Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets

 ,
,  ,
,  ,
,  , ,
, , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. The In Vivo Trial
2.2. Zootechnical Evaluation
2.3. Protein Content in Faecal Samples
2.4. Blood Sample Collection and Biochemical Analyses
2.5. Insulin and Leptin Evaluation by Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Hair Cortisol Extraction and Assay
2.7. DNA Extraction and Real-Time Polymerase Chain Reaction (PCR) to Determine Gut Microbiota
2.8. Statistical Analysis
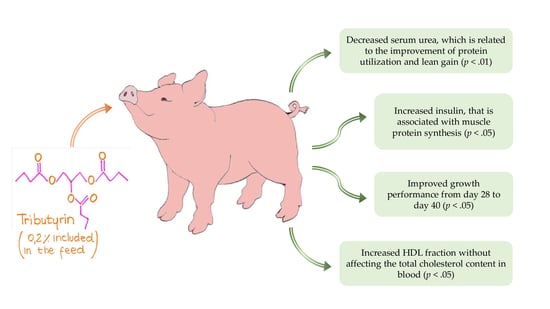
3. Results
3.1. Zootechnical Evaluation
3.2. Protein Content in Faecal Samples
3.3. Blood Sample Collection and Biochemical Analyses
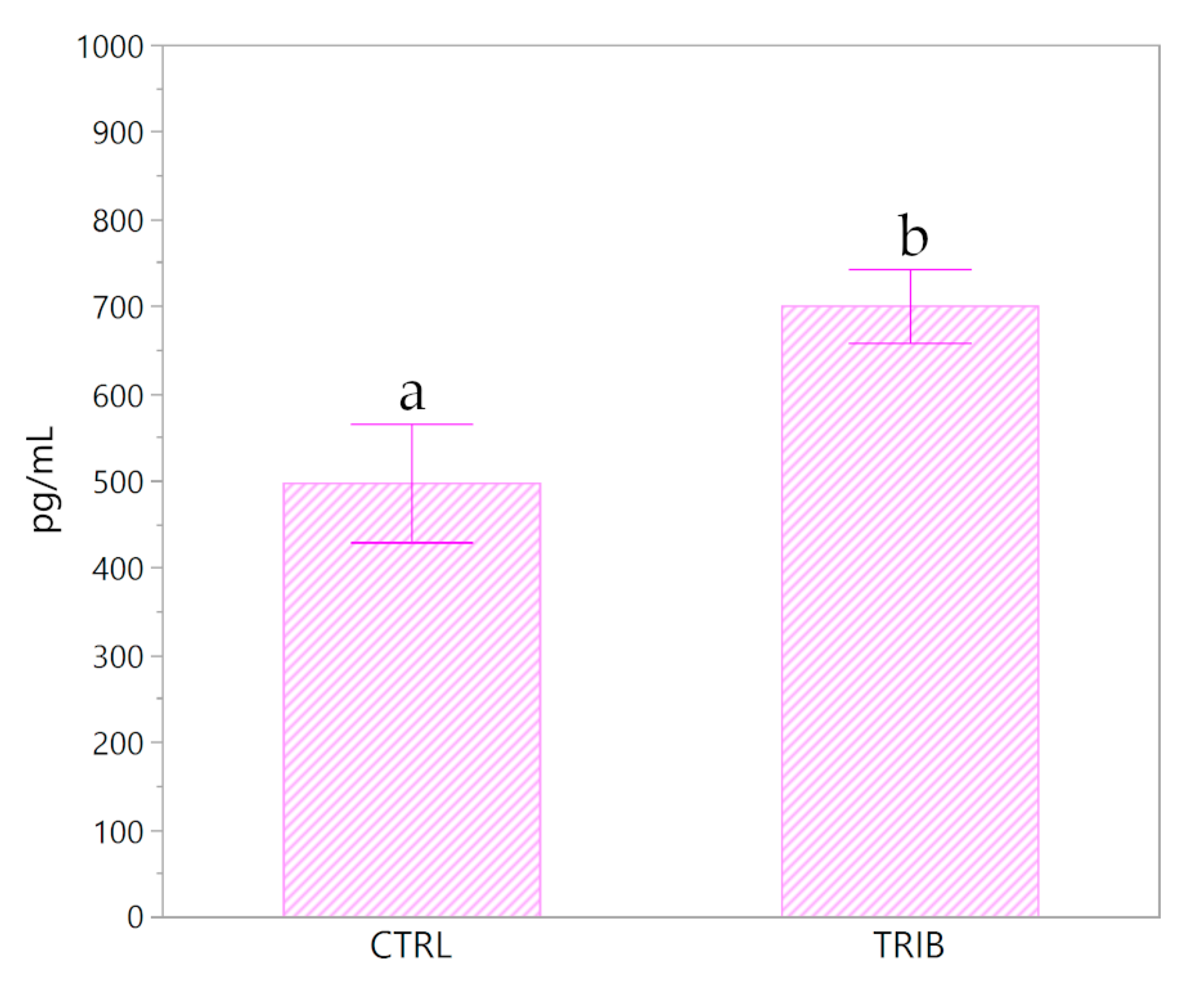
3.4. Insulin and Leptin Evaluation
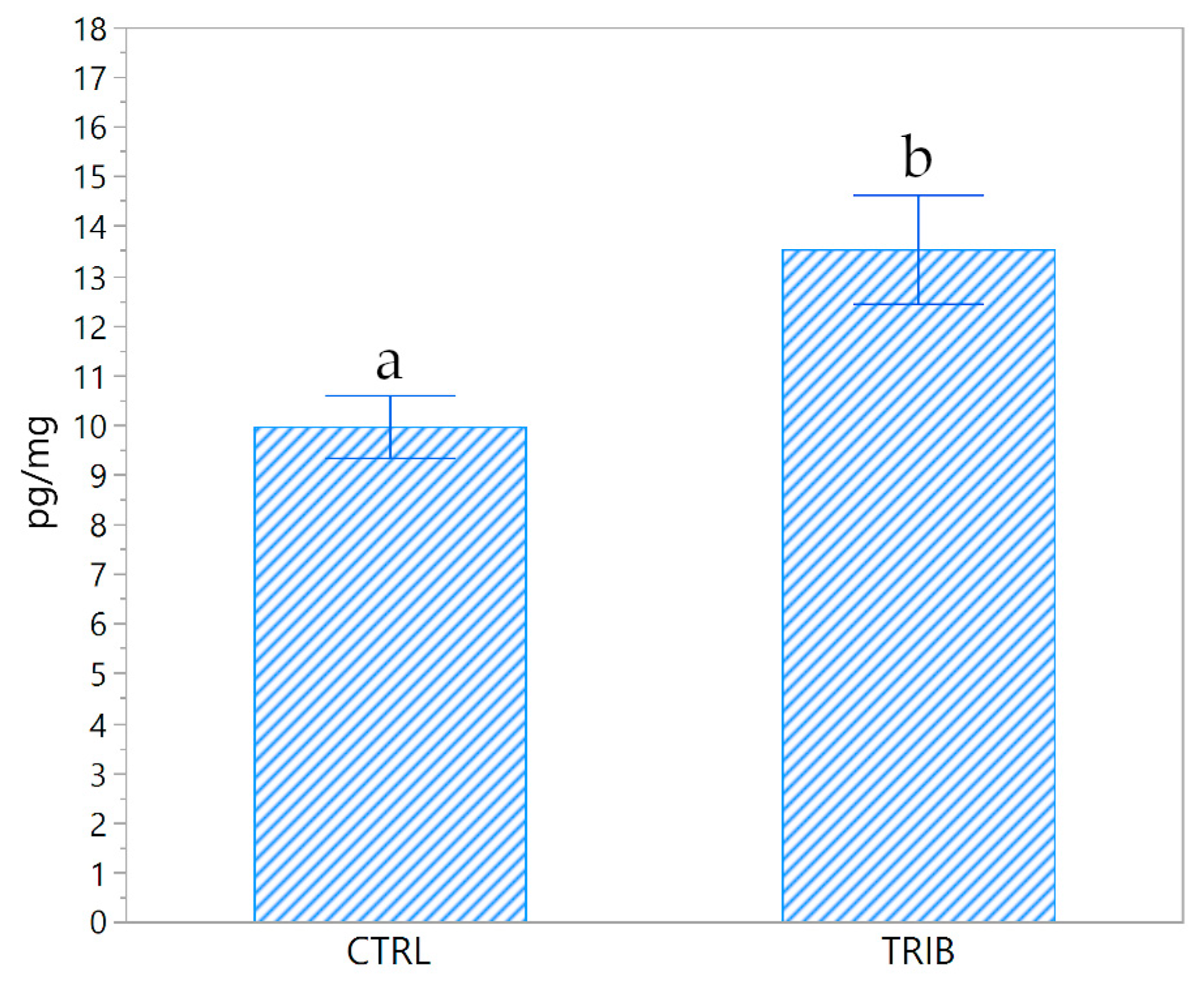
3.5. Cortisol Concentration
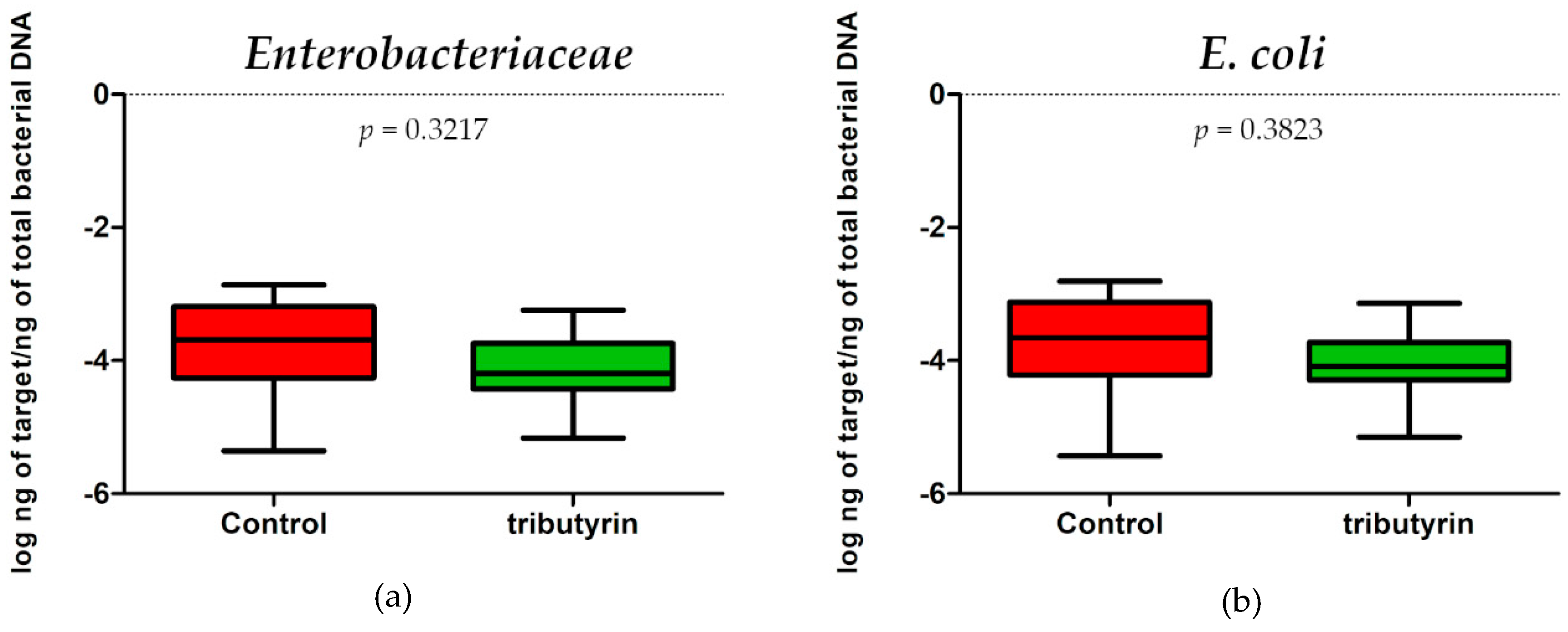
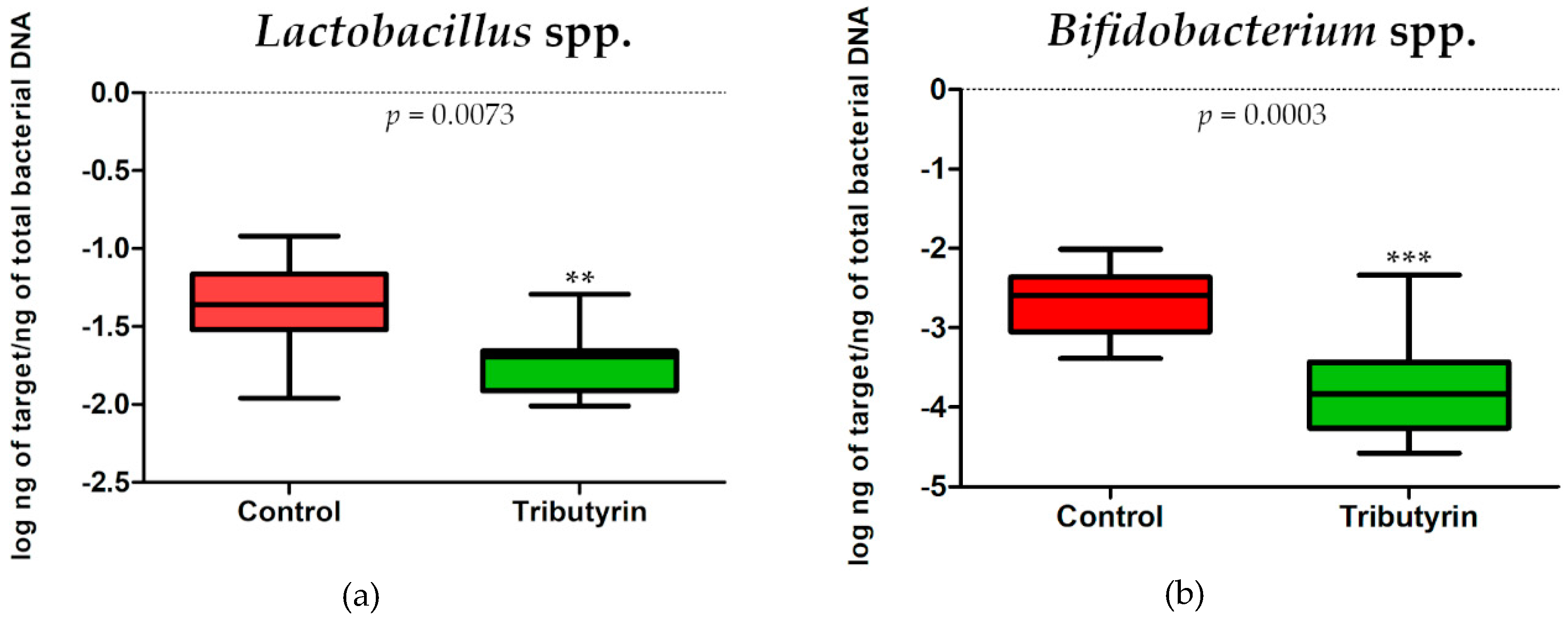
3.6. DNA Extraction and Real-Time PCR to Determine Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rossi, L.; Vagni, S.; Polidori, C.; Alborali, G.L.; Baldi, A.; Dell’Orto, V. Experimental induction of Escherichia coli diarrhoea in weaned piglets. Open. J. Vet. Med. 2012, 2, 1–8. [Google Scholar] [CrossRef]
- Burow, E.; Rostalski, A.; Harlizius, J.; Gangl, A.; Simoneit, C.; Grobbel, M.; Käsbohrer, A. Antibiotic resistance in Escherichia coli from pigs from birth to slaughter and its association with antibiotic treatment. Prev. Vet. Med. 2019, 165, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Barba-Vidal, E.; Martín-Orúe, S.M.; Castillejos, L. Practical aspects of the use of probiotics in pig production: A review. Livest. Sci. 2019, 223, 84–96. [Google Scholar] [CrossRef]
- Azagra-Boronat, I.; Rodríguez-Lagunas, M.J.; Castell, M.; Pérez-Cano, F.J. Prebiotics for gastrointestinal infections and acute diarrhea. In Dietary Interventions in Gastrointestinal Diseases; Watson, R.R., Preedy, V.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 179–191. [Google Scholar]
- Yang, C.; Zhang, L.; Cao, G.; Feng, J.; Yue, M.; Xu, Y.; Guo, X. Effects of dietary supplementation with essential oils and organic acids on the growth performance, immune system, fecal volatile fatty acids, and microflora community in weaned piglets. J. Anim. Sci. 2019, 97, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.J.; Kim, D.W.; Kang, B.N.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef]
- Cuff, M.; Dyer, J.; Jones, M.; Shirazi-Beechey, S. The human colonic monocarboxylate transporter Isoform 1: Its potential importance to colonic tissue homeostasis. Gastroenterology 2005, 128, 676–686. [Google Scholar] [CrossRef]
