Is Dietary 2-Oxoglutaric Acid Effective in Accelerating Bone Growth and Development in Experimentally-Induced Intrauterine Growth Retarded Gilts?

 ,
,  , , and
, , and
Simple Summary
Abstract
1. Introduction
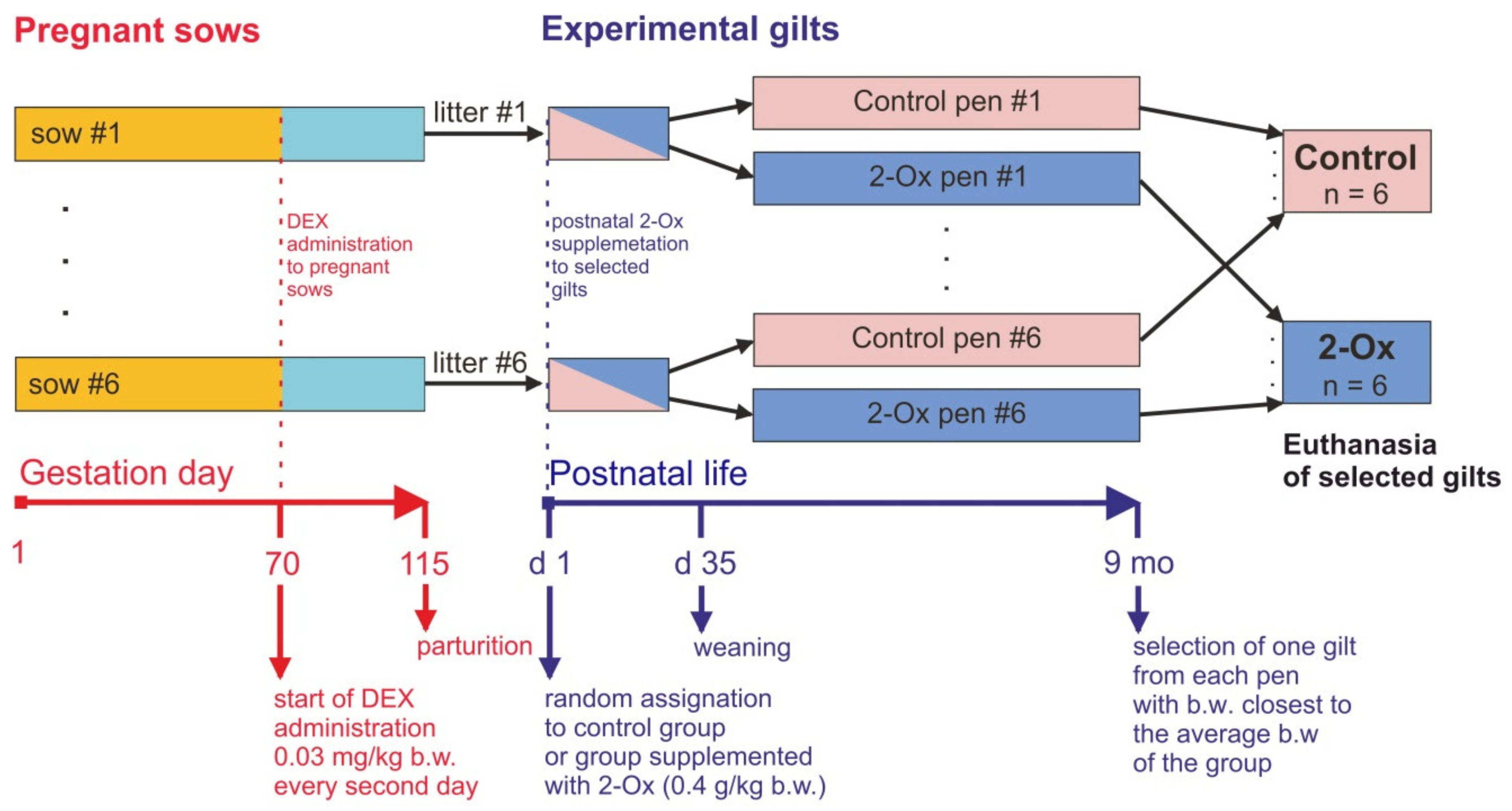
2. Materials and Methods
2.1. Ethical Approval
2.2. Pregnant and Lactating Sows
2.3. Offspring
2.4. Serum Biochemical and Hormonal Analysis
2.5. Bone Analysis
2.6. Histomorphometry
2.7. Immunohistochemistry
2.8. Statistical Analysis
3. Results
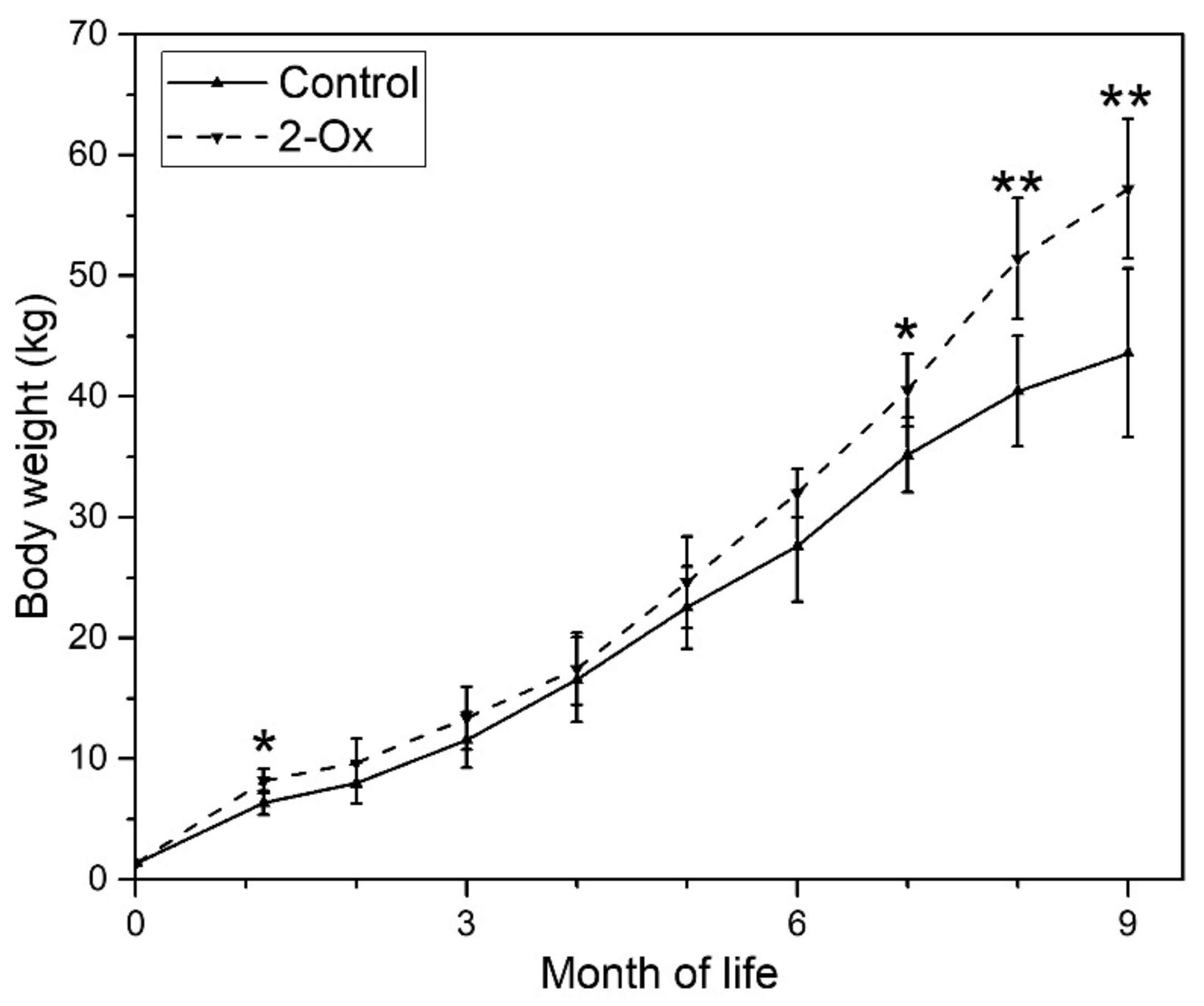
3.1. Body Weight
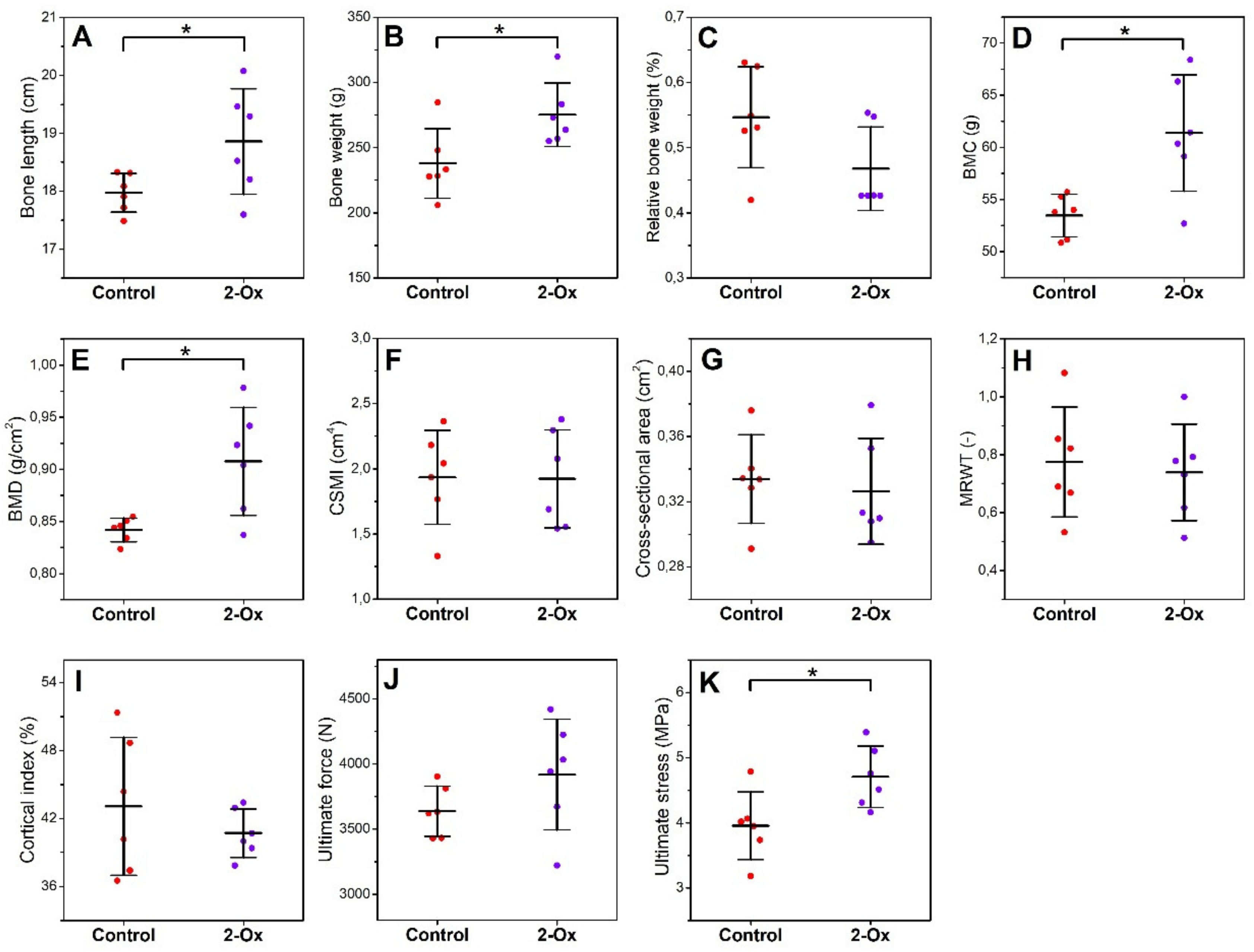
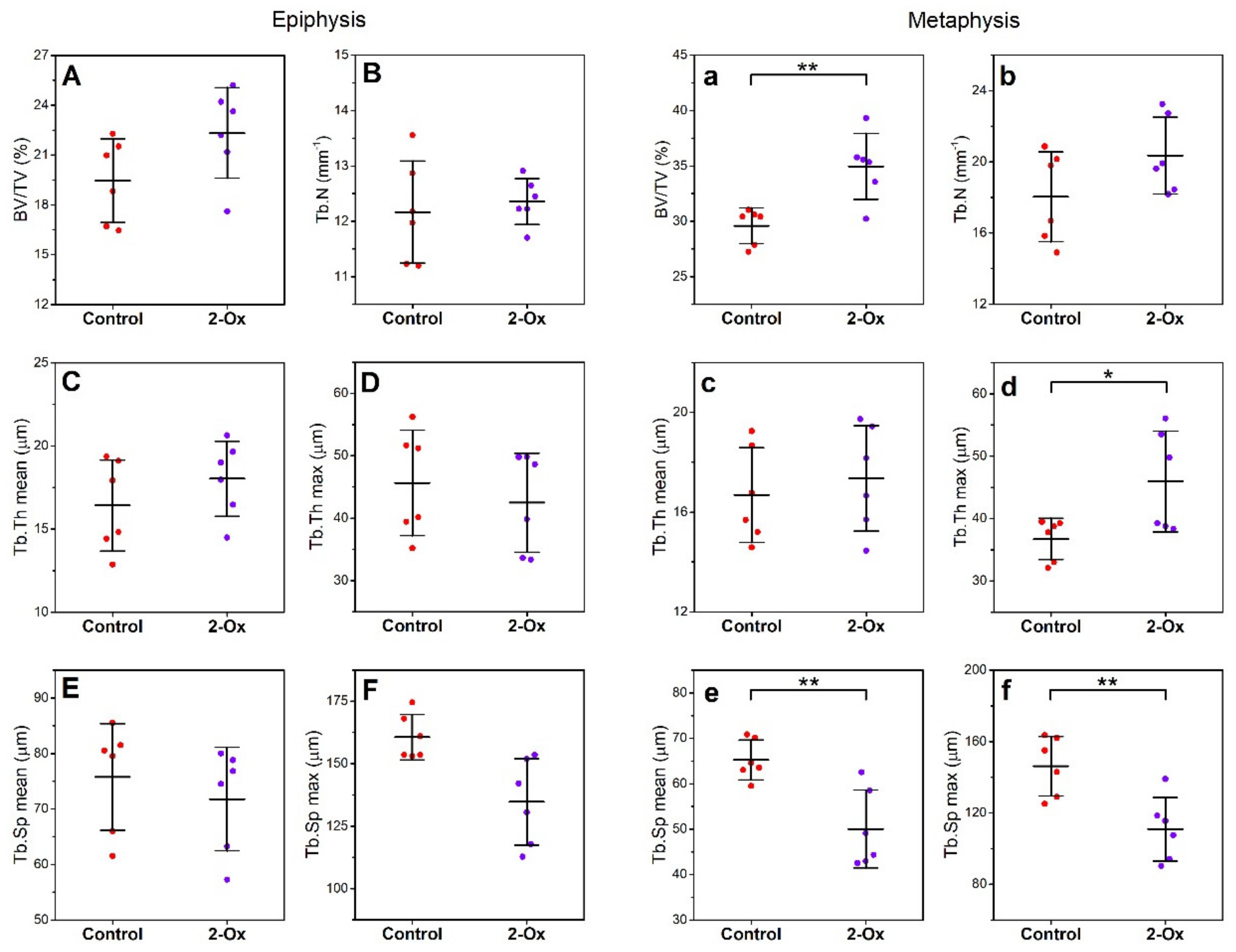
3.2. Bone Morphology, Geometry, Density, and Mechanical Properties
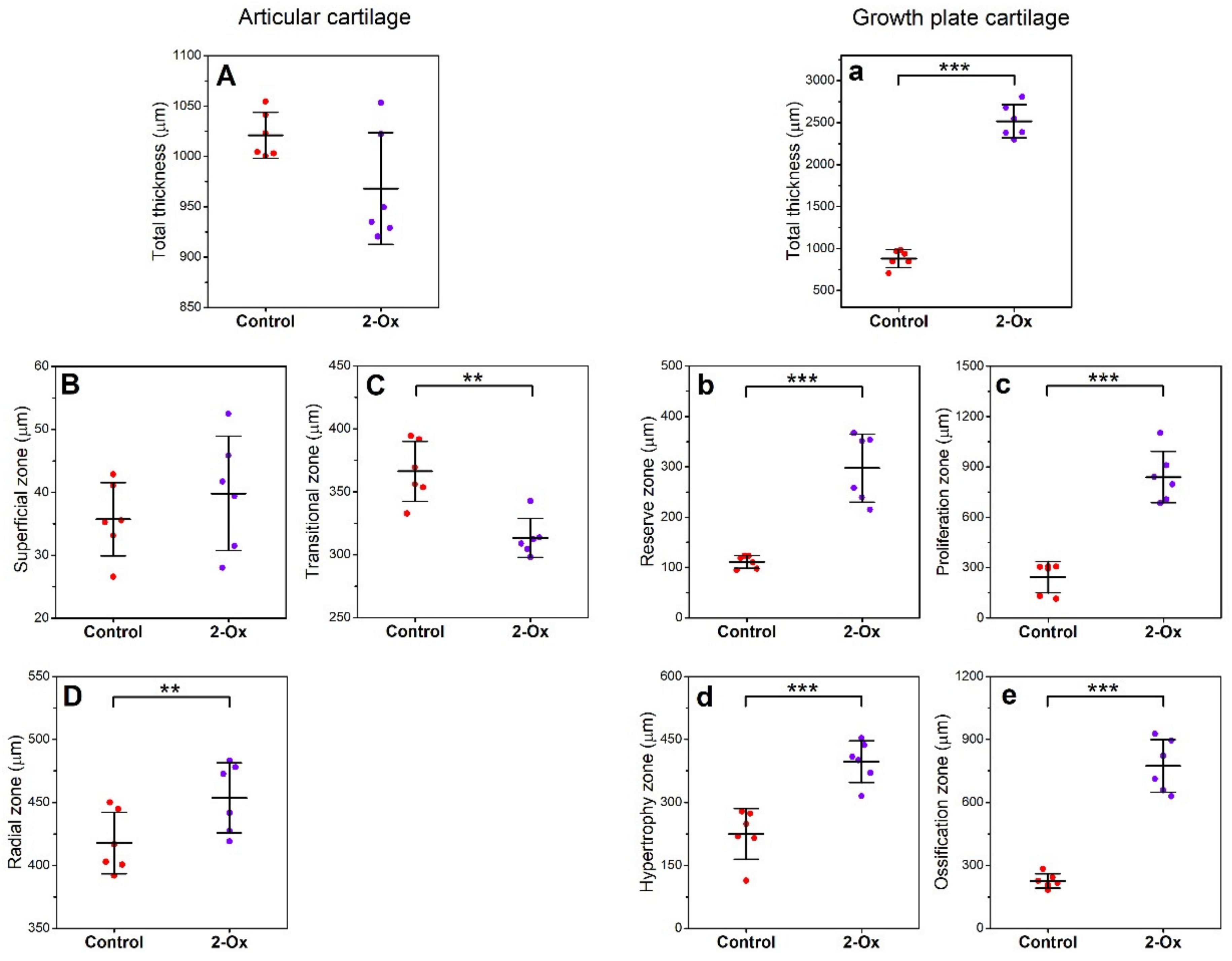
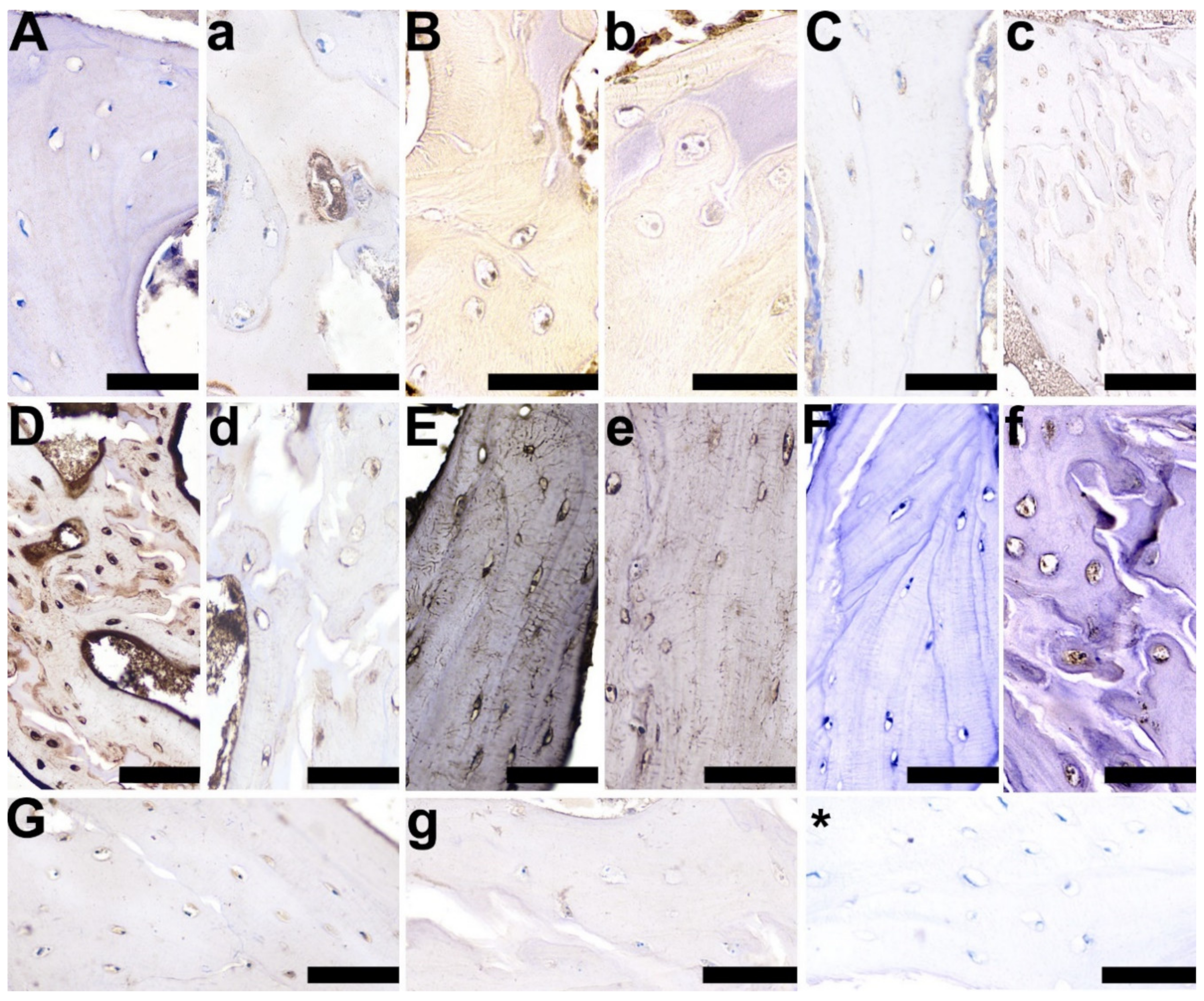
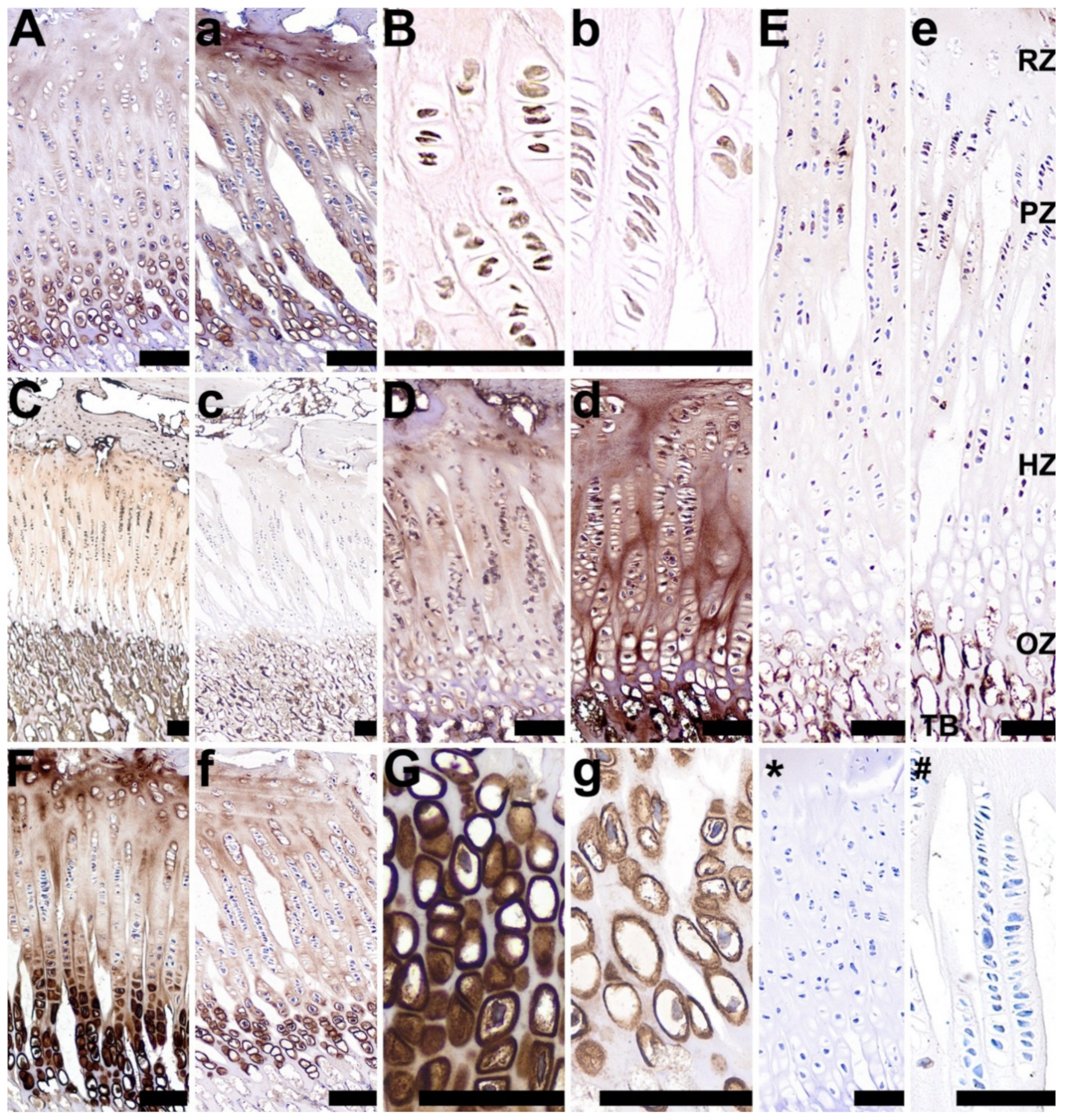
3.3. Articular Cartilage, Growth Plate Cartilage, and Trabecular Bone Morphology
3.4. Serum Biochemical and Hormonal Analysis
3.5. OC, OPG, RANKL, TIMP-2, VEGF, BMP-2, and GR Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holemans, K.; Aertis, L.; Van Assche, A. Fetal growth and long-term consequences in animal models of growth retardation. Eur. J. Obstet. Gynaecol. Reprod. Biol. 1998, 81, 149–156. [Google Scholar] [CrossRef]
- Wang, J.; Feng, C.; Liu, T.; Shi, M.; Wu, G.; Bazer, F.W. Physiological alterations associated with intrauterine growth restriction in fetal pigs: Causes and insights for nutritional optimization. Mol. Reprod. Dev. 2017, 84, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Świątkiewicz, M.; Księżak, J.; Hanczakowska, E. The effect of feeding native faba bean seeds (Vicia faba L.) to sows and supplemented with enzymes to piglets and growing pigs. Ann. Anim. Sci. 2018, 18, 1007–1027. [Google Scholar] [CrossRef]
- Nowak, P.; Kasprowicz-Potocka, M.; Zaworska, A.; Nowak, W.; Stefańska, B.; Sip, A.; Grajek, W.; Grajek, K.; Frankiewicz, A. The effect of combined feed additives on growing pigs’ performance and digestive tract parameters. Ann. Anim. Sci. 2019, 19, 807–819. [Google Scholar] [CrossRef]
- Yang, X.F.; Qin, J.F.; Wang, L.; Gao, K.G.; Zheng, C.T.; Huang, L.; Jiang, Z.Y. Improved milk glutamine level and growth performance of suckling piglets by glutamine supplementation in maternal diet. Ann. Anim. Sci. 2018, 18, 441–452. [Google Scholar] [CrossRef]
- Seckl, J.R. Glucocorticoid programming of the fetus; adult phenotypes and molecular mechanisms. Mol. Cell. Endocrinol. 2001, 185, 61–71. [Google Scholar] [CrossRef]
- Xu, R.J.; Mellor, D.J.; Birtles, M.J.; Reynolds, G.W.; Simpson, H.V. Impact of intrauterine growth retardation on the gastrointestinal tract and the pancreas in newborn pigs. J. Pediat. Gastroenterol. Nutr. 1994, 18, 231–240. [Google Scholar] [CrossRef]
