Hycole Doe Milk Properties and Kit Growth



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. The Quality of Rabbit Milk
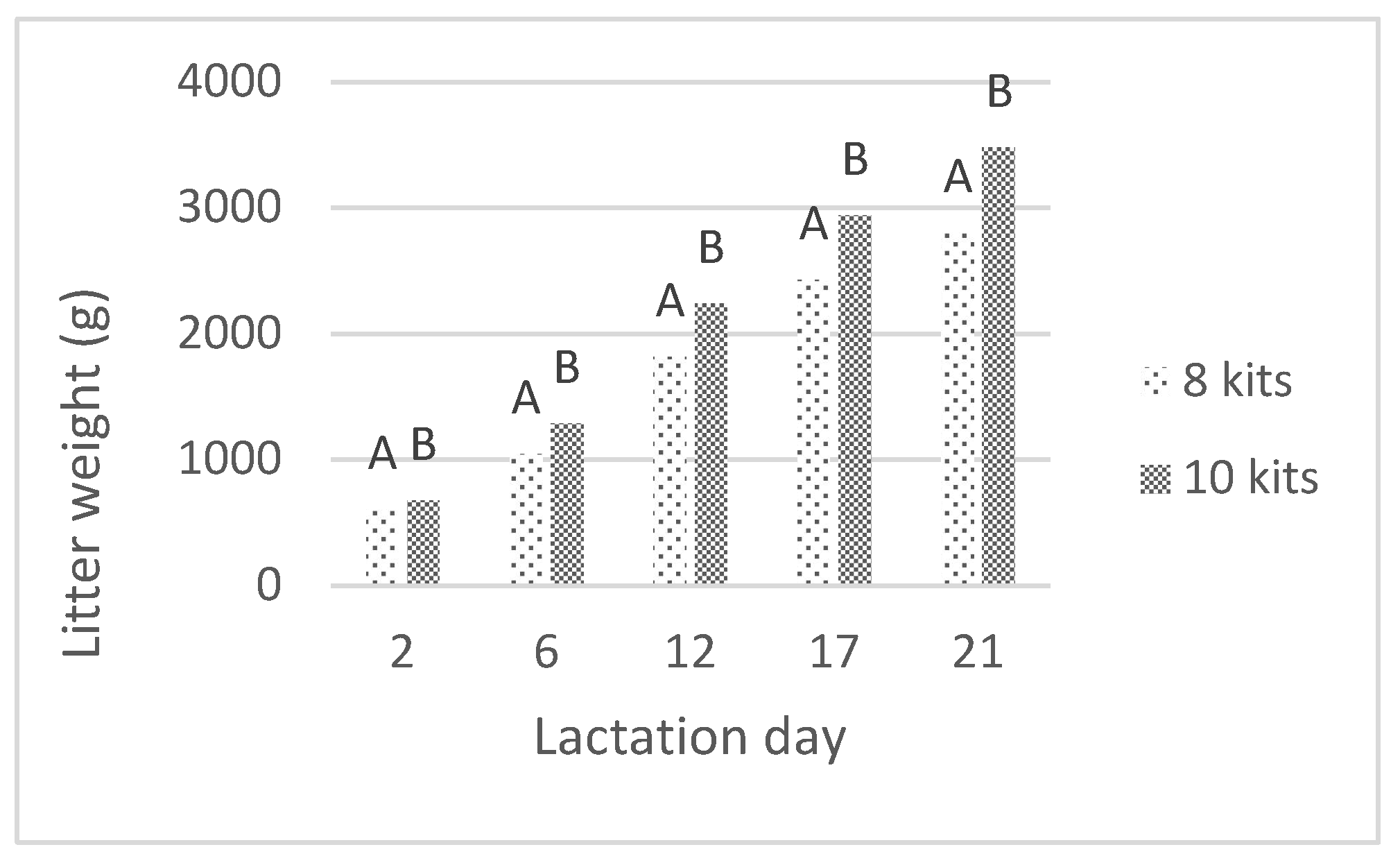
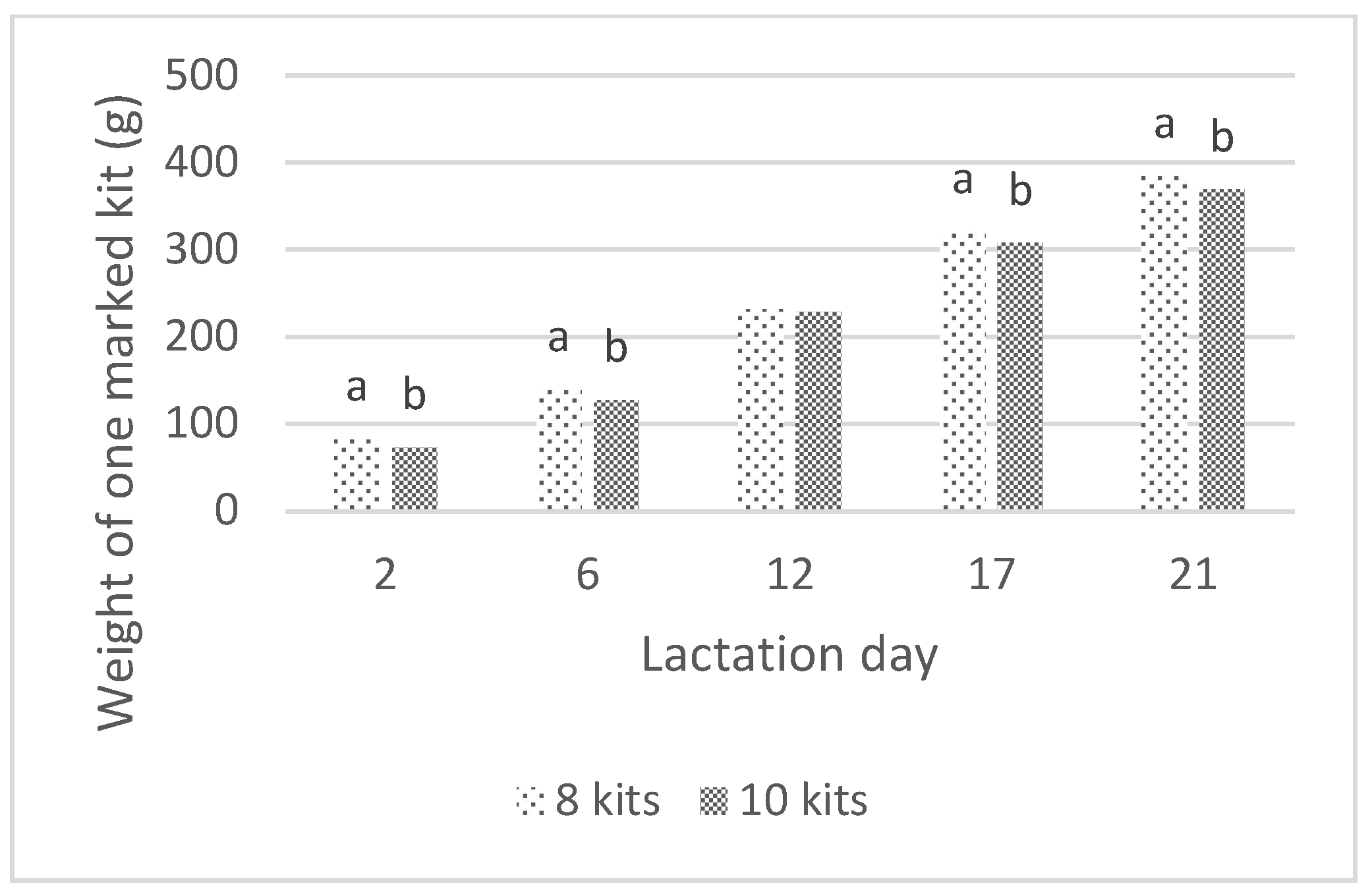
3.2. Daily Milk Production and Assocciated Traits
3.3. Phenotypic Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hudson, R.; Schaal, B.; Martínez-Gómez, M.; Distel, H. Mother-young relations in the European rabbit: Physiological and behavioural locks and keys. World Rabbit Sci. 2000, 8, 85–90. [Google Scholar] [CrossRef]
- Maertens, L.; Lebas, F.; Szendrö, Zs. Rabbit milk: A review of quantity, quality and non-dietary affecting factors. World Rabbit Sci. 2006, 14, 205–230. [Google Scholar] [CrossRef]
- Pałka, S.; Kmiecik, M.; Migdał, Ł.; Kozioł, K.; Otwinowska-Mindur, A.; Bieniek, J. Effect of housing system and breed on growth, slaughter traits and meat quality traits in rabbits (in Polish with English abstract). Roczniki Naukowe PTZ 2018, 14, 9–18. [Google Scholar]
- Fayeye, T.R.; Ayorinde, K.L. Effect of weaning litter size and sex on post weaning body weight, mortality and carcass characteristics of domestic rabbit in the humid tropics. In Proceedings of the 9th World Rabbit Congress, Verona, Italy, 10–13 June 2008; World Rabbit Science: Valencia, Spain, 2008; p. 1535. [Google Scholar]
- Bieniek, J.; Maj, D.; Derewicka, O.; Bonczar, Z. Slaughter traits of Burgundy Fawn rabbits and their crosses with New Zealand White (in Polish with English summary). Zywn-Nauk. Technol. Ja. 2012, 1, 154–163. [Google Scholar]