- Mátis, G.; Kulcsár, A.; Petrilla, J.; Talapka, P.; Neogrády, Z. Porcine hepatocyte-Kupffer cell co-culture as an in vitro model for testing the efficacy of anti-inflammatory substances. J. Anim. Physiol. Anim. Nutr. 2017, 101, 201–207. [Google Scholar] [CrossRef]
- Salsali, H.; Parker, W.J.; Sattar, S.A. The effect of volatile fatty acids on the inactivation of Clostridium perfringens in anaerobic digestion. World J. Microb. Biotechnol. 2008, 24, 659–665. [Google Scholar] [CrossRef]
- Mazzoni, M.; Le Gall, M.; De Filippi, S.; Minieri, L.; Trevisi, P.; Wolinski, J.; Bosi, P. Supplemental sodium butyrate stimulates different gastric cells in weaned pigs. J. Nutr. 2008, 138, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- Machinsky, T.G.; Kessler, A.D.M.; Ribeiro, A.M.L.; Moraes, M.D.L.; Silva, I.C.M.D.; Cortés, M.E.M. Nutrient digestibility and Ca and P balance in pigs receiving butyric acid, phytase and different calcium levels. Cienc. Rural 2010, 40, 2350–2355. [Google Scholar] [CrossRef]
- Bedford, A.; Gong, J. Implications of butyrate and its derivatives for gut health and animal production. Anim. Nutr. 2018, 4, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Biagi, G.; Piva, A.; Moschini, M.; Vezzali, E.; Roth, F.X. Performance, intestinal microflora, and wall morphology of weanling pigs fed sodium butyrate. J. Anim. Sci. 2007, 85, 1184–1191. [Google Scholar] [CrossRef]
- Sakdee, J.; Poeikhamph, T.; Rakangthon, C.; Poungpong, K.; Bunchasak, C. Effect of tributyrin supplementation in diet on production performance and gastrointestinal tract of healthy nursery pigs. Pak. J. Nutr. 2016, 15, 954–962. [Google Scholar] [CrossRef]
- Dong, L.; Zhong, X.; He, J.; Zhang, L.; Bai, K.; Xu, W.; Huang, X. Supplementation of tributyrin improves the growth and intestinal digestive and barrier functions in intrauterine growth-restricted piglets. Clin. Nutr. 2017, 35, 399–407. [Google Scholar] [CrossRef]
- Song, Y.; Gu, Y.; Yin, H.; Lin, S.; Zhang, X.; Che, L.; Fang, Z. Dietary supplementation with tributyrin prevented weaned pigs from growth retardation and lethal infection via modulation of inflammatory cytokines production, ileal FGF19 expression, and intestinal acetate fermentation. J. Anim. Sci. 2018, 95, 226–238. [Google Scholar] [CrossRef]
- NCR. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Official Methods of Analysis 2005; AOAC International: Washington, DC, USA, 2005. [Google Scholar]
- Casal, N.; Manteca, X.; Peña, R.; Bassols, A.; Fàbrega, E. Analysis of cortisol in hair samples as an indicator of stress in pigs. J. Vet. Behav. 2017, 19, 1–6. [Google Scholar] [CrossRef]
- Burnett, T.A.; Madureira, A.M.L.; Silper, B.F.; Nadalin, A.; Tahmasbi, A.M.; Veira, D.M.; Cerri, R.L.A. Short communication: Factors affecting hair cortisol concentration in lactating dairy cows. J. Dairy Sci. 2014, 97, 7685–7690. [Google Scholar] [CrossRef]