- Śliwa, E. Effect of simultaneous versus apart administration of dexamethasone and alpha-ketoglutarate on growth hormone, cortisol and insulin-like growth factor-I in piglets. Bull. Vet. Inst. Pulawy 2006, 50, 205–210. [Google Scholar]
- Śliwa, E.; Tatara, M.R.; Nowakowski, H.; Pierzynowski, S.G.; Studziński, T. Effect of maternal dexamethasone and alpha-ketoglutarate administration on skeletal development during the last three weeks of prenatal life in pigs. J. Matern. Fetal. Neonatal Med. 2006, 19, 489–493. [Google Scholar] [CrossRef]
- Śliwa, E.; Dobrowolski, P.; Tatara, M.R.; Piersiak, T.; Siwicki, A.; Rokita, E.; Pierzynowski, S.G. Alpha-ketoglutarate protects the liver of piglets exposed during prenatal life to chronic excess of dexamethasone from metabolic and structural changes. J. Anim. Physiol. Anim. Nutr. 2009, 93, 192–202. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Puzio, I. Postnatal administration of 2-oxoglutaric acid improves the intestinal barrier affected by the prenatal action of dexamethasone in pigs. Nutrition 2012, 28, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Śliwa, E.; Dobrowolski, P.; Piersiak, T. Bone development of suckling piglets after prenatal, neonatal or perinatal treatment with dexamethasone. J. Anim. Physiol. Anim. Nutr. 2010, 94, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Lingas, R.; Dean, F.; Matthews, S.G. Maternal nutrient restriction (48h) modifies brain corticosteroid receptors expression and endocrine function in the fetal guinea pig. Brain Res. 1999, 846, 236–242. [Google Scholar] [CrossRef]
- Gill, J.W.; Hosking, B.J.; Egan, A.R. Prenatal programing of mammalian growth—A review of the role of steroids. Livest. Prod. Sci. 1998, 54, 251–267. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P. Maternal glucocorticoid treatment as a model for examining foetal gender-specific effects on the development that influences the bone metabolism of neonatal piglets. Bull. Vet. Inst. Pulawy 2012, 56, 247–253. [Google Scholar] [CrossRef]
- Śliwa, E.; Kowalik, S.; Tatara, M.R.; Studziński, T. Effect of maternal administration of dexamethasone during last weeks of pregnancy on foetal skeletal system in pigs. Bull. Vet. Inst. Pulawy 2004, 48, 449–452. [Google Scholar]
- Tomaszewska, E.; Dobrowolski, P.; Puzio, I. Morphological changes of the cartilage and bone in newborn piglets evoked by experimentally induced glucocorticoid excess during pregnancy. J. Anim. Physiol. Anim. Nutr. 2013, 97, 785–796. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Siwicki, A.K. Maternal treatment with dexamethasone at minimal therapeutic doses inhibits neonatal bone development in a gender-dependent manner. Livst. Sci. 2012, 146, 175–182. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Wydrych, J. Postnatal administration of 2-oxoglutaric acid improves articular and growth plate cartilages and bone tissue morphology in pigs prenatally treated with dexamethasone. J. Physiol. Pharmacol. 2012, 63, 547–554. [Google Scholar]
