Impact of Thermal and High-Pressure Treatments on the Microbiological Quality and In Vitro Digestibility of Black Soldier Fly (Hermetia illucens) Larvae

 ,
, 
 ,
, 
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
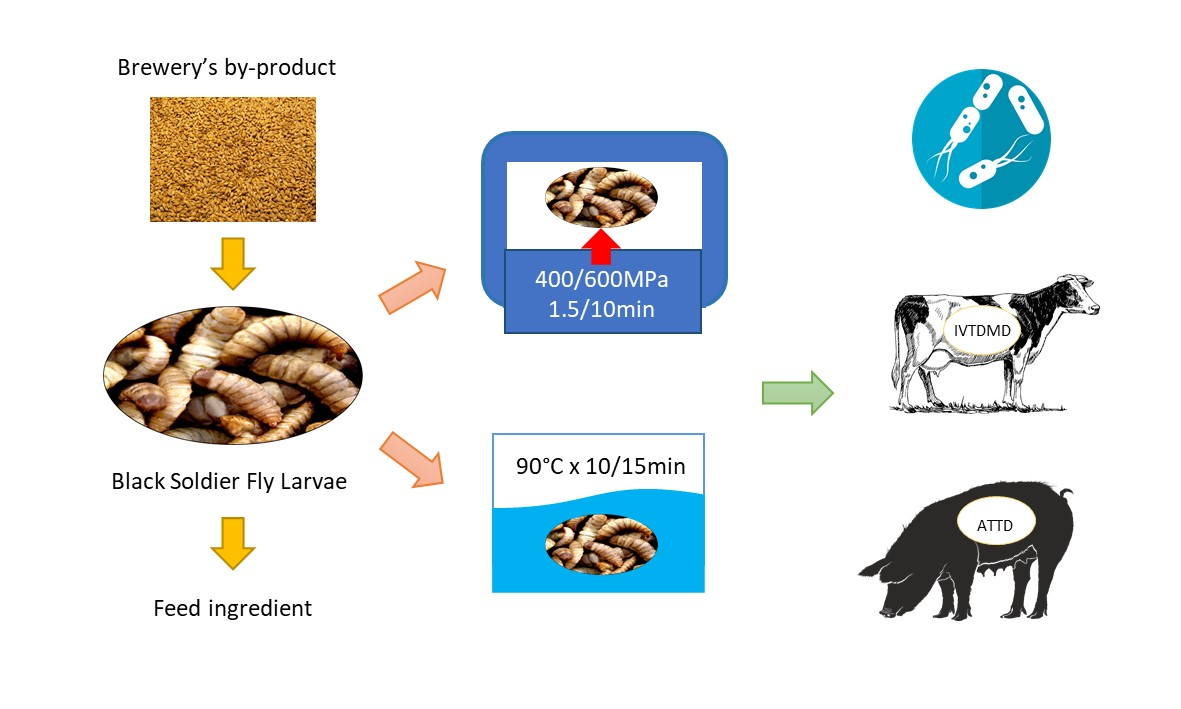
2.1. Preparation of Larvae
2.2. Chemical Composition
2.3. Thermal Processing (TP)
2.4. High-Pressure Processing (HPP)
2.5. Microbiological Analyses
2.6. In Vitro Dry Matter Digestibility
2.7. Statistical Analysis
3. Results
3.1. Nutritional Composition
3.2. Microbial Analysis
3.3. In Vitro Digestibility
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- European Commission Commission Regulation (EU) 2017/1151. Off. J. Eur. Union 2017, L175, 1–643.