- Koren, L.; Mokady, O.; Karaskov, T.; Klein, J.; Koren, G.; Geffen, E. A novel method using hair for determining hormonal levels in wildlife. Anim. Behav. 2002, 63, 403–406. [Google Scholar] [CrossRef]
- Patrone, V.; Minuti, A.; Lizier, M.; Miragoli, F.; Lucchini, F.; Trevisi, E.; Rossi, F.; Callegari, M.L. Differential effects of coconut versus soy oil on gut microbiota composition and predicted metabolic function in adult mice. BMC Genom. 2018, 19, 808. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Vink, C.; Driessen, C.; London, N.; Thijs, C.; Stobberingh, E.E. Quantification of Bifidobacterium spp., Escherichia coli and Clostridium difficile in faecal samples of breast-fed and formula-fed infants by real-time PCR. FEMS Microbiol. Lett. 2005, 243, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, S.; Fite, A.; Macfarlan, G.T.; McMurdo, M.E. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl. Environ. Microbiol. 2004, 70, 3575–3581. [Google Scholar] [CrossRef] [PubMed]
- Byun, R.; Nadkarni, M.A.; Chhour, K.L.; Martin, F.E.; Jacques, N.A.; Hunter, N. Quantitative analysis of diverse Lactobacillus species present in advanced dental caries. J. Clin. Microbiol. 2004, 42, 3128–3136. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.Q.; Liu, Y.L.; Hu, J.; Shen, W.H. Effects of lactitol and tributyrin on growth performance, small intestinal morphology and enzyme activity in weaned pigs. Asian-Australas. J. Anim. 2006, 19, 1470–1477. [Google Scholar] [CrossRef]
- Murray, R.L. Tributyrin, a Butyrate Pro-Drug, as a Muscle Growth Promoter in a Porcine Model. Ph.D. Thesis, University of Maryland, College Park, MD, USA, 2018; p. 164. [Google Scholar]
- Kotunia, A.; Wolinski, J.; Laubitz, D.; Jurkowska, M.; Rome, V.; Guilloteau, P.; Zabielski, R. Effect of sodium butyrate on the small intestine. J. Physiol. Pharmacol. 2004, 55, 59–68. [Google Scholar]
- Liu, Y.; Xiong, X.; Tan, B.E.; Song, M.; Ji, P.; Kim, K.; Yin, Y. Nutritional intervention for the intestinal development and health of weaned pigs. Front. Vet. 2019, 6, 46. [Google Scholar] [CrossRef]
- Tugnoli, B.; Giovagnoni, G.; Piva, A.; Grilli, E. From Acidifiers to Intestinal Health Enhancers: How Organic Acids Can Improve Growth Efficiency of Pigs. Animals 2020, 10, 134. [Google Scholar] [CrossRef]
- Piva, A.; Morlacchini, M.; Casadei, G.; Gatta, P.P.; Biagi, G.; Prandini, A. Sodium butyrate improves growth performance of weaned piglets during the first period after weaning. Ital. J. Anim. Sci. 2002, 1, 35–41. [Google Scholar] [CrossRef]
- Lallès, J.P.; Bosi, P.; Janczyk, P.; Koopmans, S.J.; Torrallardona, D. Impact of bioactive substances on the gastrointestinal tract and performance of weaned piglets: A review. Animal 2009, 3, 1625–1643. [Google Scholar] [CrossRef]
