1. Introduction
Recently, much attention has been paid to broiler meat due to its characteristics of low fat, high protein, and richness of functional peptides [
1], which are all favored by health-conscious consumers. The nutritional supply is a vital element affecting the quality and composition of poultry meat [
2]. As modern broiler chickens grow faster and the time-to-market becomes shorter, the embryonic development in the hatching phase becomes more and more important [
3]. Therefore, the nutritional supply to broiler embryos may be a beneficial pathway to enhance the quality and composition of poultry meat.
In ovo feeding (IOF) is an emerging technology to administer exogenous nutrients into the hatching eggs for better embryonic development and postnatal growth exhibition. Preferably, the amnion of the late-term embryo (17.5–19 d of incubation) is the ideal site for nutrients administration to enhance growth performance and energy reserves [
4,
5,
6]. In this context, in order to obtain favorable broiler meat quality, the modulation of the embryo nutritional supply in the incubation stage is a promising solution. Indeed, evidence has shown that administration of equol, L-ascorbic acid, or synbiotics in the incubation phase may effectively improve broiler meat quality [
3,
7,
8,
9]. Since the risk for protein catabolism in late embryos exists [
10], it is interesting to probe the efficacy of IOF of amino acids on meat quality of market size broilers.
Arginine (Arg), one of the nutritionally essential amino acids in birds, had been used in the poultry diet to make meat quality better and increase antioxidant defense under normal circumstances [
11]. N-carbamylglutamate (NCG), a stable activator for endogenous Arg synthesis, has been demonstrated to enhance Arg concentration, stimulate protein synthesis in skeletal muscle, [
12] as well as improve meat quality in pigs [
13] and sheep [
14]. Up to now, the application of NCG in broilers is scarce. The only available study regarding NCG in broilers showed that diet supplementation of NCG in Chinese yellow-feathered broilers positively affected growth performance, tissue development, and blood parameters [
15]. Whether the exposure of NCG in the incubation phase will exert positive effects on growth and meat quality in broilers is still unknown. Based on the above literature, exploring the influence of IOF of NCG on the meat quality of broilers could be valuable. In addition, it was reported that some nutrients or bioactive substances exert the same functions irrespective of delivery route, as evidenced by the reduction of susceptibility of poultry to pulmonary hypertension syndrome with supplemental Arg performed in ovo or in the feed [
16], and the positive effects on meat quality of broilers affected by prebiotics either by an in ovo or in water route [
17]. However, IOF uses smaller amounts of supplements than diet supplementation, and the comprehensive effects of IOF technology are superior to diet addition for broilers in some supplements [
16,
18]. Thus, it is reasonable to hypothesize that in ovo delivery of NCG can convey profitable influences on broiler meat quality. The objectives of this research were to investigate the influences of amniotic injection of NCG on meat quality of broiler cocks as well as to demonstrate whether exposure to NCG modulated the glycolytic and free Arg content, nutritional properties, and antioxidant capacity in the pectoral muscle.
2. Materials and Methods
2.1. Incubation
Fifty fertilized broiler eggs with the mean weight of 58–62 g from Ross 308 breeder were randomly assigned to every replicate. A total of 18 replicates were randomly placed in a microcomputer automatic incubator (37.8 ± 0.1 °C, 60%). After injection (at 17.5 d of incubation), hatching eggs were diverted to the hatching basket, which was placed in a microcomputer automatic hatcher (37.2 ± 0.1 °C, 70%) until chicks were hatched.
2.2. Treatment Solutions and Injection Procedure
NCG of 2 g, which was accurately weighed, was dissolved in 100 mL aseptic saline (0.85%) to prepare the NCG solution. All solutions were passed through a syringe filter membrane for sterilization, and then were drawn into 1 mL syringes pending for injection.
After 17.5 d of incubation, the fertile eggs were candled and the sites of injection were marked. Subsequently, 0.1 mL saline or NCG solution were injected into the amniotic cavity of eggs by a 2.54 cm, 22-gauge needle possessing a short beveled tip from the large end of the egg. To make sure the needle thrust entirely into the amnion, the whole needle was inserted into the large end of the egg. The injection site remained open. Based on our previous study, the accuracy of amnion injection is more than 95%. The routine procedure of checking the success of amnion injection was done by coomassie brilliant blue dye injection and autopsy observation.
