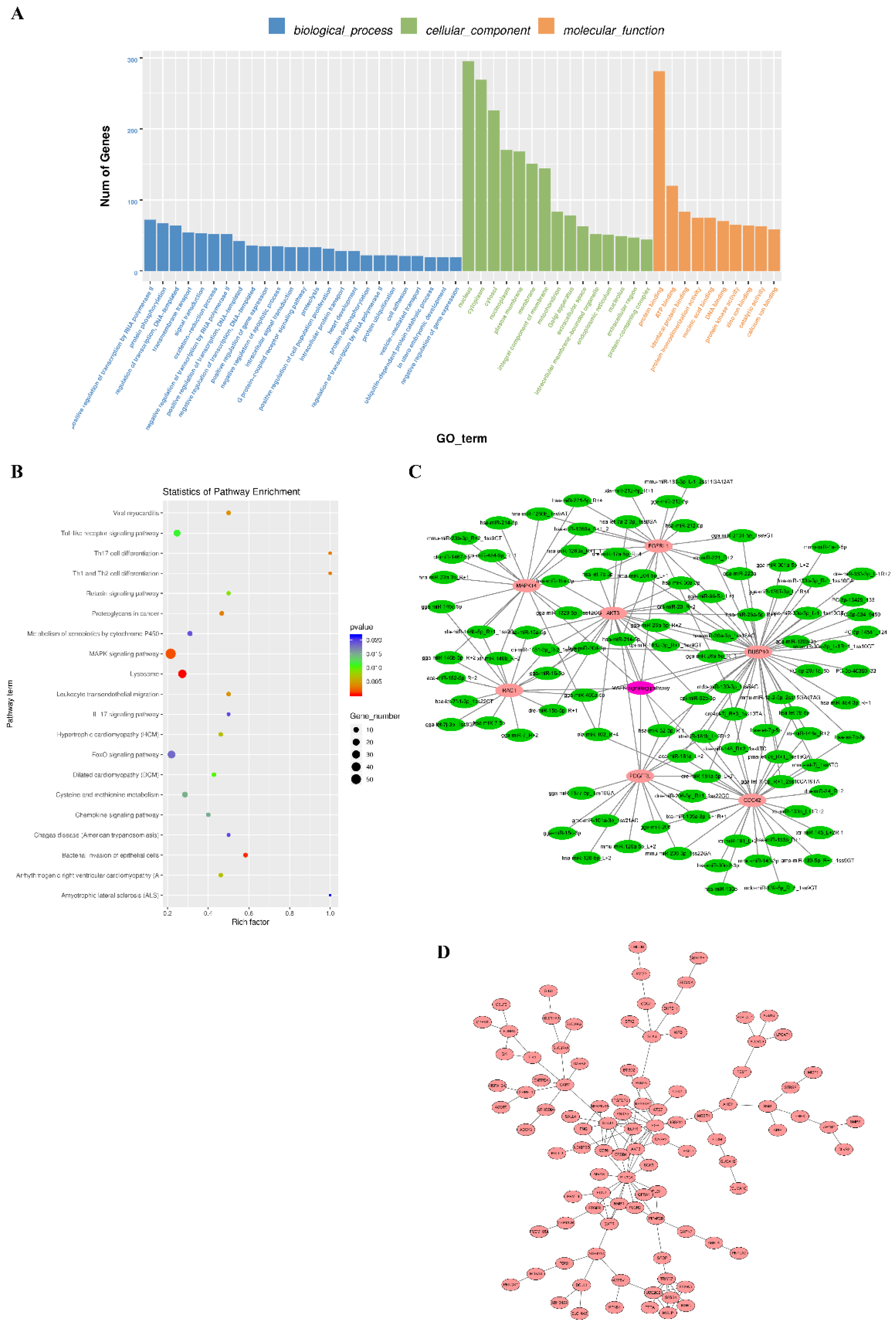
3.5. Target Gene Prediction and Enrichment Analysis
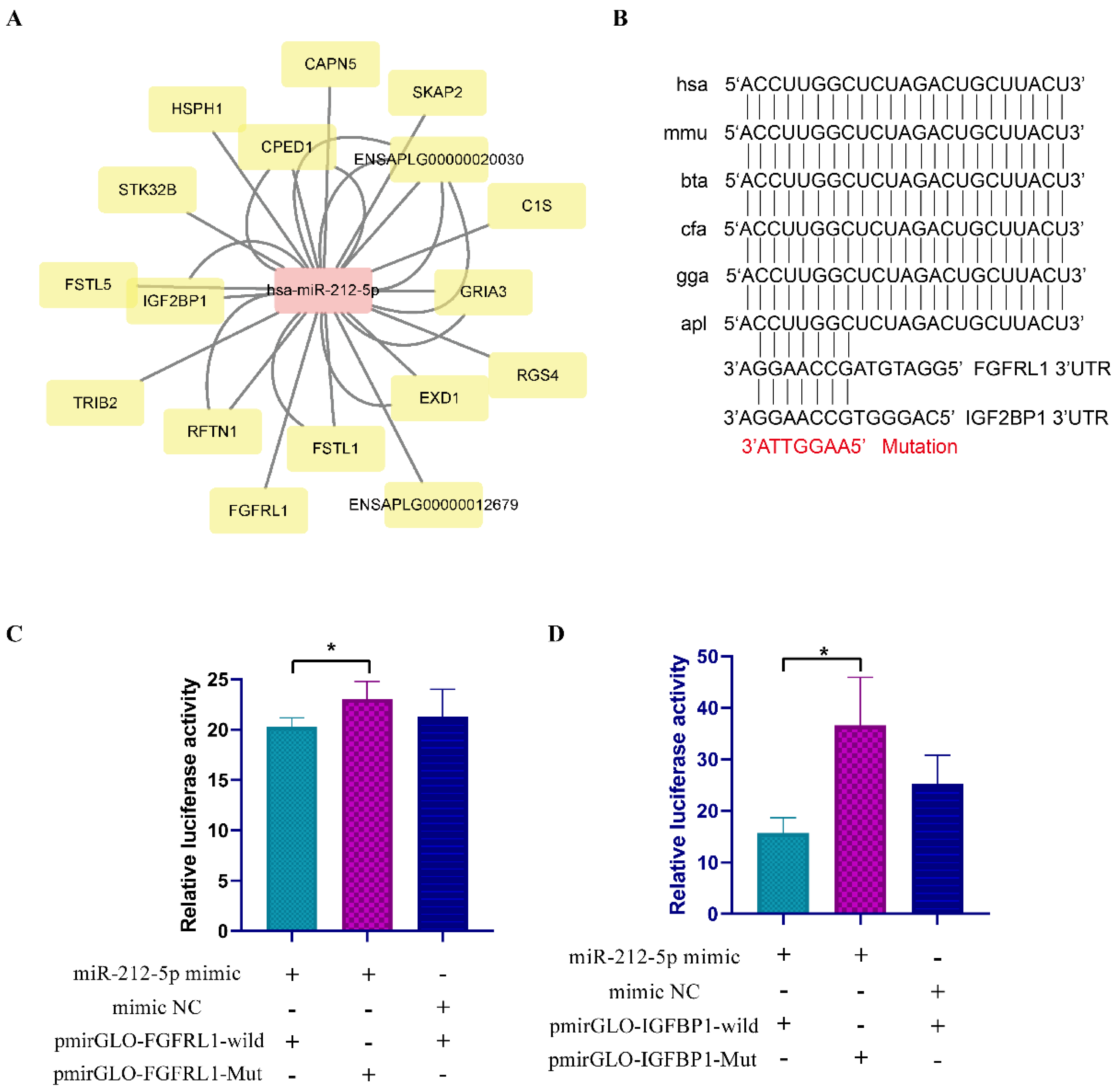
3.6. Verification of the Interaction between miRNA and Target Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.E.; Boxhorn, L.K. Regulation of skeletal muscle satellite cell proliferation and differentiation by transforming growth factor-beta, insulin-like growth factor I, and fibroblast growth factor. J. Cell. Physiol. 1989, 138, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Chen, X.; Wang, Z.; Yu, J.; Jia, X.; Li, Z.; Luo, W.; Abdalla, B.A.; Jebessa, E.; Nie, Q.; et al. Circular RNAs are abundant and dynamically expressed during embryonic muscle development in chickens. DNA Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cai, B.; Abdalla, B.A.; Zhu, X.; Zheng, M.; Han, P.; Nie, Q.; Zhang, X. LncIRS1 controls muscle atrophy via sponging miR-15 family to activate IGF1-PI3K/AKT pathway. J. Cachexia Sarcopenia Muscle 2019, 10, 391–410. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.-S.; Gu, L.-H.; Huang, W.; Xia, W.-L.; Zhang, Y.-S.; Zhang, Y.-G.; Rong, G.; Schachtschneider, K.; Hou, S.-S. Gene expression profiling in Pekin duck embryonic breast muscle. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.H.; Xu, T.S.; Huang, W.; Xie, M.; Shi, W.B.; Sun, S.D.; Hou, S.S. Developmental characteristics of pectoralis muscle in Pekin duck embryos. Genet. Mol. Res. 2013, 12, 6733–6742. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.-Y. Secreted microRNAs: A new form of intercellular communication. Trends Cell Biol. 2012, 22, 125–132. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Brisnt, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513. [Google Scholar] [CrossRef]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef]
- Luo, W.; Wu, H.; Ye, Y.; Li, Z.; Hao, S.; Kong, L.; Zheng, X.; Lin, S.; Nie, Q.; Zhang, X. The transient expression of miR-203 and its inhibiting effects on skeletal muscle cell proliferation and differentiation. Cell Death Dis. 2014, 5, e1347. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Ma, M.; Chen, B.; Li, Z.; Abdalla, B.A.; Nie, Q.; Zhang, X. MiR-16-5p targets SESN1 to regulate the p53 signaling pathway, affecting myoblast proliferation and apoptosis, and is involved in myoblast differentiation. Cell Death Dis. 2018, 9. [Google Scholar] [CrossRef]