- Etim, N.N.; Offiong, E.E.; Williams, M.E.; Asuquo, L.E. Influence of nutrition on blood parameters of pigs. Am. J. Biol. Life Sci. 2014, 2, 46–52. [Google Scholar]
- Manell, E.; Hedenqvist, P.; Svensson, A.; Jensen-Waern, M. Establishment of a refined oral glucose tolerance test in pigs, and assessment of insulin, glucagon and glucagon-like peptide-1 responses. PLoS ONE 2016, 11, e0148896. [Google Scholar] [CrossRef] [PubMed]
- Davie, J.R. Inhibition of histone deacetylase activity by butyrate. J. Nutr. 2003, 133, 2485S–2493S. [Google Scholar] [CrossRef]
- Christensen, D.P.; Dahllöf, M.; Lundh, M.; Rasmussen, D.N.; Nielsen, M.D.; Billestrup, N.; Mandrup-Poulsen, T. Histone deacetylase (HDAC) inhibition as a novel treatment for diabetes mellitus. Mol. Med. 2011, 17, 378. [Google Scholar] [CrossRef] [PubMed]
- Kawada, Y.; Asahara, S.I.; Sugiura, Y.; Sato, A.; Furubayashi, A.; Kawamura, M.; Koyanagi-Kimura, M. Histone deacetylase regulates insulin signaling via two pathways in pancreatic β cells. PLoS ONE 2017, 12, e0184435. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Jena, G.B. Protective role of sodium butyrate, a HDAC inhibitor on beta-cell proliferation, function and glucose homeostasis through modulation of p38/ERK MAPK and apoptotic pathways: Study in juvenile diabetic rat. Chem. Biol. Interact. 2014, 213, 1–12. [Google Scholar] [CrossRef]
- Podrez, E.A. Anti-oxidant properties of high-density lipoprotein and atherosclerosis. Clin. Exp. Pharmacol. Physiol. 2010, 37, 719–725. [Google Scholar] [CrossRef]
- Navab, M.; Reddy, S.T.; Van Lenten, B.J.; Fogelman, A.M. HDL and cardiovascular disease: Atherogenic and atheroprotective mechanisms. Nat. Rev. Cardiol. 2011, 8, 222. [Google Scholar] [CrossRef]
- Nazih, H.; Nazih-Sanderson, F.; Krempf, M.; Michel Huvelin, J.; Mercier, S.; Marie Bard, J. Butyrate stimulates ApoA-IV-containing lipoprotein secretion in differentiated Caco-2 cells: Role in cholesterol efflux. J. Cell. Biochem. 2001, 83, 230–238. [Google Scholar] [CrossRef]
- Duka, A.; Fotakis, P.; Georgiadou, D.; Kateifides, A.; Tzavlaki, K.; von Eckardstein, L.; Zannis, V.I. ApoA-IV promotes the biogenesis of apoA-IV-containing HDL particles with the participation of ABCA1 and LCAT. J. Lipid Res. 2013, 54, 107–115. [Google Scholar] [CrossRef]
- Xiong, J.; Qiu, H.; Bi, Y.; Zhou, H.L.; Guo, S.; Ding, B. Effects of Dietary Supplementation with Tributyrin and Coated Sodium Butyrate on Intestinal Morphology, Disaccharidase Activity and Intramuscular Fat of Lipopolysaccharide-Challenged Broilers. Braz. J. Poult. Sci. 2018, 20, 707–716. [Google Scholar] [CrossRef]
- Dvorák, M. Changes in blood protein levels in piglets during development and during stress. Vet. Med. Czech 1986, 31, 403–414. [Google Scholar]