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Pastor, B.; Velasquez, Y.; Gobbi, P.; Rojo, S. Conversion of organic wastes into fly larval biomass: Bottlenecks and challenges. J. Insects Food Feed 2015, 1, 179–193. [Google Scholar] [CrossRef]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals fed insect-based diets: State-of-the-art on digestibility, performance and product quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef]
- Poelaert, C.; Despret, X.; Sindic, M.; Beckers, Y.; Francis, F.; Portetelle, D.; Soyeurt, H.; Théwis, A.; Bindelle, J. Cooking has variable effects on the fermentability in the large intestine of the fraction of meats, grain legumes, and insects that is resistant to digestion in the small intestine in an in vitro model of the pig’s gastrointestinal tract. J. Agric. Food Chem. 2017, 65, 435–444. [Google Scholar] [CrossRef]
- Poelaert, C.; Francis, F.; Alabi, T.; Caparros Megido, R.; Crahay, B.; Bindelle, J.; Beckers, Y. Protein value of two insects, subjected to various heat treatments, using growing rats and the protein digestibility-corrected amino acid score. J. Insects Food Feed 2018, 4, 77–87. [Google Scholar] [CrossRef]
- Kinyuru, J.N.; Kenji, G.M.; Njoroge, S.M.; Ayieko, M. Effect of processing methods on the in vitro protein digestibility and vitamin content of edible winged termite (Macrotermes subhylanus) and grasshopper (Ruspolia differens). Food Bioprocess Technol. 2010, 3, 778–782. [Google Scholar] [CrossRef]
- Ekpo, K.E. Effect of processing on the protein quality of four popular insects consumed in Southern Nigeria. Arch. Appl. Sci. Res. 2011, 3, 307–326. [Google Scholar]
- Rumpold, B.A.; Schlüter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Fröhling, A.; Reineke, K.; Knorr, D.; Boguslawski, S.; Ehlbeck, J.; Schlüter, O. Comparison of volumetric and surface decontamination techniques for innovative processing of mealworm larvae (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2014, 26, 232–241. [Google Scholar] [CrossRef]
- Caparros Megido, R.; Poelaert, C.; Ernens, M.; Liotta, M.; Blecker, C.; Danthine, S.; Tyteca, E.; Haubruge, É.; Alabi, T.; Bindelle, J.; et al. Effect of household cooking techniques on the microbiological load and the nutritional quality of mealworms (Tenebrio molitor L. 1758). Food Res. Int. 2018, 106, 503–508. [Google Scholar] [CrossRef]
- Bolumar, T.; Middendorf, D.; Toepfl, S.; Heinz, V. Structural Changes in Foods Caused by High-Pressure Processing. In High Pressure Processing of Food; Food Engineering Series; Balasubramaniam, V., Barbosa-Cánovas, G., Lelieveld, H., Eds.; Springer: New York, NY, USA, 2016. [Google Scholar]
- AOAC. Official Methods of Analysis, Association of Analytical Chemists, 15th ed.; AOAC: Washington, DC, USA, 2000; pp. 141–144. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Robertson, J.B.; Van Soest, P.J. The detergent system of analysis and its application to human foods. In The Analysis of Dietary Fiber in Food; James, W.P.T., Theander, O., Eds.; Marcel Dekker: New York, NY, USA, 1981. [Google Scholar]
- European Commission Commission Regulation (EC) No 152/2009. Off. J. Eur. Union 2009, L54, 1–130.
- Ichihara, K.; Fukubayashi, Y. Preparation of fatty acid methyl esters for gas-liquid chromatography. J. Lipid Res. 2010, 51, 635–640. [Google Scholar] [CrossRef]
- Boisen, S.; Fernández, J.A. Prediction of the total tract digestibility of energy in feedstuffs and pig diets by in vitro analyses. Anim. Feed Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- ANKOM In Vitro True Digestibility using DAISY II Incubator. Available online: https://www.ankom.com/analytical-methods-support/daisy-incubators. (accessed on 23 February 2020).