- Tomaszewska, E.; Dobrowolski, P.; Bieńko, M.; Prost, Ł.; Szymańczyk, S.; Zdybel, A. Effects of 2-oxoglutaric acid on bone morphometry, densitometry, mechanics, and immunohistochemistry in 9-month-old boars with prenatal dexamethasone-induced osteopenia. Connect. Tissue Res. 2015, 56, 483–492. [Google Scholar] [CrossRef]
- Blicharski, T.; Tomaszewska, E.; Dobrowolski, P.; Hułas-Stasiak, M.; Muszyński, S. A metabolite of leucine (β-hydroxy-β-methylbutyrate) given to sows during pregnancy alters bone development of their newborn offspring by hormonal modulation. PLoS ONE 2017, 12, e0179693. [Google Scholar] [CrossRef] [PubMed]
- Rudyk, H.; Tomaszewska, E.; Kotsyumbas, I.; Muszyński, S.; Tomczyk-Warunek, A.; Szymańczyk, S.; Dobrowolski, P.; Wiącek, D.; Kamiński, D.; Brezvyn, O. Bone homeostasis in experimental fumonisins intoxication of rats. Ann. Anim. Sci. 2019, 19, 403–419. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Świetlicka, I.; Muszyński, S.; Kostro, K.; Jakubczak, A.; Taszkun, I.; Żmuda, A.; Rycerz, K.; Blicharski, T.; et al. Effects of maternal treatment with β-hydroxy-β-metylbutyrate and 2-oxoglutaric acid on femur development in offspring of minks of the standard dark brown type. J. Anim. Physiol. Anim. Nutr. 2018, 102, e299–e308. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, E.; Muszyński, S.; Dobrowolski, P.; Kamiński, D.; Czech, A.; Grela, E.R.; Wiącek, D.; Tomczyk-Warunek, A. Dried fermented post-extraction rapeseed meal given to sows as an alternative protein source for soybean meal during pregnancy improves bone development of their offspring. Livest. Sci. 2019, 224, 60–68. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Muszyński, S.; Dobrowolski, P.; Wiącek, D.; Tomczyk-Warunek, A.; Świetlicka, I.; Pierzynowski, S.G. Maternal HMB treatment affects bone and hyaline cartilage development in their weaned piglets via the leptin/osteoprotegerin system. J. Anim. Physiol. Anim. Nutr. 2019, 103, 626–643. [Google Scholar] [CrossRef] [PubMed]
- Dauncey, M.J.; Bicknell, R.J. Nutrition and neurodevelopment: Mechanisms of developmental dysfunction and disease in later life. Nutr. Res. Rev. 1999, 12, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Neu, J.; Hauser, N.; Douglas-Escobar, M. Postnatal nutrition and adult health programming. Semin. Fetal Neonat. Med. 2007, 12, 78–86. [Google Scholar] [CrossRef]
- Dobrowolski, P.; Tomaszewska, E.; Muszyński, S.; Blicharski, T.; Pierzynowski, S.G. Dietary 2-oxoglutarate prevents bone loss caused by neonatal treatment with maximal dexamethasone dose. Exp. Biol. Med. 2017, 242, 671–682. [Google Scholar] [CrossRef]