- Fuhrman, M.P.; Charney, P.; Mueller, C.M. Hepatic proteins and nutrition assessment. J. Am. Diet. Assoc. 2004, 104, 1258–1264. [Google Scholar] [CrossRef]
- Doornenbal, H.; Tong, A.K.W.; Sather, A.P. Relationships among serum characteristics and performance and carcass traits in growing pigs. J. Anim. Sci. 1986, 62, 1675–1681. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Counotte, G.H.M.; Tielen, M.J.M. Haematological and clinicochemical blood profiles in slaughter pigs: Reference values, between and within herd variability and the relationship with herd factors and growth performance. Vet. Q. 1992, 14, 57–62. [Google Scholar] [CrossRef]
- Liang, T.J.; Grieninger, G. Direct effect of insulin on the synthesis of specific plasma proteins: Biphasic response of hepatocytes cultured in serum and hormone free medium. Proc. Natl. Acad. Sci. USA 1981, 78, 6972–6976. [Google Scholar] [CrossRef]
- Chen, Q.; Lu, M.; Monks, B.R.; Birnbaum, M.J. Insulin is required to maintain albumin expression by inhibiting forkhead box O1 protein. J. Biol. Chem. 2016, 291, 2371–2378. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, P.M.; Bush, J.A.; Suryawan, A.; Nguyen, H.V.; Davis, T.A. Insulin and amino acids independently stimulate skeletal muscle protein synthesis in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E110–E119. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Suryawan, A.; Orellana, R.A.; Fiorotto, M.L.; Burrin, D.G. Amino acids and insulin are regulators of muscle protein synthesis in neonatal pigs. Animal 2010, 4, 1790–1796. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, J.L.; Martínez, M.; Trujillo, J.E.; Zamora, V.; Cordero, J.L.; Sánchez-Torres, M.T. Plasma urea nitrogen concentration and growth performance of finishing pigs fed sorghum-soybean meal, low-protein diets. J. Appl. Anim. Res. 2008, 33, 7–12. [Google Scholar] [CrossRef]
- Whang, K.Y.; Easter, R.A. Blood urea nitrogen as an index of feed efficiency and lean growth potential in growing-finishing swine. Asian-Australas. J. Anim. 2000, 13, 811–816. [Google Scholar] [CrossRef]
- Coma, J. Use of plasma urea nitrogen as a rapid response criterion to estimate the lysine requirements of growing and lactating pigs. J. Anim. Sci. 1995, 73, 472–481. [Google Scholar] [CrossRef]
- Suiryanrayna, M.V.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Partanen, K.H.; Mroz, Z. Organic acids for performance enhancement in pig diets. Nutr. Res. Rev. 1999, 12, 117–145. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, G.; Nguyen, H.T.T.; Yan, Y.; Charrier-Hisamuddin, L.; Sitaraman, S.V.; Merlin, D. Butyrate transcriptionally enhances peptide transporter PepT1 expression and activity. PLoS ONE 2008, 3, e2476. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Dong, L.; Xu, W.; Bai, K.; Lu, C.; Wu, Y.; Wang, T. Dietary tributyrin supplementation attenuates insulin resistance and abnormal lipid metabolism in suckling piglets with intrauterine growth retardation. PLoS ONE 2015, 10, e0136848. [Google Scholar] [CrossRef]