- Fortun-Lamothe, L.; Sabater, F. Estimation de la production laitière à partir de la croissance des lapereaux. In Proceedings of the 10èmes Journ. Rech. Cunicole, Paris, France, 19–20 November 2003; ITAVI éd. Paris: Paris, France, 2003; pp. 69–72. (In French). [Google Scholar]
- Xiccato, G.; Trocino, A.; Sartori, A.; Queaque, P.I. Effect of parity order and litter weaning age on the performance and body energy balance of rabbit does. Livest. Prod. Sci. 2004, 85, 239–251. [Google Scholar] [CrossRef]
- Xiccato, G.; Trocino, A.; Boiti, C.; Brecchia, G. Reproductive rhythm and litter weaning age as they affect rabbit doe performance and body energy balance. Anim. Sci. 2005, 81, 289–296. [Google Scholar] [CrossRef]
- Peaker, M.; Taylor, J.C. Milk secretion in the rabbit: Changes during lactation and the mechanism of ion transport. J. Physiol. 1975, 253, 527–545. [Google Scholar] [CrossRef]
- Lebas, F. Effet de la simultanéité de la lactation et de la gestation sur les performances laitières chez la lapine. Ann. Zootech. 1972, 21, 129–131. (In French) [Google Scholar] [CrossRef]
- Xiccato, G.; Parigi-Bini, R.; Dalle Zotte, A.; Carazzolo, A.; Cossu, M.E. Effect of dietary energy level, addition of fat and physiological state on performance and energy balance of lactating and pregnant rabbit does. Anim. Sci. 1995, 61, 387–398. [Google Scholar] [CrossRef]
- Coates, M.E.; Gregory, M.E.; Thompson, S.Y. The composition of rabbit’s meat. Brit. J. Nutr. 1964, 18, 583. [Google Scholar] [CrossRef]
- Lebas, F. Mesure quantitative de la production laitière chez la lapine. Ann. Zootech. 1968, 17, 169–182. [Google Scholar] [CrossRef]
- Lebas, F. Alimentation lactée et croissance pondérée du lapin avant sevrage. Ann. Zootech. 1969, 18, 197–208. [Google Scholar] [CrossRef]