- Chemello, F.; Grespi, F.; Zulian, A.; Cancellara, P.; Hebert-Chatelain, E.; Martini, P.; Bean, C.; Alessio, E.; Buson, L.; Bazzega, M.; et al. Transcriptomic Analysis of Single Isolated Myofibers Identifies miR-27a-3p and miR-142-3p as Regulators of Metabolism in Skeletal Muscle. Cell Rep. 2019, 26, 3784–3797. [Google Scholar] [CrossRef] [PubMed]
- Elsaeid Elnour, I.; Dong, D.; Wang, X.; Zhansaya, T.; Khan, R.; Jian, W.; Jie, C.; Chen, H. Bta-miR-885 promotes proliferation and inhibits differentiation of myoblasts by targeting MyoD1. J. Cell. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Huang, W.; Chen, B.; Jebessa, B.E.; Chen, X.; Cai, B.; Nie, Q. gga-mir-133a-3p Regulates Myoblasts Proliferation and Differentiation by Targeting PRRX1. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Lin, H.; Abdalla, B.; Nie, Q. Characterization of miR-206 Promoter and Its Association with Birthweight in Chicken. Int. J. Mol. Sci. 2016, 17, 559. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Gu, T.; Lu, L.; Cao, Z.; Song, Q.; Wang, Z.; Zhang, Y.; Chang, G.; Xu, Q.; Chen, G. Roles of miRNA-1 and miRNA-133 in the proliferation and differentiation of myoblasts in duck skeletal muscle. J. Cell. Physiol. 2019, 234, 3490–3499. [Google Scholar] [CrossRef]
- Gu, L.; Xu, T.; Huang, W.; Xie, M.; Sun, S.; Hou, S. Identification and Profiling of MicroRNAs in the Embryonic Breast Muscle of Pekin Duck. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Li, H.; Hu, Y.; Song, C.; Ji, G.; Liu, H.; Xu, W.; Ding, J. A New Primer for Sex Identification of Ducks and a Minimally Invasive Technique for Sampling of Allantoic Fluid to Detect Sex during Bird Embryo Development. Sex. Dev. 2015, 9, 173–181. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Li, X.; Shahid, M.; Wu, J.; Wang, L.; Liu, X.; Lu, Y. Comparative Small RNA Analysis of Pollen Development in Autotetraploid and Diploid Rice. Int. J. Mol. Sci. 2016, 17, 499. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Betel, D.; Koppal, A.; Agius, P.; Sander, C.; Leslie, C. Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol. 2010, 11, R90. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–Protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S. Rfam: An RNA family database. Nucleic Acids Res. 2003, 31, 439–441. [Google Scholar] [CrossRef]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2017, 46, D335–D342. [Google Scholar] [CrossRef]