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Gasco, L.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Shumo, M.; Osuga, I.M.; Khamis, F.M.; Tanga, C.M.; Fiab, K.K.M.; Subramanian, S.; Ekesi, S.; Van Huis, A.; Borgemeister, C.; Fiaboe, K.K.M.; et al. The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- Sauvant, D.; Perez, J.; Tran, G. Tables of Composition and Nutritional Value of Feed Materials; INRA Editions: Paris, France, 2004; ISBN 9789076998411. [Google Scholar]
- Bosch, G.; Vervoort, J.J.M.; Hendriks, W.H. In vitro digestibility and fermentability of selected insects for dog foods. Anim. Feed Sci. Technol. 2016, 221, 174–184. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Clercq, D.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens ) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2016, 97, 2594–2600. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagb, R.; Torstensen, B.E.; Lock, E. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- Spranghers, T.; Michiels, J.; Vrancx, J.; Ovyn, A.; Eeckhout, M.; De Clercq, P.; De Smet, S. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim. Feed Sci. Technol. 2018, 235, 33–42. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish Offal Recycling by the Black Soldier Fly Produces a Foodstuff High in Omega-3 Fatty Acids. J. World Aquac. Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Jayanegara, A.; Yantina, N.; Novandri, B.; Laconi, E.B.; Nahrowi, N.; Ridla, M. Evaluation of some insects as potential feed ingredients for ruminants: Chemical composition, in vitro rumen fermentation and methane emissions. J. Indones. Trop. Anim. Agric. 2017, 42, 247–254. [Google Scholar] [CrossRef]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J. Insects Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; Camenzuli, L.; Belluco, S.; Meijer, N.; Ricci, A.; Camenzuli, L.; Belluco, S.; Meijer, N.; Ricci, A. Food Safety Issues Related to Uses of Insects for Feeds and Foods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1172–1183. [Google Scholar] [CrossRef]
- European Commission Commission Directive 2002/32/EC. Off. J. Eur. Union 2002, L140, 1–22.
- Hahn, T.; Roth, A.; Febel, E.; Fijalkowska, M.; Schmitt, E.; Arsiwalla, T.; Zibek, S. New methods for high-accuracy insect chitin measurement. J. Sci. Food Agric. 2018, 98, 5069–5073. [Google Scholar] [CrossRef]
- Larouche, J.; Deschamps, M.H.; Saucier, L.; Lebeuf, Y.; Doyen, A.; Vandenberg, G.W. Effects of Killing Methods on Lipid Oxidation, Colour and Microbial Load of Black Soldier Fly (Hermetia illucens) Larvae. Animals 2019, 9, 182. [Google Scholar] [CrossRef]
- Kashiri, M.; Marin, C.; Garzo, R.; Rodrigo, D.; Martı, A.; Garzón, R.; Rosell, C.M.; Rodrigo, D.; Martínez, A. Use of high hydrostatic pressure to inactivate natural contaminating microorganisms and inoculated E. coli O157:H7 on Hermetia illucens larvae. PLoS ONE 2018, 13, e0194477. [Google Scholar] [CrossRef]
- Reineke, K.; Doehner, I.; Schlumbach, K.; Baier, D.; Mathys, A.; Knorr, D. The different pathways of spore germination and inactivation in dependence of pressure and temperature. Innov. Food Sci. Emerg. Technol. 2012, 13, 34–41. [Google Scholar] [CrossRef]