2.3. Bird Housing and Feeding
Upon hatching, 72 healthy male chicks of each treatment were randomly assigned to six replicates for a 42-day grow-out feeding. A corn–soybean meal diet for starter (1–21 d) or grower (22–42 d) was formulated based on the National Research Council (1994). The starter diet contained 22% crude protein, 2950 kcal/kg metabolizable energy (ME), 1.15% digestible lysine, 0.85% digestible methionine + cystine, and 0.75% digestible threonine. The grower diet contained 20% crude protein, 3050 kcal/kg ME, 1.05% digestible lysine, 0.79% digestible methionine + cystine, and 0.69% digestible threonine. Chicks had free access to feed and water. Chicks were raised in wire floor cages (cage size, 110 × 100 × 55 cm
3) in a three-level battery under environmentally controlled room conditions in the Nankou CAAS experimental base (Beijing, China). All the animal management was in accordance with the Ross 308 broiler management guide (Aviagen 2015) [
19]. After the experiment, the remaining broilers were quarantined and sent to a local slaughterhouse. All the experimental protocols (AEC-CAAS- 20190105) for this study were approved by the Animal Care and Use Committee of the Feed Research Institute of the Chinese Academy of Agricultural Sciences.
2.4. Sample Collection and Breast Weight Measurement
One healthy bird with the body weight (BW) similar to the mean value on day 42 of cage life was selected and sacrificed by jugular cutting for blood and muscle sample collection. After being plucked and eviscerated, the pectoral muscles were weighed. The left pectoral, which was snapped frozen using liquid nitrogen, was kept at −80 °C pending measurement of various physical parameters.
2.5. Hatchability and Growth Performance Measurement
On the day of hatching, the number of chicks in each replicate were counted to calculate the hatchability. Hatchability equals the ratio between the number of alive embryos at E 17 (17 d of incubation) and the number of chicks.
At the end of experiment, all broilers in each replicate were weighed and feed intake was recorded. The growth performance was expressed by average daily gain (ADG), average daily feed intake (ADFI), and F/G (ADFI/ADG).
2.6. Physical Parameters of Pectoral Muscle
The method of determining pH values was improved based on the foundation of Qi et al. (2018) [
20]. Three different locations (cranial, medial, and caudal) within the medial surface (bone side) of the breast muscle were used for pH readings by a pH meter (CyberScan pH 310, Eutech Instruments Pte. Ltd., Singapore) at 45 min and 24 h postmortem. The ΔpH was calculated as pH 24 h − pH 45 min.
The method of determining the meat color was improved based on the foundation of Zhang et al. (2009) [
21]. Meat color was measured three times using a Chroma Meter (Chroma Meter WSC-S, Shanghai Precision and Scientific Instrument Co., Shanghai, China). The parameters (L*, a*, b*) of color were shown in the CIELab system.
The method of determining dripping loss was improved based on the foundation of Zhang et al. (2009) [
21]. Approximately 30 g of pectoralis major muscle (W1) was suspended in a sealed plastic case with precooling at 4 °C. After 24 h in a 4 °C refrigerator, filter paper was used to absorb the surface moisture of samples and they were weighed again (W2). Drip loss (%) = (W1− W2)/W1 × 100%.
The method of determining cooking loss was improved based on the foundation of Qi et al. (2018) [
20]. Meat samples in zip-sealed polyethylene bags were boiled in a water bath to 80 °C (i.e., until the sample’s center was 80 °C). Then, the samples were brought down to ambient temperature by running them under water. The samples were then dried using paper towel and weighed again (W3). Cooking loss (%) = (W2 − W3)/W2 × 100%.
2.7. Glycolytic Metabolite Measurements
Measurements of muscle glycolytic potential (GP) used the method recorded by Zhang et al. [
21]. Specifically, the sample (frozen muscle, 0.5 g) was placed in a 4.5 mL perchloric acid solution and homogenized for 3 min. To remove protein and other impurities, the homogenate was centrifuged (4 °C, 3000×
g, 10 min). To measure lactate in the supernatant, we used spectrophotometry, according to the instructions of the lactic acid determination kit (Nanjing Jiancheng Bioengineering Institute). In acetate buffer (pH 4.8, 55 °C, 2 h), glycogen was enzymatically hydrolyzed to glucose by amyloglucosidase (A7420, Sigma-Aldrich Inc., St. Louis, MO). Then, for measurement of glucose, we used a commercial glucose oxidase kit (Shanghai Rong Sheng Biotech Co. Ltd., Shanghai, China). GP = 2 × [glycogen] + [lactate].