- Jurka, J. Repbase Update: A database and an electronic journal of repetitive elements. Trends Genet. 2000, 16, 418–420. [Google Scholar] [CrossRef]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a database of repetitive elements in eukaryotic genomes. Mob. DNA 2015, 6. [Google Scholar] [CrossRef]
- Ouyang, H.; He, X.; Li, G.; Xu, H.; Jia, X.; Nie, Q.; Zhang, X. Deep Sequencing Analysis of miRNA Expression in Breast Muscle of Fast-Growing and Slow-Growing Broilers. Int. J. Mol. Sci. 2015, 16, 16242–16262. [Google Scholar] [CrossRef]
- Wang, Q.; Qi, R.; Wang, J.; Huang, W.; Wu, Y.; Huang, X.; Yang, F.; Huang, J. Differential expression profile of miRNAs in porcine muscle and adipose tissue during development. Gene 2017, 618, 49–56. [Google Scholar] [CrossRef]
- He, J.; Wang, W.; Lu, L.; Tian, Y.; Niu, D.; Ren, J.; Dong, L.; Sun, S.; Zhao, Y.; Chen, L.; et al. Analysis of miRNAs and their target genes associated with lipid metabolism in duck liver. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, S.; Wang, Y.; Deng, Y.; Yang, S.; Hu, J.; Li, L.; Wang, J. mRNA and miRNA Transcriptome Profiling of Granulosa and Theca Layers From Geese Ovarian Follicles Reveals the Crucial Pathways and Interaction Networks for Regulation of Follicle Selection. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhu, J.; Zhang, R.; Liang, W.; Ma, W.; Zhang, Q.; Huang, Z.; Ding, F.; Sun, H. miR-125b-5p targeting TRAF6 relieves skeletal muscle atrophy induced by fasting or denervation. Ann. Transl. Med. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; D’Souza, R.F.; Schierding, W.; Zeng, N.; Ramzan, F.; O’Sullivan, J.M.; Poppitt, S.D.; Cameron-Smith, D. Identification of human skeletal muscle miRNA related to strength by high-throughput sequencing. Physiol. Genom. 2018, 50, 416–424. [Google Scholar] [CrossRef]
- Zhou, G.; Li, C.; Feng, J.; Zhang, J.; Fang, Y. lncRNA UCA1 Is a Novel Regulator in Cardiomyocyte Hypertrophy through Targeting the miR-184/HOXA9 Axis. Cardiorenal. Med. 2018, 8, 130–139. [Google Scholar] [CrossRef]
- Liu, B.; Shi, Y.; He, H.; Cai, M.; Xiao, W.; Yang, X.; Chen, S.; Jia, X.; Wang, J.; Lai, S. miR-221 modulates skeletal muscle satellite cells proliferation and differentiation. Vitr. Cell. Dev. Biol. Anim. 2018, 54, 147–155. [Google Scholar] [CrossRef]