- Regmi, P.R.; Ferguson, N.S.; Zijlstra, R.T. In vitro digestibility techniques to predict apparent total tract energy digestibility of wheat in grower pigs. J. Anim. Sci. 2009, 87, 3620–3629. [Google Scholar] [CrossRef]
- Poelaert, C.; Beckers, Y.; Despret, X.; Portetelle, D.; Francis, F.; Bindelle, J. In vitro evaluation of fermentation characteristics of two types of insects as potential novel protein feeds for pigs. J. Anim. Sci. 2016, 94, 198–201. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| DM (g/100g) | CP | EE | Ash | aNDF | ADF | ADL |
| 34.1 ± 0.19 | 49.1 ± 0.51 | 36.6 ± 0.69 | 5.45 ± 0.21 | 13.0 ± 1.04 | 7.92 ± 0.48 | 1.45 ± 0.12 |
| Ca | K | Mg | P | Fe | Mn | Zn |
| 18.5 ± 0.38 | 6.91 ± 0.19 | 2.87 ± 0.11 | 4.40 ± 0.19 | 0.15 ± 0.04 | 0.12 ± 0.03 | 0.11 ± 0.04 |
| Alanine | 2.44 | Lysine | 2.60 |
| Arginine | 2.15 | Methionine | 0.84 |
| Aspartic acid | 3.95 | Phenylalanine | 1.78 |
| Cystine | 0.36 | Proline | 2.08 |
| Glutamic acid | 4.75 | Serine | 1.66 |
| Glycine | 2.25 | Threonine | 1.62 |
| Histidine | 1.33 | Tryptophan | 0.60 |
| Isoleucine | 1.75 | Tyrosine | 2.26 |
| Leucine | 2.78 | Valine | 2.41 |
| C10:0 | 0.92 | C16:1 | 2.29 | SFA | 64.5 |
| C12:0 | 36.0 | C18:0 | 2.28 | MUFA | 12.3 |
| C14:0 | 7.83 | C18:1 | 9.38 | PUFA | 19.6 |
| C14:1 | 0.15 | C18:2n6 | 18.1 | ||
| C15:0 | 0.21 | C18:3n3 | 1.48 | ||
| C15:1 | 0.10 | C20:0 | 0.11 | ||
| C16:0 | 16.8 | C20:1 | 0.19 |
| Treatment | Control | TP | HPP | SEM | p | ||||
|---|---|---|---|---|---|---|---|---|---|
| Pressure (MPa) | 400 | 600 | |||||||
| Time (min) | 10 | 15 | 1.5 | 10 | 1.5 | 10 | |||
| TVC | 7.97 a | 5.52 e | 5.63 e | 7.65 a,b | 6.53 d | 7.28 b,c | 6.67 c,d | 0.201 | <0.001 |
| ENB | 7.65 a | <2 d | <2 d | 6.12 b | 3.32 c | 2.09 d | <2 d | 0.485 | <0.001 |
| LAB | 6.50 a | <2 c | <2 c | 6.60 a | 4.73 b | <2 c | <2 c | 0.476 | <0.001 |
| YM | 5.07 a | <2 c | <2 c | 3.36 b | <2 c | <2 c | <2 c | 0.265 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, M.; Ortuño, J.; Stratakos, A.C.; Linton, M.; Corcionivoschi, N.; Elliott, T.; Koidis, A.; Theodoridou, K. Impact of Thermal and High-Pressure Treatments on the Microbiological Quality and In Vitro Digestibility of Black Soldier Fly (Hermetia illucens) Larvae. Animals 2020, 10, 682. https://doi.org/10.3390/ani10040682
Campbell M, Ortuño J, Stratakos AC, Linton M, Corcionivoschi N, Elliott T, Koidis A, Theodoridou K. Impact of Thermal and High-Pressure Treatments on the Microbiological Quality and In Vitro Digestibility of Black Soldier Fly (Hermetia illucens) Larvae. Animals. 2020; 10(4):682. https://doi.org/10.3390/ani10040682
Chicago/Turabian StyleCampbell, Mairead, Jordi Ortuño, Alexandros Ch. Stratakos, Mark Linton, Nicolae Corcionivoschi, Tara Elliott, Anastasios Koidis, and Katerina Theodoridou. 2020. "Impact of Thermal and High-Pressure Treatments on the Microbiological Quality and In Vitro Digestibility of Black Soldier Fly (Hermetia illucens) Larvae" Animals 10, no. 4: 682. https://doi.org/10.3390/ani10040682
APA StyleCampbell, M., Ortuño, J., Stratakos, A. C., Linton, M., Corcionivoschi, N., Elliott, T., Koidis, A., & Theodoridou, K. (2020). Impact of Thermal and High-Pressure Treatments on the Microbiological Quality and In Vitro Digestibility of Black Soldier Fly (Hermetia illucens) Larvae. Animals, 10(4), 682. https://doi.org/10.3390/ani10040682