2.8. Determination of the Antioxidant Index in Muscle
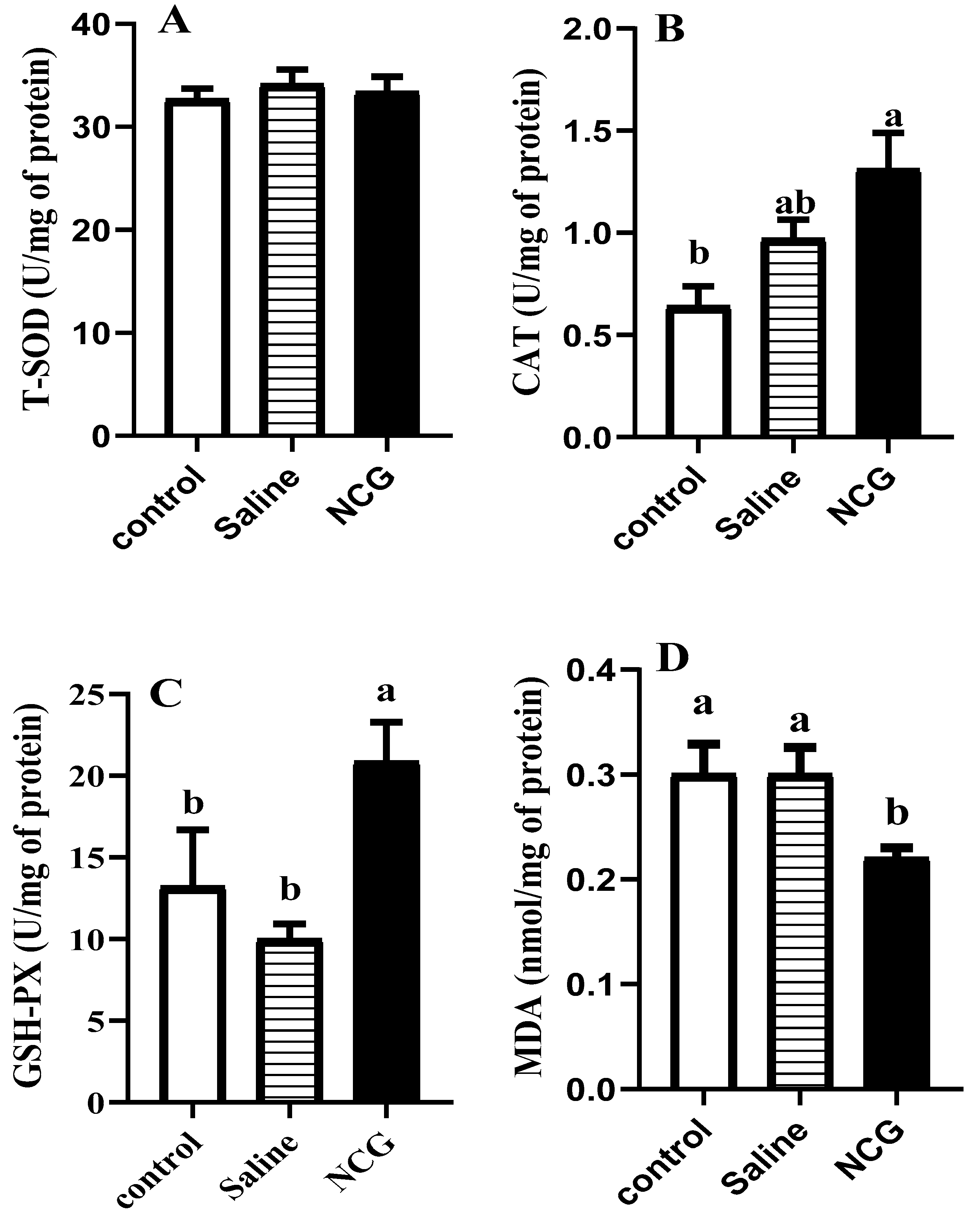
T sample (frozen muscle, 0.5 g) was put in 4.5 mL of a 0.8% saline solution and homogenized for 3 min. After centrifugation (4 °C, 3000× g, 10 min), supernatant fluid was collected for measurement of the activities of total superoxide dismutase (T-SOD), catalase (CAT), and glutathione peroxidase (GSH-Px) and of the volume of malondialdehyde (MDA). All the indicators were determined following by the manual provided with the kit (Nanjing Jiancheng Bioengineering Institute).