- Tomaszewska, E.; Dobrowolski, P.; Prost, Ł.; Hułas-Stasiak, M.; Muszyński, S.; Blicharski, T. The effect of supplementation of glutamine precursor on the growth plate, articular cartilage and cancellous bone in fundectomy-induces osteopenic bone. J. Vet. Med. Sci. 2016, 78, 563–571. [Google Scholar] [CrossRef]
- Kowalik, S.; Śliwa, E.; Tatara, M.R.; Krupski, W.; Majcher, P.; Studziński, T. Influence of alpha-ketoglutarate (AKG) on mineral density and geometrical and mechanical parameters of femora during postnatal life in piglets. Bull. Vet. Inst. Pulawy 2005, 49, 107–111. [Google Scholar]
- Kristensen, N.B.; Jungvid, H.; Fernández, J.A.; Pierzynowski, S.G. Absorption and metabolism of alpha-ketoglutarate in growing pigs. J. Anim. Physiol. Anim. Nutr. 2002, 86, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolski, P.; Tomaszewska, E.; Radzki, R.P.; Bienko, M.; Wydrych, J.; Zdybel, A.; Pierzynowski, S.G. Can 2-oxoglutarate prevent changes in bone evoked by omeprazole? Nutrition 2013, 29, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Śliwa, E.; Kowalik, S.; Tatara, M.R.; Krupski, W.; Majcher, P.; Łuszczewska-Sierakowska, I.; Pierzynowski, S.G.; Studziński, T. Effect of alpha-ketoglutarate (AKG) given to pregnant sows on development of humerus and femur in newborns. Bull. Vet. Inst. Pulawy 2005, 49, 117–120. [Google Scholar]
- Śliwa, E.; Dobrowolski, P.; Tatara, M.R.; Pierzynowski, S.G. Alpha-ketoglutarate partially protects newborns from metabolic changes evoked by chronic maternal exposure to glucocorticoids. J. Pre Clin. Clin. Res. 2007, 1, 55–59. [Google Scholar]
- Patti, A.; Gennari, L.; Merlotti, D.; Dotta, F.; Nuti, R. Endocrine action of osteocalcin. Int. J. Endocrinol. 2013, 846480. [Google Scholar] [CrossRef]
- Tajan, M.; Hock, A.H.; Blagih, J.; Robertson, N.A.; Labuschage, C.F.; Kruiswijk, F.; Humpton, T.J.; Adams, P.D.; Vousden, K.H. A role for p53 in the adaptation to glutamine starvation through the expression of SLC1A3. Cell Metabolism 2018, 28, 721–736. [Google Scholar] [CrossRef]
- Grela, E.R.; Skomiał, J. Nutritional Recommendations and Nutritional Value of Feed for Pigs, 2nd ed.; Institute of Physiology and Animal Nutrition of Polish Academy of Science: Jabłonna, Poland, 2015; pp. 1–95. [Google Scholar]
- Śliwa, E.; Tatara, M.R.; Pierzynowski, S.G. Total cholesterol, glucose and electrolytes in piglets serum after alpha-ketoglutarate (AKG) and dexamethasone treatment during prenatal and neonatal life. Bull. Vet. Inst. Pulawy 2006, 50, 561–566. [Google Scholar]
- Tomaszewska, E.; Dobrowolski, P.; Kwiecień, M.; Winiarska-Mieczan, A.; Tomczyk, A.; Muszyński, S.; Gładyszewska, B. Dose-dependent influence of dietary Cu-glycine complex on bone and hyaline cartilage development in adolescent rats. Ann. Anim. Sci. 2017, 17, 1089–1105. [Google Scholar] [CrossRef]
