Genetics of Lifetime Reproductive Performance in Italian Heavy Draught Horse Mares



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subject and Organization of the Study
2.2. Training Dataset and Analysis
- the percentage squared bias (PSB; [26]), obtained from the formula:where y is a vector of actual and ŷ is a vector of predicted values;PSB = 100 (y − ŷ)’ (y − ŷ)/(y’ y)
- the mean absolute deviation of residuals (MAD; [27]) calculated from the formula:where |y − ŷ| are absolute differences between actual (y) and predicted values (ŷ), respectively, and n is the number of observations;MAD = Σ |(y − ŷ)|/n
- the standard deviation of residuals obtained as y − ŷ, where y and ŷ are vectors of actual and predicted values, respectively.
2.3. Full Dataset and Analysis
3. Results
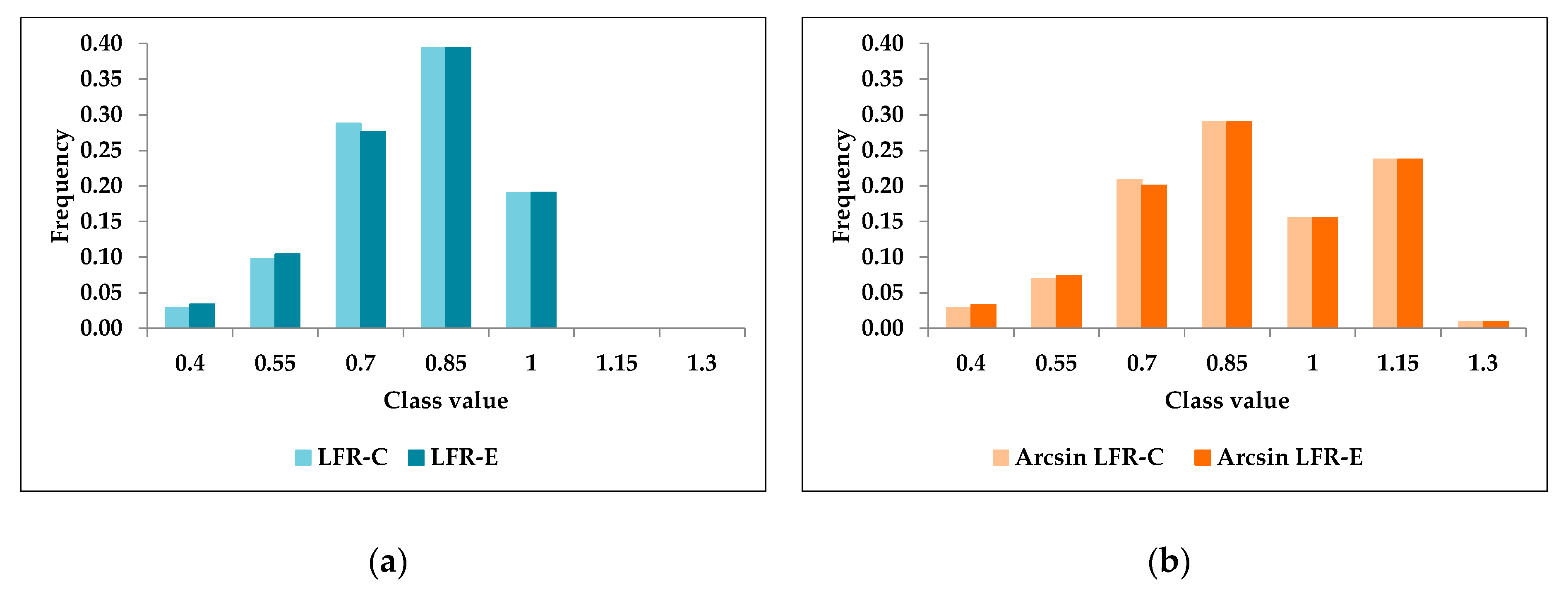
3.1. Validation of the Phenotypic Variable to Measure Lifetime Fertility
3.2. Genetic Analysis of Lifetime Fertility
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stott, A.W.; Veerkamp, R.F.; Wassel, T.R. The economics of fertility in the dairy herd. Anim. Sci. 1999, 68, 49–57. [Google Scholar] [CrossRef]
- Langlois, B.; Blouin, C. Statistical analysis of some factors affecting the number of horse births in France. Reprod. Nutr. Dev. 2004, 44, 583–595. [Google Scholar] [CrossRef]
- Bormann, J.M.; Wilson, D.E. Calving day and age at first calving in Angus heifers. J. Anim. Sci. 2010, 88, 1947–1956. [Google Scholar] [CrossRef]
- Meyer, K.; Hammond, K.; Parnell, P.F.; MacKinnon, M.J.; Sivarajasingam, S. Estimates of heritability and repeatability for reproductive traits in Australian beef cattle. Livest. Prod. Sci. 1990, 25, 15–30. [Google Scholar] [CrossRef]
- Phocas, F.; Bloch, C.; Chapelle, P.; Becherel, F.; Renand, G.; Menissier, F. Developing a breeding objective for a French purebred beef cattle selection programme. Livest. Prod. Sci. 1998, 57, 49–65. [Google Scholar] [CrossRef]
- Robinson, D.L. Days to calving in artificially inseminated cattle: Alternative models and analyses. Livest. Sci. 2008, 117, 34–42. [Google Scholar] [CrossRef]
- Liu, Z.; Jaitner, J.; Reinhardt, F.; Pasman, E.; Rensing, S.; Reents, R. Genetic evaluation of fertility traits of dairy cattle using a multiple-trait animal model. J. Dairy Sci. 2008, 91, 4333–4343. [Google Scholar] [CrossRef] [PubMed]
- VanRaden, P.M.; Tooker, M.E.; Wright, J.R.; Sun, C.; Hutchison, J.L. Comparison of single-trait to multi-trait national evaluations for yield, health, and fertility. J. Dairy Sci. 2014, 97, 7952–7962. [Google Scholar] [CrossRef] [PubMed]
- Dezetter, C.; Leclerc, H.; Mattalia, S.; Barbat, A.; Boichard, D.; Ducrocq, V. Inbreeding and crossbreeding parameters for production and fertility traits in Holstein, Montbéliarde, and Normande cows. J. Dairy Sci. 2015, 98, 4904–4913. [Google Scholar] [CrossRef]