- Husveth, F.; Szegleti, C.; Neogrady, Z. Infusion of various short chain fatty acids causes different changes in the blood glucose and insulin concentrations in growing lambs deprived of food overnight. J. Vet. Med. Sci. 1996, 43, 437–444. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577. [Google Scholar] [CrossRef]
- Liu, J.L.; Segovia, I.; Yuan, X.L.; Gao, Z.H. Controversial roles of gut microbiota-derived short chain fatty acids (SCFAs) on pancreatic beta-cell growth and insulin secretion. Int. J. Mol. Sci. 2020, 21, 910. [Google Scholar] [CrossRef]
- Boukhliq, R.; Martin, G.B. Administration of fatty acids and gonadotrophin secretion in the mature ram. Anim. Reprod. Sci. 1997, 49, 143–159. [Google Scholar] [CrossRef]
- Weber, T.E.; Kerr, B.J. Effect of sodium butyrate on growth performance and response to lipopolysaccharide in weanling pigs. J. Anim. Sci. 2008, 86, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Hillmann, E.; Schrader, L.; Mayer, C.; Gygax, L. Effects of weight, temperature and behaviour on the circadian rhythm of salivary cortisol in growing pigs. Animal 2008, 2, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras Durand, F.; Dunière, L.; Blanquet-Diot, S.; Forano, E. Microbiota Composition and Functional Profiling Throughout the Gastrointestinal Tract of Commercial Weaning Piglets. Microorganisms 2019, 7, 343. [Google Scholar] [CrossRef]
- Schierack, P.; Walk, N.; Reiter, K.; Weyrauch, K.D.; Wieler, L.H. Composition of intestinal Enterobacteriaceae populations of healthy domestic pigs. Microbiology 2007, 153, 3830–3837. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhu, W.; Hang, S. Effects of long-term dietary protein restriction on intestinal morphology, digestive enzymes, gut hormones, and colonic microbiota in pigs. Animals 2019, 9, 180. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lei, Z.; Chen, L.; Qi, Z.; Wang, W.; Qiao, J. Lactobacillus acidophilus, alleviates the inflammatory response to enterotoxigenic Escherichia coli, k88 via inhibition of the nf-κb and p38 mitogen-activated protein kinase signaling pathways in piglets. BMC Microbiol. 2016, 16, 273. [Google Scholar] [CrossRef]
- Fouhse, J.M.; Zijlstra, R.T.; Willing, B.P. The role of gut microbiota in the health and disease of pigs. Anim. Front. 2016, 6, 30–36. [Google Scholar] [CrossRef]
- Heinritz, S.N.; Weiss, E.; Eklund, M.; Aumiller, T.; Louis, S.; Rings, A.; Messner, S.; Camarinha-Silva, A.; Seifert, J.; Bischoff, S.C.; et al. Intestinal microbiota and microbial metabolites are changed in a pig model fed a high-fat/low-fiber or a low-fat/high-fiber diet. PLoS ONE 2016, 11, e0154329. [Google Scholar] [CrossRef]