2.9. Hydrolytic Amino Acids Analysis
The method of determining hydrolyzed amino acids was improved based on the foundation of Nasr et al. [
22] Briefly, each fat-free sample (0.5 g) was placed in a nitrogen-filled sealed tube and was hydrolyzed with 10 mL HCl (6 M) in an oven (110 °C, 24 h). The hydrolysate was adjusted to a volume of 500 mL with chromatographic water before passing through a 0.2 μm filter membrane. The concentrations of amino acids labeled with phthalaldehyde were determined at 338 nanometer derivative wavelength using high performance liquid chromatography (HPLC) with Shimadzu SPD-20A. Mobile phase A is an organic phase (acetonitrile/methanol/chromatographic water = 4.5:4.5:1) and mobile phase B is an inorganic phase (4.5 g disodium hydrogen phosphate dodecahydrate, 4.75 g sodium tetraborate decahydrate, 1000 mL chromatographic water). The flow rates of the mobile phases are shown in
Table 1.
2.10. Free Amino Acid Analysis
The method of determining free amino acids was improved based on the foundation of Hartman et al. [
23] Briefly, the sample (frozen muscle, 0.5 g) was placed into a saline solution (ice-cold, 4.5 mL, 0.8%) to homogenize. After centrifugation (4 °C, 9000×
g, 10 min), 300 μL supernatant was mixed with 300 μL 1.5 M perchloric acid, left to stand for 10 min, then mixed with 150 μL of a 2 M potassium carbonate solution and left to stand for 20 min. The mixture was centrifuged and the supernatant filtered through a 0.2 μm membrane. Free amino acids were then determined by HPLC.
2.11. Chemical Composition of Pectoral Muscle
Muscle samples were put in a Petri dish and frozen and dried in a thermo freeze dryer (100 mbar pressure, −50 °C, Pilot7–12L, BIOCOOL) for 3 days. The Soxhlet extraction method was used to measure crude fat. Crude protein was measured using a Foss Kjeltec TM 8400 nitrogen/protein analyzer.
2.12. Statistical Analysis
All data analyses were implemented using SAS (version 9.4; SAS Institute, 2013). Means were separated for significance by Duncan’s multiple range tests. The data for meat quality, amino acids, antioxidants, glycolytic potential, and chemical parameters were analyzed using one-way ANOVA in SAS. Phenotypic correlations were performed using PROC CORR in SAS. The discrepancy was deemed statistically significant at p ≤ 0.05, and results are shown as means and SEM.
4. Discussion
N-carbamylglutamate (NCG) is a metabolic activator that stimulates the endogenous synthesis of Arg. NCG participates in activation of pyrrol 5 carboxylate synthase (P5CS) and carbamyl phosphate synthase 1 (CPS-I), then promotes the synthesis of citrulline, thus stimulating endogenous synthesis of Arg by argininosuccinate synthase (ASS) and argininosuccinate lyase (ASL) [
24]. The current study demonstrated that early exposure of NCG in the hatching phase may increase the Arg content in muscle of market size broilers. The increase of Arg had been revealed in the muscle of finishing pigs fed NCG-containing diets [
13]. Because Arg is involved in the metabolism of glucose and amino acids [
25], protein synthesis [
26], and antioxidant capacity [
27], it is reasonable to predict that the increased Arg induced by NCG exposure may affect glycolysis and antioxidant ability in broiler chickens.
It is through the process of energy depletion that muscle turns to meat. In early postmortem, ATP is supplied by the decomposition of phosphocreatine (PCr) and then by the decomposition of glycogen to lactate after the 70% of the PCr is used up [
28]. Glycolysis in muscle is a vital action in the postmortem period, when the oxygen supply for oxidative metabolism is lost due to exsanguination [
29]. The end product of glycolysis is lactic acid, whose content could serve as an indicator to reflect the process of glycolysis. In the current study, the reduced lactic acid content may be due to the delayed occurrence of glycolysis in the presence of the stimulated production of creatine from the increased Arg, which has been observed in early studies [
30,
31]. Lower lactic acid would contribute to the improvement of meat quality.