- Mollard, R.; Viville, S.; Ward, S.J.; Décimo, D.; Chambon, P.; Dollé, P. Tissue-Specific expression of retinoic acid receptor isoform transcripts in the mouse embryo. Mech. Dev. 2000, 94, 223–232. [Google Scholar] [CrossRef]
- Xie, S.-J.; Li, J.-H.; Chen, H.-F.; Tan, Y.-Y.; Liu, S.-R.; Zhang, Y.; Xu, H.; Yang, J.-H.; Liu, S.; Zheng, L.-L.; et al. Inhibition of the JNK/MAPK signaling pathway by myogenesis-associated miRNAs is required for skeletal muscle development. Cell Death Differ. 2018, 25, 1581. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, W.-Z.; Liu, T.; Feng, X.; Yang, N.; Zhou, H.-F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J. Recept. Signal Transduct. Res. 2015, 35, 600–604. [Google Scholar] [CrossRef]
- Shin, S.-Y.; Nguyen, L.K. Dissecting Cell-Fate Determination through Integrated Mathematical Modeling of the ERK/MAPK Signaling Pathway. Method. Mol. Biol. 2017, 1487, 409–432. [Google Scholar] [CrossRef]
- Stramucci, L.; Pranteda, A.; Bossi, G. Insights of Crosstalk between p53 Protein and the MKK3/MKK6/p38 MAPK Signaling Pathway in Cancer. Cancers 2018, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Matheny, R.W.; Geddis, A.V.; Abdalla, M.N.; Leandry, L.A.; Ford, M.; McClung, H.L.; Pasiakos, S.M. AKT2 is the predominant AKT isoform expressed in human skeletal muscle. Physiol. Rep. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; An, J.; Lian, L.; Qu, L.; Zheng, J.; Xu, G.; Yang, N. Polymorphisms in AKT3, FIGF, PRKAG3, and TGF-β genes are associated with myofiber characteristics in chickens. Poult. Sci. 2013, 92, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Vasyutina, E.; Martarelli, B.; Brakebusch, C.; Wende, H.; Birchmeier, C. The small G-proteins Rac1 and Cdc42 are essential for myoblast fusion in the mouse. Proc. Natl. Acad. Sci. USA 2009, 106, 8935. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-S.; Bae, G.-U.; Yi, M.-J.; Yang, Y.-J.; Oh, J.-E.; Takaesu, G.; Zhou, Y.T.; Low, B.C.; Krauss, R.S. A Cdo–Bnip-2–Cdc42 signaling pathway regulates p38α/β MAPK activity and myogenic differentiation. J. Cell Biol. 2008, 182, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Yamane, A.; Mayo, M.L.; Bringas, J.P.; Chen, L.; Huynh, M.; Thai, K.; Shum, L.; Slavkin, H.C.; Bringas, P. TGF-alpha, EGF, and their cognate EGF receptor are co-expressed with desmin during embryonic, fetal, and neonatal myogenesis in mouse tongue development. Dev. Dyn. 1997, 209, 353–366. [Google Scholar] [CrossRef]