- Muszyński, S.; Kwiecień, M.; Tomaszewska, E.; Świetlicka, I.; Dobrowolski, P.; Kasperek, K.; Jeżewska-Witkowska, G. Effect of caponization on performance and quality characteristics of long bones in Polbar chickens. Poult. Sci. 2017, 96, 491–500. [Google Scholar] [CrossRef]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirus staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Camplejohn, K.L.; Allard, S.A. Limitations of safranin ‘O’ staining in proteoglycan-depleted cartilage demonstrated with monoclonal antibodies. Histochemistry 1988, 89, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Rich, L.; Whittaker, P. Collagen and Picrosirius Red staining: A polarized light assessment of fibrillar hue and spatial distribution. Braz. J. Morphol. Sci. 2005, 22, 97–104. [Google Scholar]
- Pearle, A.D.; Warren, R.F.; Rodeo, S.A. Basic science of articular cartilage and osteoarthritis. Clin. J. Sport. Med. 2005, 24, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, Z. Clinical physiology and pathology of the growth plate. Best Pract. Res. Clin. Endocrinol. Metab. 2002, 16, 399–419. [Google Scholar] [CrossRef]
- Wikström, B.; Hjerpe, A.; Hultenby, K.; Reinholt, F.; Engfeldt, B. Stereological analysis of the epiphyseal growth cartilage in the brachymorphic (bm/bm) mouse, with special reference to the distribution of matrix vesicles. Virchows Arch. B Cell Pathol. 1984, 47, 199–210. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Tatara, M.R.; Brodzki, A.; Krupski, W.; Śliwa, E.; Silmanowicz, P.; Majcher, P.; Pierzynowski, S.G.; Studziński, T. Effect of alpha-ketoglutarate (AKG) on bone homeostasis and plasma amino acids in turkeys. Poult. Sci. 2005, 84, 1604–1609. [Google Scholar] [CrossRef]
- Wu, N.; Yang, M.; Gaur, U.; Xu, H.; Yao, Y.; Li, D. Alpha-ketoglutarate: Physiological functions and applications. Biomol. Ther. 2016, 24, 1–8. [Google Scholar] [CrossRef]
- Harrison, A.P.; Pierzynowski, S.G. Biological effects of 2-oxoglutarate with particular emphasis on the regulation of protein, mineral and lipid absorption/metabolism, muscle performance, kidney function, bone formation and cancerogenesis, all viewed from a healthy ageing perspective state of the art: Review article. J. Physiol. Pharmacol. 2008, 59 (Suppl. 1), 91–106. [Google Scholar]
- Śliwa, E.; Kowalik, S.; Tatara, M.R.; Majcher, P.; Krupski, W.; Studziński, T. Effects of dexamethasone on physical properties and mineral density of long bones in piglets. Bull. Vet. Inst. Pulawy 2005, 49, 97–100. [Google Scholar]
- Śliwa, E.; Tatara, M.R.; Dudek, K.; Siwicki, A.K.; Kowalik, S.; Pierzynowski, S.G.; Studziński, T. Influence of alpha-ketoglutarate administered to pregnant sows on nonspecific and specific humoral defense mechanism in newborn piglets and during two weeks of neonatal life. Pol. J. Environ. Stud. 2005, 14 (Suppl. II), 356–359. [Google Scholar]