- Vidament, M. French field results (1985–2005) on factors affecting fertility of frozen stallion semen. Anim. Reprod. Sci. 2005, 89, 115–136. [Google Scholar] [CrossRef]
- Guillaume, D.; Salazar-Ortiz, J.; Martin-Rosset, W. Effects of nutrition level in mares’ ovarian activity and in equines’ puberty. In Nutrition and Feeding of the Broodmare; Miraglia, N., Martin-Rosset, W., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 315–339. [Google Scholar]
- Davies Morel, M.C.G. Equine Reproductive Physiology Breeding and Stud Management, 3rd ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Ellis, A.D.; Boekhoff, M.; Bailoni, L.; Mantovani, R. Nutrition and equine fertility. In Nutrition and Feeding of the Broodmare; Miraglia, N., Martin-Rosset, W., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 341–366. [Google Scholar]
- Giesecke, K.; Sieme, H.; Distl, O. Infertility and candidate gene markers for fertility in stallions: A review. Vet. J. 2010, 185, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Hemberg, E.; Lundeheim, N.; Einarsson, S. Reproductive Performance of Thoroughbred Mares in Sweden. Reprod. Domest. Anim. 2004, 39, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, R.; Sartori, C.; Pigozzi, G. Retrospective and statistical analysis of breeding management on the Italian Heavy Draught Horse breed. Animal 2013, 7, 1053–1059. [Google Scholar] [CrossRef]
- Wolc, A.; Torzynski, G.; Szwaczkowski, T. Genetic effects on reproductive traits in Warmblood horses. Can. J. Anim. Sci. 2009, 89, 215–218. [Google Scholar] [CrossRef]
- Sairanen, J.; Nivola, K.; Katila, T.; Virtala, A.M.; Ojala, M. Effects of inbreeding and other genetic components on equine fertility. Animal 2009, 3, 1662–1672. [Google Scholar] [CrossRef] [PubMed]
- Mucha, S.; Wolc, A.; Szwaczkowski, T. Bayesian and REML analysis of twinning and fertility in Thoroughbred horses. Livest. Sci. 2012, 144, 82–88. [Google Scholar] [CrossRef]
- Sabeva, I.; Apostolov, A. Longevity and life reproductive efficiency in Arabian broodmares. Arch. Zootech. 2011, 14, 66–74. [Google Scholar]
- Cammack, K.M.; Thomas, M.G.; Enns, R.M. Review: Reproductive traits and their heritabilities in beef cattle. Prof. Anim. Sci. 2009, 25, 517–528. [Google Scholar] [CrossRef]
- Ponzoni, R.W. Which trait for genetic improvement of beef cattle reproduction-calving rate or calving day? J. Anim. Breed. Genet. 1992, 109, 119–128. [Google Scholar] [CrossRef]
- Santschi, E.M.; Leibsle, S.R.; Morehead, J.P.; Prichard, M.A.; Clayton, M.K.; Keuler, N.S. Carpal and fetlock conformation of the juvenile Thoroughbred from birth to yearling auction age. Equine Vet. J. 2006, 38, 604–609. [Google Scholar] [CrossRef]