- Sun, J.; Du, L.; Li, X.; Zhong, H.; Ding, Y.; Liu, Z.; Ge, L. Identification of the core bacteria in rectums of diarrheic and non-diarrheic piglets. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Hu, Z.; Guo, Y. Effects of dietary sodium butyrate supplementation on the intestinal morphological structure, absorptive function and gut flora in chickens. Anim. Feed Sci. Technol. 2007, 132, 240–249. [Google Scholar] [CrossRef]
- Papon, M.; Talon, R. Factors affecting growth and lipase production by meat lactobacilli strains and Brochothrix thermosphacta. J. Appl. Bacteriol. 1988, 64, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut microbiota dysbiosis in postweaning piglets: Understanding the keys to health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Huang, X.; Wang, P.; Yan, Z.; Sun, W.; Zhao, S.; Gun, S. Longitudinal development of the gut microbiota in healthy and diarrheic piglets induced by age-related dietary changes. MicrobiologyOpen 2019, 8, e923. [Google Scholar] [CrossRef] [PubMed]
- McCormack, U.M.; Curião, T.; Buzoianu, S.G.; Prieto, M.L.; Ryan, T.; Varley, P.; O’Sullivan, O. Exploring a possible link between the intestinal microbiota and feed efficiency in pigs. Appl. Environ. Microbiol. 2017, 83, e00380. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Basal Diet |
|---|---|
| Ingredients | % as Fed Basis |
| Barley, meal | 25.15 |
| Wheat, meal | 19.41 |
| Corn, flakes | 14.03 |
| Corn, meal | 4.85 |
| Soybean, meal | 4.65 |
| Soy protein concentrates | 4.11 |
| Biscuits, meal | 4.00 |
| Dextrose monohydrate | 3.50 |
| Wheat middling | 4.32 |
| Whey protein concentrate | 3.00 |
| Fish, meal | 2.50 |
| Milk whey | 2.50 |
| Coconut oil | 1.00 |
| Soy oil | 1.00 |
| Plasma, meal | 1.00 |
| Organic Acids 1 | 1.00 |
| Dicalcium phosphate | 0.85 |
| Animal fats | 0.70 |
| L-Lysine | 0.50 |
| Benzoic acid | 0.40 |
| L-Threonine | 0.35 |
| DL-Methionine | 0.35 |
| Sodium Chloride | 0.27 |
| Vitamins 2 | 0.25 |
| L-Valine (96.5%) | 0.15 |
| L-Tryptophan | 0.08 |
| Flavouring 3 | 0.04 |
| Copper sulphate | 0.04 |
| Calculated Nutrient Levels 4 | |
| Crude protein | 16.92 |
| Ether extract | 5.06 |
| Crude fibre | 3.15 |
| Ash | 5.1 |
| DE 5 (Mc/Kg) | 3.43 |
| p-Value | DF (Num DF; Den DF) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Items 1 | CTRL | TRIB | SE | TRT | DAY | TRT × DAY | TRT | DAY | TRT × DAY |
| BW (kg) | 0.362 | <0.001 | <0.001 | 1; 10 | 3; 459 | 3; 459 | |||
| day 0 | 8.70 | 8.76 | 1.04 | ||||||
| day 14 | 11.10 | 11.84 | 1.04 | ||||||
| day 28 | 15.40 a | 17.10 b | 1.04 | ||||||
| day 40 | 20.10 a | 23.20 b | 1.04 | ||||||
| ADG (kg) | 0.034 | <0.001 | 0.125 | 1; 10 | 2; 20 | 2; 20 | |||
| day 0–day 14 | 0.171 | 0.220 | 0.03 | ||||||