- D’Andrea, P.; Sciancalepore, M.; Veltruska, K.; Lorenzon, P.; Bandiera, A. Epidermal Growth Factor—Based adhesion substrates elicit myoblast scattering, proliferation, differentiation and promote satellite cell myogenic activation. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 504–517. [Google Scholar] [CrossRef]
- Dobyns, W.B.; Mirzaa, G.M. Megalencephaly syndromes associated with mutations of core components of the PI3K-AKT–MTOR pathway: PIK3CA, PIK3R2, AKT3, and MTOR. Am. J. Med. Genet. Part C Semin. Med. Genet. 2019, 181, 582–590. [Google Scholar] [CrossRef]
- Timmer, L.T.; Hoogaars, W.M.H.; Jaspers, R.T. The Role of IGF-1 Signaling in Skeletal Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 109–137. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Mapped Reads | Mapping Percentage | Validated Reads | Unique Reads |
|---|---|---|---|---|---|
| E131 | 14,875,288 | 13,717,484 | 92.22% | 13,445,305 | 295,703 |
| E132 | 13,189,920 | 11,378,369 | 86.27% | 10,042,092 | 355,298 |
| E133 | 18,065,989 | 16,529,699 | 91.50% | 16,320,782 | 376,515 |
| E191 | 11,626,527 | 9,711,731 | 83.53% | 8,707,352 | 267,865 |
| E192 | 13,795,701 | 12,385,025 | 89.77% | 12,472,937 | 236,010 |
| E193 | 16,353,157 | 14,598,732 | 89.27% | 13,723,856 | 329,106 |
| miRNAs | Normalized Counts | |||||
|---|---|---|---|---|---|---|
| E131 | E132 | E133 | E191 | E192 | E193 | |
| hsa-miR-125b-5p | 891,404 | 859,062 | 569,685 | 381,424 | 317,414 | 408,938 |
| hsa-miR-100-5p_1ss9GT | 631,277 | 624,407 | 471,452 | 345,984 | 356,079 | 419,505 |
| hsa-miR-99a-5p_R-1_1ss9GT | 542,627 | 567,141 | 448,064 | 345,143 | 336,464 | 387,616 |
| hsa-miR-92a-3p | 424,997 | 675,529 | 338,265 | 249,506 | 212,056 | 265,128 |
| hsa-miR-140-3p_L-1R + 2_1ss10GT | 1,261,316 | 57,304 | 318,557 | 129,118 | 61,786 | 109,581 |
| hsa-miR-30c-5p_R + 1 | 277,921 | 264,896 | 237,026 | 395,513 | 351,692 | 357,762 |
| hsa-miR-199a-5p | 392,226 | 260,782 | 335,561 | 220,482 | 246,505 | 242,580 |
| gga-miR-26a-5p_R + 1 | 241,562 | 144,476 | 224,490 | 320,746 | 422,701 | 315,698 |