- Newsholme, P.; Lima, M.M.R.; Procopio, J.; Pithon-Curi, T.C.; Doi, S.Q.; Bazotte, R.B.; Curi, R. Glutamine and glutamate as vital metabolites. Braz. J. Med. Biol. Res. 2003, 36, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Zhang, X.H.; Li, X.M.; Zhou, Y.M.; Li, W.; Huang, X.X.; Zhang, L.L.; Wang, T. Intestinal growth and morphology is associated with the increase in heat shock protein 70 expression in weaning piglets through supplementation with glutamine. J. Anim. Sci. 2001, 89, 3634–3642. [Google Scholar] [CrossRef]
- Hanczakowska, E.; Niwinska, B.; Grela, E.R.; Węglarzy, K.; Okoń, K. Effect of dietary glutamine, glucose and/or sodium butyrate on piglet growth, intestinal environment, subsequent fattener performance, and meat quality. Czech J. Anim. Sci. 2014, 59, 460–470. [Google Scholar] [CrossRef]
- Hanczakowska, E.; Świątkiewicz, M.; Natonek-Wiśniewska, M.; Okoń, K. Effect of glutamine and/or probiotic (Enterococcus faecium) feed supplementation on piglet performance, intestines structure, and antibacterial activity. Czech J. Anim. Sci. 2017, 62, 313–322. [Google Scholar] [CrossRef]
- Ruzzo, E.K.; Capo-Chichi, J.M.; Ben-Zeev, B.; Chitayat, D.; Mao, H.; Pappas, A.L.; Hitomi, Y.; Lu, Y.F.; Yao, X.; Hamdan, F.F.; et al. Deficiency of asparagine synthetase causes congenital microcephaly and a progressive form of encephalopathy. Neuron 2013, 80, 429–441. [Google Scholar] [CrossRef]
- Meyer, T.; Gustafsson, J.-Å.; Carlstedt-Duke, J. Glucocorticoid-dependent transcriptional repression of the osteocalcin gene by competitive binding at the TATA box. DNA Cell Biol. 1997, 8, 919–927. [Google Scholar] [CrossRef]
- Lee, A.J.; Hodges, S.; Eastell, R. Measurement of osteocalcin. Ann. Clin. Biochem. 2000, 37, 432–446. [Google Scholar] [CrossRef]
- Neve, A.; Corrado, A.; Cantatore, F.P. Osteoblast physiology in normal and pathological condition. Cell Tissue Res. 2011, 342, 289–302. [Google Scholar] [CrossRef]
- Blair, J.M.; Zheng, Y.; Dunstan, C.R. RANK ligand. Int. J. Biochem. Cell Biol. 2007, 39, 1077–1081. [Google Scholar] [CrossRef]
- Deng, X.; He, G.; Levine, A.; Cao, Y.; Mullins, C. Adenovirus-mediated expression of TIMP-1 and TIMP-2 in bone inhibits osteolytic degradation by human prostate cancer. Int. J. Cancer 2008, 122, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [PubMed]
- Lawson, K.A.; Dunn, N.R.; Roelen, B.A.; Zeinstra, L.M.; Davis, A.M.; Wright, C.V.; Korving, J.P.; Hogan, B.L. BMP4 is required for the generation of primordial germ cells in the mouse embryo. Genes Dev. 1999, 13, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Yu, Y.; Chen, Q.; Li, C.; Wu, H.; Wan, Y.; Ma, J.; Sun, F. Soc9, a key transcription factor of bone morphogenetic protein–2–induced chondrogenesis, is activated through BMP pathway and a CCAAT box in the proximal promoter. J. Cell. Physiol. 2008, 217, 228–241. [Google Scholar] [CrossRef]