- Folla, F.; Sartori, C.; Guzzo, N.; Pigozzi, G.; Mantovani, R. Genetics of linear type traits scored on young foals belonging to the Italian Heavy Draught Horse breed. Livest. Sci. 2019, 219, 91–96. [Google Scholar] [CrossRef]
- Mantovani, R.; Pigozzi, G.; Bittante, G. The Italian Heavy Draught Horse breed: Origin, breeding program, efficiency of the selection scheme and inbreeding. Publ. Assoc. Anim. Prod. 2005, 116, 155. [Google Scholar]
- Ali, T.E.; Schaeffer, L.R. Accounting for covariances among test day milk yields in dairy cows. Can. J. Anim. Sci. 1987, 67, 637–644. [Google Scholar] [CrossRef]
- Vargas, B.; Koops, W.J.; Herrero, M.; Van Arendonk, J.A.M. Modeling extended lactations of dairy cows. J. Dairy Sci. 2000, 83, 1371–1380. [Google Scholar] [CrossRef]
- Fernandez, G.C.J. Residual analysis and data transformations: Important tools in statistical analysis. HortScience 1992, 27, 297–300. [Google Scholar] [CrossRef]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Automat. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Aguilar, I.; Misztal, I. Technical note: Recursive algorithm for inbreeding coefficients assuming nonzero inbreeding of unknown parents. J. Dairy Sci. 2008, 91, 1669–1672. [Google Scholar] [CrossRef]
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Aguilar, I.; Legarra, A.; Vitezica, Z. Manual for BLUPF90 Family of Programs; University of Georgia: Athens, GA, USA, 2014. [Google Scholar]
- Falconer, D.S. Quantitative Genetics, 3rd ed.; Longman Scientific and Technical: Harlow, UK, 1989; pp. 182–183. [Google Scholar]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Mrode, R.A. Linear Models for the Prediction of aNimal Breeding Values, 3rd ed.; CABI Publishing Company: Cambridge, UK, 2014. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Gottschalk, M.; Sieme, H.; Martinsson, G.; Distl, O. Heritability of semen traits in German Warmblood stallions. Anim. Reprod. Sci. 2016, 170, 10–14. [Google Scholar] [CrossRef]
- Albertsdóttir, E.; Arnason, T.; Eriksson, S.; Sigurdsson, A.; Fikse, W.F. Effects of integrated genetic evaluations for Icelandic horses on predictive ability, accuracy and selection bias. J. Anim. Breed. Genet. 2012, 129, 41–49. [Google Scholar] [CrossRef]