| hsa-miR-199a-3p_R-1 | 284,056 | 212,832 | 308,059 | 212,511 | 245,447 | 192,145 |
| hsa-miR-30d-5p_R + 2 | 181,276 | 149,487 | 160,244 | 232,991 | 270,104 | 255,296 |
| hsa-miR-214-3p | 366,065 | 343,733 | 222,584 | 108,833 | 79,526 | 101,199 |
| gga-miR-206 | 123,149 | 175,227 | 144,083 | 289,543 | 273,010 | 202,096 |
| gga-miR-130b-3p | 221,332 | 229,131 | 209,359 | 181,602 | 176,295 | 158,693 |
| hsa-miR-363-3p_1ss9GT | 147,958 | 188,728 | 183,421 | 179,713 | 215,735 | 184,929 |
| hsa-miR-133a-3p_L-1R + 1 | 72,388 | 76,236 | 72,565 | 257,146 | 273,573 | 197,363 |
| mmu-miR-143-3p | 99,894 | 93,780 | 128,401 | 178,804 | 238,750 | 170,472 |
| hsa-miR-10a-5p_R-1 | 160,898 | 141,885 | 210,035 | 123,003 | 92,059 | 118,950 |
| hsa-let-7g-5p | 76,031 | 94,215 | 88,589 | 131,530 | 165,784 | 122,125 |
| hsa-miR-148a-3p | 96,660 | 74,660 | 104,199 | 101,884 | 150,264 | 132,439 |
| hsa-miR-221-3p | 66,915 | 82,560 | 87,721 | 79,087 | 87,797 | 89,385 |
| mmu-miR-27b-3p_1ss9GT | 60,768 | 67,128 | 67,252 | 80,895 | 120,432 | 90,534 |
| mmu-miR-181a-5p | 89,950 | 65,948 | 71,395 | 82,974 | 90,517 | 80,138 |
| gga-miR-191-5p_R-1 | 61,450 | 67,512 | 45,486 | 105,610 | 88,121 | 100,506 |
| gga-miR-2954_R + 1 | 85,299 | 123,188 | 72,930 | 53,778 | 38,901 | 57,196 |
| gga-miR-222a | 68,825 | 81,020 | 77,822 | 52,757 | 50,097 | 64,669 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Xiong, T.; Zhou, M.; Wan, L.; Xi, S.; Liu, Q.; Chen, Y.; Mao, H.; Liu, S.; Chen, B. Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck. Animals 2020, 10, 1417. https://doi.org/10.3390/ani10081417
Li C, Xiong T, Zhou M, Wan L, Xi S, Liu Q, Chen Y, Mao H, Liu S, Chen B. Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck. Animals. 2020; 10(8):1417. https://doi.org/10.3390/ani10081417
Chicago/Turabian StyleLi, Chuan, Ting Xiong, Mingfang Zhou, Lei Wan, Suwang Xi, Qiuhong Liu, Yi Chen, Huirong Mao, Sanfeng Liu, and Biao Chen. 2020. "Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck" Animals 10, no. 8: 1417. https://doi.org/10.3390/ani10081417
APA StyleLi, C., Xiong, T., Zhou, M., Wan, L., Xi, S., Liu, Q., Chen, Y., Mao, H., Liu, S., & Chen, B. (2020). Characterization of microRNAs during Embryonic Skeletal Muscle Development in the Shan Ma Duck. Animals, 10(8), 1417. https://doi.org/10.3390/ani10081417