| day 14–day 28 | 0.309 | 0.375 | 0.03 | ||||||
| day 28–day 40 | 0.395 a | 0.509 b | 0.03 | ||||||
| ADFI (kg) | 0.095 | <0.001 | 0.139 | 1; 10 | 4; 40 | 4; 40 | |||
| day 0–day 14 | 0.327 | 0.426 | 0.03 | ||||||
| day 14–day 21 | 0.596 | 0.601 | 0.03 | ||||||
| day 21–day 28 | 0.797 | 0.901 | 0.03 | ||||||
| G:F (%) | 0.049 | 0.189 | 0.160 | 1; 30 | 2; 30 | 2; 30 | |||
| Overall | 51.41 a | 56.53 b | 1.76 | ||||||
| day 0–day 14 | 52.44 | 50.75 | 3.06 | ||||||
| day 14–day 28 | 52.26 | 62.06 | 3.06 | ||||||
| day 28–day 40 | 49.54 | 56.79 | 3.06 | ||||||
| Items 1 | CTRL | TRIB | SE | p-Value | DF |
|---|---|---|---|---|---|
| Total protein content (g/L) | 52.87 | 56.82 | 1.78 | 0.1314 | 21 |
| Albumin (g/L) | 19.30 a | 24.06 b | 0.76 | 0.0002 | 21 |
| Globulin (g/L) | 33.58 | 32.77 | 1.78 | 0.7504 | 21 |
| A/G * | 0.58 a | 0.78 b | 0.05 | 0.0117 | 21 |
| Urea (mmol/L) | 2.18 a | 1.08 b | 0.23 | 0.0026 | 21 |
| ALT-GPT * (IU/L) | 38.33 | 32.91 | 1.87 | 0.0528 | 21 |
| AST-GOT * (IU/L) | 54.16 | 48.27 | 4.10 | 0.3218 | 21 |
| ALP * (UI/L) | 165.67 | 194.00 | 16.68 | 0.2432 | 21 |
| Total bilirubin (umol/l) | 1.98 | 1.79 | 0.11 | 0.2425 | 21 |
| Glucose (mmol/L) | 5.00 a | 5.83 b | 0.27 | 0.0396 | 21 |
| Total cholesterol (mmol/L) | 2.50 | 2.73 | 0.09 | 0.1008 | 21 |
| Calcium (mmol/L) | 2.28 | 2.44 | 0.07 | 0.1068 | 21 |
| Phosphorus (mmol/L) | 3.05 | 3.19 | 0.10 | 0.3253 | 21 |
| Magnesium (mmol/L) | 0.84 | 0.87 | 0.02 | 0.3726 | 21 |
| HDL (mmol/L) | 0.76 a | 0.88 b | 0.04 | 0.0349 | 21 |
| LDL (mmol/L) | 1.60 | 1.73 | 0.07 | 0.1961 | 21 |
| Creatinine (μmol/L) | 78.91 | 72.18 | 4.79 | 0.3318 | 21 |
| Triglycerides (mmol/L) | 0.68 | 0.63 | 0.05 | 0.5331 | 21 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotira, S.; Dell’Anno, M.; Caprarulo, V.; Hejna, M.; Pirrone, F.; Callegari, M.L.; Tucci, T.V.; Rossi, L. Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets. Animals 2020, 10, 726. https://doi.org/10.3390/ani10040726
Sotira S, Dell’Anno M, Caprarulo V, Hejna M, Pirrone F, Callegari ML, Tucci TV, Rossi L. Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets. Animals. 2020; 10(4):726. https://doi.org/10.3390/ani10040726
Chicago/Turabian StyleSotira, Stefania, Matteo Dell’Anno, Valentina Caprarulo, Monika Hejna, Federica Pirrone, Maria Luisa Callegari, Telma Vieira Tucci, and Luciana Rossi. 2020. "Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets" Animals 10, no. 4: 726. https://doi.org/10.3390/ani10040726
APA StyleSotira, S., Dell’Anno, M., Caprarulo, V., Hejna, M., Pirrone, F., Callegari, M. L., Tucci, T. V., & Rossi, L. (2020). Effects of Tributyrin Supplementation on Growth Performance, Insulin, Blood Metabolites and Gut Microbiota in Weaned Piglets. Animals, 10(4), 726. https://doi.org/10.3390/ani10040726