- Reverter, A.; Golden, B.L.; Bourdon, R.M.; Brinks, J.S. Technical note: Detection of bias in genetic predictions. J. Anim. Sci. 1994, 72, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Olsen, H.F.; Klemetsdal, G.; Ødegård, J.; Árnason, T. Validation of alternative models in genetic evaluation of racing performance in North Swedish and Norwegian cold-blooded trotters. J. Anim. Breed. Genet. 2012, 129, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Guerrero, M.J.; Cervantes, I.; Valera, M.; Gutiérrez, J.P. Modelling genetic evaluation for dressage in Pura Raza Español horses with focus on the rider effect. J. Anim. Breed. Genet. 2014, 131, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.J.; Bunter, K.L. Days to calving in Angus cattle: Genetic and environmental effects, and covariances with other traits. Livest. Prod. Sci. 1996, 45, 13–22. [Google Scholar] [CrossRef]
- Gutiérrez, J.P.; Alvarez, I.; Fernández, I.; Royo, L.J.; Díez, J.; Goyache, F. Genetic relationships between calving date, calving interval, age at first calving and type traits in beef cattle. Livest. Prod. Sci. 2002, 78, 215–222. [Google Scholar] [CrossRef]
- Evans, J.L.; Golden, B.L.; Bourdon, R.M.; Long, K.L. Additive genetic relationships between heifer pregnancy and scrotal circumference in Hereford cattle. J. Anim. Sci. 1999, 77, 2621–2628. [Google Scholar] [CrossRef]
- Doyle, S.P.; Golden, B.L.; Green, R.D.; Brinks, J.S. Additive genetic parameter estimates for heifer pregnancy and subsequent reproduction in Angus females. J. Anim. Sci. 2000, 78, 2091–2098. [Google Scholar] [CrossRef]
- Thallman, R.M.; Cundiff, L.V.; Gregory, K.E.; Koch, R.M. Germplasm evaluation in beef cattle-Cycle IV: Postweaning growth and puberty of heifers. J. Anim. Sci. 1999, 77, 2651–2659. [Google Scholar] [CrossRef]
- Morris, C.A.; Wilson, J.A.; Bennett, G.L.; Cullen, N.G.; Hickey, S.M.; Hunter, J.C. Genetic parameters for growth, puberty, and beef cow reproductive traits in a puberty selection experiment. N. Z. J. Agric. Res. 2000, 43, 83–91. [Google Scholar] [CrossRef]
- Clément, V.; Bibé, B.; Verrier, É.; Elsen, J.M.; Manfredi, E.; Bouix, J.; Hanocq, É. Simulation analysis to test the influence of model adequacy and data structure on the estimation of genetic parameters for traits with direct and maternal effects. Genet. Sel. Evol. 2001, 33, 369–396. [Google Scholar] [CrossRef]
- Seierø, T.; Mark, T.; Jönsson, L. Genetic parameters for longevity and informative value of early indicator traits in Danish show jumping horses. Livest. Sci. 2016, 184, 126–133. [Google Scholar] [CrossRef]
- Solé, M.; Sánchez, M.J.; Valera, M.; Molina, A.; Azor, P.J.; Sölkner, J.; Mészáros, G. Assessment of sportive longevity in Pura Raza Español dressage horses. Livest. Sci. 2017, 203, 69–75. [Google Scholar] [CrossRef]
- Folla, F. Genetic of Linear Type Traits and Lifetime Fertility Rate in Italian Heavy Draught Horse. Ph.D. Thesis, University of Padova, Legnaro, Padova, Italy, 30 July 2015. Available online: http://paduaresearch.cab.unipd.it/8915/1/Folla_Fabio_Tesi.pdf (accessed on 22 June 2020).