- Zehentner, B.K.; Dony, C.; Burtscher, H. The transcription factor Sox9 is involved in BMP–2 signaling. J. Bone Miner. Res. 1999, 14, 1734–1741. [Google Scholar] [CrossRef]
- Hu, K.; Olsen, B.R. Osteoblast–derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J. Clin. Invest. 2016, 26, 509–526. [Google Scholar] [CrossRef]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell … and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef]
- Athanasiou, K.A.; Darling, E.M.; Duraine, G.D.; Hu, J.C.; Reddi, A.H. Articular cartilage tissue engineering. Synth. Lect. Tissue Eng. 2009, 1, 1–182. [Google Scholar] [CrossRef]
- Moutsatsou, P.; Kassi, E.; Papavassiliou, A.G. Glucocorticoid receptor signaling in bone cells. Trends Mol. Med. 2012, 18, 348–359. [Google Scholar] [CrossRef]
- Abcouwer, S.F.; Marjon, P.L.; Loper, R.K.; Vander Jagt, D.L. Response of VEGF expression to amino acid deprivation and inducers of endoplasmic reticulum stress. Invest. Ophthalmol. Vis. Sci. 2002, 43, 2791–2798. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content, % |
|---|---|
| Wheat | 19.00 |
| Barley | 23.30 |
| Corn | 20.00 |
| Soybean meal | 17.00 |
| Wheat bran | 8.00 |
| Dried alfalfa | 6.00 |
| Rapeseed oil | 4.00 |
| Calcium carbonate | 1.00 |
| Calcium phosphate | 0.60 |
| Salt | 0.35 |
| Vitamin-mineral premix | 0.50 |
| L-Lysine | 0.20 |
| DL-Methionine | 0.028 |
| L-Threonine | 0.03 |
| Ingredients | Content, % | |
|---|---|---|
| Piglets (until 59th Day) | Experimental Gilts (from 60th Day) | |
| Wheat | 47.11 | 30.10 |
| Barley | - | 29.00 |
| Corn | 15.00 | 10.00 |
| Soybean meal | 18.00 | 20.00 |
| Wheat bran | - | 6.00 |
| Skim milk powder | 10.00 | - |
| Dried whey | 5.00 | - |
| Rapeseed oil | 2.00 | 2.00 |
| Calcium carbonate | 1.10 | 1.30 |
| Calcium phosphate | 0.60 | 0.50 |
| Salt | 0.13 | 0.28 |
| Vitamin-mineral premix | 0.50 | 0.50 |
| L-lysine | 0.30 | 0.20 |
| DL-methionine | 0.14 | 0.06 |
| L-threonine | 0.12 | 0.06 |
| Analysed Fragment | Control | 2-Ox | p-Value |
|---|---|---|---|
| Epiphysieal trabeculae | 1.39 ± 0.75 | 1.45 ± 0.59 | 0.797 |
| Metaphyseal trabeculae | 29.2 ± 10.4 | 20.2 ± 8.9 | 0.005 |
| Articular cartilage | 1.54 ± 0.83 | 1.46 ± 1.19 | 0.721 |
| Parameter | Control | 2-Ox | p-Value |
|---|---|---|---|
| Cholesterol, mmol/L | 1.61 ± 0.13 | 1.34 ± 0.17 | 0.012 |
| TG, mmol/L | 0.251 ± 0.056 | 0.255 ± 0.045 | 0.422 |
| HDL, mmol/L | 0.095 ± 0.014 | 0.101 ± 0.006 | 0.424 |
| LDL, mmol/L | 1.45 ± 0.17 | 1.11 ± 0.16 | 0.005 |
| Urea, mmol/L | 5.12 ± 0.58 | 5.64 ± 0.20 | 0.059 |