Meat quality could be greatly affected by the postmortem redox state [
32]. Metabolic processes in body and muscular tissue cause the formation of different reactive oxygen species (ROS), peroxides, singlet oxygen, and free radicals [
33,
34]. Excessive amounts of ROS or free radicals in muscles cause great damage to cell walls, functional proteins, and lipids, thus leading to the reduction of meat quality [
35]. In organisms, the antioxidative defense system, including enzymatic and non-enzymatic antioxidants, can prevent the production of ROS [
36]. Enzymatic antioxidant systems of superoxide dismutases (SOD), glutathione peroxidases (GSH-PX), and catalase (CAT), effectively decrease the accumulation and production of oxides [
37], further reducing the damage to cells and tissues. The dismutation of superoxide radicals (O
2−) is promoted by SOD to form oxygen and hydrogen peroxide (H
2O
2), then, H
2O
2 is reduced to glutathione and water by GSH-PX, and to oxygen and water by CAT [
38]. In our study, the increasing activity of CAT and GSH-PX indicated that IOF of NCG would benefit the enzymatic antioxidant system. Malondialdehyde (MDA), a biomarker for the evaluation of lipid peroxidation, usually increases in muscle with elevated oxidative production [
39]. The relatively lower concentration of MDA means that the muscle cells were more intact and the oxidation rate of lipids was slower in pectoral muscle of NCG-treated birds.
A major finding in our study was that injection of NCG in amnion markedly enhanced the protein content in pectoral muscle. The elevation of protein synthesis was also revealed in a previous study in piglets, in which NCG supplementation in diet improved the absolute rates of protein synthesis in skeletal muscle by increasing the level of Arg [
12]. Enhancement of protein synthesis in skeletal muscles has been shown to be stimulated by Arg through the mTOR signaling pathway [
40]. In addition, Arg supplementation increased fat content through the increase of PPARγ expression and stimulation of differentiation and proliferation in bovine adipocytes [
41]. In the current study, crude fat in pectoral muscle increased in the NCG in ovo supplementation group. The increase of protein and fat in pectoral muscle may be partly associated with the improvement of oxidation resistance. Previous research noted that free radicals of oxidized products could induce the increase of protein denaturation [
42] and lipid superoxidation in muscle [
43]. Our results showed that the increased crude fat content was closely correlated with the MDA content and protein content was closely related to GSH-PX, in pectoral muscle.
Water holding capacity (WHC) is a vital indicator of meat characteristics that affect processing and sensory qualities, and lower WHC is always linked with economic losses [
44]. Dripping loss is an easy and important parameter to reflect the WHC. Lactic acid and reactive oxygen generated from the biochemical reactions in the course of muscle turning to meat may decrease the completeness of muscle cell membranes and break down the functional proteins in muscle [
36], which would result in the increase of drip loss of muscle [
45]. In the current study, the reduction of drip loss was coincident with the decrease of MDA content in the pectoral muscle. Similar to our results, simultaneous decline of drip loss and MDA content was observed in porcine longissimus dorsi [
46]. These observations suggested that the reduction of oxidation products contributes to the reduction of drip loss in muscle. A previous study reported that decreases of glycolytic rate and lactic acid content may lead to enhancement of pH and decreased drip loss in pork [
47]. However, our results did not show the tie between lactic acid content, pH value, and drip loss. The discrepancy was probably due to the large variations during the course of pH decline in practical situations [
28]. It was suggested that pH values were closely connected with glycogen rather than the lactate concentration [
48]. In the current study, the decreased drip loss may be mainly related to the enhancement of antioxidative status. The antioxidant vitamin E has been shown to maintain muscle cell membranes [
49]. However, more investigation is needed to support this idea.
Meat flavor is also a vital index for meat quality for consumers, which is expressed by odor and taste. Free amino acids in muscle are important flavor-presenting substances and flavor precursors of chicken meat. The main substance of umami is Glu, and Ala, Ser, Thr, and Lys are the main substances of sweetness [
50]. Among the free amino acids, Arg and Ala content account for a higher proportion and are key flavor amino acids. The Maillard process, which occurs between amino acids and reducing sugars, is the major course for producing meat-fragrant compounds [
51]. The current research showed that IOF of NCG enhanced the content of free Arg and Ala, and the total delicious amino acids in pectoral muscle of broilers. The reason for this result may be that the NCG stimulated synthesis of Arg [
24] and then the Arg increased content of Ala [
52,
53]. The results showed that amniotic injection of NCG could positively affect the flavor of pectoral muscle in broilers.




{kind=link}