{kind=link}
| Item | Training Dataset | Full Dataset |
|---|---|---|
| Mares with actual records, number (no.) | 1487 | 3033 |
| Mares with projected records, no. | - | 1443 |
| Environmental units 1 by birth year (EU-BY), no. | 97 | 125 |
| Mean records in EU-BY, no. | 15.3 | 24.2 |
| Age at first known breeding season, months (mo.) | 43.7 ± 6.6 | 43.9 ± 6.5 |
| 3 years first foaling mares, mo. | 36.6 ± 1.8 | 36.6 ± 1.8 |
| 4 years first foaling mares, mo. | 49.1 ± 2.5 | 49.0 ± 2.4 |
| Animals in the pedigree file, no. | - | 6803 |
| Sires of mares with record, no. | 400 | 602 |
| Dams of mares with record, no. | 1011 | 1848 |
| Daughters/sire | 3.5 | 4.8 |
| Daughters/dam | 1.3 | 1.5 |
| Item | Projection Method | |
|---|---|---|
| Coefficients (LFR-C) | Equations (LFR-E) | |
| Projection from 3 known breeding seasons | ||
| - PSB 1 | 0.0604 | 0.0148 |
| - MAD 2 | 0.1813 | 0.0793 |
| - SDR 3 | 0.1028 | 0.0986 |
| Projection from 4 known breeding seasons | ||
| - PSB | 0.0437 | 0.0092 |
| - MAD | 0.1561 | 0.0606 |
| - SDR | 0.0832 | 0.0784 |
| Projection from 5 known breeding seasons | ||
| - PSB | 0.0319 | 0.0041 |
| - MAD | 0.1377 | 0.0350 |
| - SDR | 0.0622 | 0.0537 |
| Average | ||
| - PSB | 0.0453 | 0.0094 |
| - MAD | 0.1584 | 0.0583 |
| - SDR | 0.0827 | 0.0769 |
| Statistic | LFR-C 1 | LRF-E 2 | Arcsine LFR-C 3 | Arcsine LFR-E 4 |
|---|---|---|---|---|
| Mean ± standard deviation | 0.700 ± 0.142 | 0.699 ± 0.144 | 0.794 ± 0.195 | 0.793 ± 0.197 |
| Kolmogorov-Smirnov D | 0.16 (p < 0.01) | 0.14 (p < 0.01) | 0.15 (p < 0.01) | 0.11 (p < 0.01) |
| Anderson-Darling A-Sq | 82.9 (p < 0.01) | 78.7 (p < 0.01) | 67.2 (p < 0.01) | 60.1 (p < 0.01) |
| Skewness | −0.881 | 0.144 | −0.506 | 0.197 |
| Kurtosis | 0.485 | 0.986 | −0.300 | −0.004 |
| Item | LFR-C | LRF-E | Arcsine LFR-C | Arcsine LFR-E |
|---|---|---|---|---|
| Herd Variance 1 | 0.141 | 0.160 | 0.259 | 0.276 |
| Genetic Variance 1 | 4.848 | 4.693 | 8.975 | 8.829 |
| Dominance Variance 1 | 1.628 | 1.358 | 2.896 | 2.573 |
| Residual Variance 1 | 13.426 | 14.259 | 25.660 | 25.463 |
| Phenotypic Variance 1 | 20.043 | 19.469 | 37.790 | 37.141 |
| Heritability | 0.242 | 0.241 | 0.238 | 0.237 |
| SE Heritability | 0.043 | 0.042 | 0.042 | 0.042 |
| AIC | −3290 | −3380 | −1370 | −1424 |
| Comparison | Mares with Actual or Predicted LFR | Stallions with ≥9 Daughters with Actual or Predicted LFR |
|---|---|---|
| LFR-C 2 vs. LFR-E 3 | 0.997 | 0.993 |
| LFR-C vs. Arcsine LFR-C 4 | 0.996 | 0.996 |
| LFR-E vs. Arcsine LFR-E 5 | 0.996 | 0.995 |
| Arcsine LFR-C vs. Arcsine LFR-E | 0.997 | 0.993 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantovani, R.; Folla, F.; Pigozzi, G.; Tsuruta, S.; Sartori, C. Genetics of Lifetime Reproductive Performance in Italian Heavy Draught Horse Mares. Animals 2020, 10, 1085. https://doi.org/10.3390/ani10061085
Mantovani R, Folla F, Pigozzi G, Tsuruta S, Sartori C. Genetics of Lifetime Reproductive Performance in Italian Heavy Draught Horse Mares. Animals. 2020; 10(6):1085. https://doi.org/10.3390/ani10061085
Chicago/Turabian StyleMantovani, Roberto, Fabio Folla, Giuseppe Pigozzi, Shogo Tsuruta, and Cristina Sartori. 2020. "Genetics of Lifetime Reproductive Performance in Italian Heavy Draught Horse Mares" Animals 10, no. 6: 1085. https://doi.org/10.3390/ani10061085
APA StyleMantovani, R., Folla, F., Pigozzi, G., Tsuruta, S., & Sartori, C. (2020). Genetics of Lifetime Reproductive Performance in Italian Heavy Draught Horse Mares. Animals, 10(6), 1085. https://doi.org/10.3390/ani10061085