| Creatinine, mmol/L | 169.4 ± 34.2 | 173.8 ± 34.3 | 0.828 |
| Uric acid, mmol/L | 0.055 ± 0.013 | 0.060 ± 0.024 | 0.691 |
| Albumin, g/L | 40.3 ± 3.4 | 39.0 ± 1.7 | 0.387 |
| ALT, U/L | 33.8 ± 7.4 | 28.7 ± 9.3 | 0.313 |
| AST, U/L | 52.5 ± 12.7 | 42.0 ± 14.9 | 0.218 |
| LDH, U/L | 2818.5 ± 447.3 | 2635.7 ± 638.8 | 0.578 |
| ALP, U/L | 186.8 ± 50.4 | 223.5 ± 29.6 | 0.155 |
| Fe, mmol/L | 10.7 ± 0.6 | 11.4 ± 0.7 | 0.109 |
| Mg, mmol/L | 1.11 ± 0.06 | 1.21 ± 0.11 | 0.173 |
| P, mmol/L | 2.25 ± 0.50 | 2.33 ± 0.43 | 0.751 |
| Ca, mmol/L | 2.72 ± 0.68 | 2.80 ± 0.77 | 0.847 |
| IGF-1, ng/mL | 77.8 ± 19.7 | 92.6 ± 23.8 | 0.268 |
| Amino Acid (nmol/mL) | Control | 2-Ox | p-Value |
|---|---|---|---|
| Arginine | 130.3 ± 19.1 | 123.0 ± 22.4 | 0.555 |
| Aspartic acid | 11.0 ± 0.9 | 13.7 ± 2.2 | 0.019 |
| Glutamine | 234.7 ± 19.0 | 313.0 ± 101.5 | 0.093 |
| Glutamic acid | 94.0 ± 9.7 | 147.2 ± 22.8 | 0.005 |
| Glycine | 1186.7 ± 331.2 | 736.3 ± 95.6 | 0.066 |
| Histidine | 115.3 ± 17.5 | 108.5 ± 31.7 | 0.654 |
| Isoleucine | 125.2 ± 23.3 | 81.3 ± 21.4 | 0.007 |
| Leucine | 265.3 ± 44.4 | 205.5 ± 30.4 | 0.021 |
| Lysine | 276.3 ± 43.2 | 297.7 ± 59.6 | 0.494 |
| Methionine | 41.5 ± 7.2 | 32.2 ± 9.3 | 0.080 |
| Ornithine | 94.8 ± 10.0 | 116.7 ± 27.3 | 0.230 |
| Proline | 1018.7 ± 94.3 | 600.0 ± 118.7 | <0.001 |
| Serine | 165.0 ± 35.8 | 132.0 ± 39.6 | 0.066 |
| Threonine | 187.2 ± 49.3 | 124.5 ± 52.0 | 0.058 |
| Tryptophan | 50.0 ± 9.2 | 52.5 ± 20.7 | 0.792 |
| Valine | 396.3 ± 80.8 | 299.2 ± 76.5 | 0.149 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomaszewska, E.; Dobrowolski, P.; Świątkiewicz, M.; Donaldson, J.; Puzio, I.; Muszyński, S. Is Dietary 2-Oxoglutaric Acid Effective in Accelerating Bone Growth and Development in Experimentally-Induced Intrauterine Growth Retarded Gilts? Animals 2020, 10, 728. https://doi.org/10.3390/ani10040728
Tomaszewska E, Dobrowolski P, Świątkiewicz M, Donaldson J, Puzio I, Muszyński S. Is Dietary 2-Oxoglutaric Acid Effective in Accelerating Bone Growth and Development in Experimentally-Induced Intrauterine Growth Retarded Gilts? Animals. 2020; 10(4):728. https://doi.org/10.3390/ani10040728
Chicago/Turabian StyleTomaszewska, Ewa, Piotr Dobrowolski, Małgorzata Świątkiewicz, Janine Donaldson, Iwona Puzio, and Siemowit Muszyński. 2020. "Is Dietary 2-Oxoglutaric Acid Effective in Accelerating Bone Growth and Development in Experimentally-Induced Intrauterine Growth Retarded Gilts?" Animals 10, no. 4: 728. https://doi.org/10.3390/ani10040728
APA StyleTomaszewska, E., Dobrowolski, P., Świątkiewicz, M., Donaldson, J., Puzio, I., & Muszyński, S. (2020). Is Dietary 2-Oxoglutaric Acid Effective in Accelerating Bone Growth and Development in Experimentally-Induced Intrauterine Growth Retarded Gilts? Animals, 10(4), 728. https://doi.org/10.3390/ani10040728