- Lebas, F. Description d’une machine à traire les lapines. Ann. Zootech. 1970, 19, 223–228. [Google Scholar] [CrossRef]
- Lebas, F. Composition chimique du lait de lapine, evolution au cours de la traite et en fonction du stade de la lactation. Ann. Zootech. 1971, 20, 185–191. [Google Scholar] [CrossRef]
- Goode, J.A.; Taylor, J.C. Proceedings: A simple method of milking small animals. J. Physiol. 1974, 242, 5–7. [Google Scholar]
- Pascual, J.J.; Motta, W.; Cervera, C.; Quevedo, F.; Blas, E.; Fernández-Carmona, J. Effect of dietary energy source on the performance and perirenal fat thickness evolution of primiparous rabbit does. Anim. Sci. 2002, 75, 267–279. [Google Scholar] [CrossRef]
- Chrenek, P.; Chrastinova, L.; Kirchenrova, K.; Makarevich, V.; Foltys, V. The yield and composition of milk from transgenic rabbits. Asian-Aust. J. Anim. Sci. 2007, 4, 482–486. [Google Scholar] [CrossRef]
- Alhussien, M.N.; Dang, A.K. Milk somatic cells, factors influencing their release, future prospects, and practical utility in dairy animals: An overview. Vet. World 2018, 11, 562–577. [Google Scholar] [CrossRef]
- Qayyum, A.; Khan, J.A.; Hussain, R.; Avais, M.; Ahmad, N.; Khan, S. Investigation of milk and blood serum biochemical profile as an indicator of sub-clinical mastitis in Cholistani cattle. Pak. Vet. J. 2016, 36, 275–279. [Google Scholar]
- Kandeel, S.A.; Megahed, A.A.; Ebeid, M.H.; Constable, P.D. Ability of milk pH to predict subclinical mastitis and intramammary infection in quarters from lactating dairy cattle. J. Diary Sci. 2019, 102, 1417–1427. [Google Scholar] [CrossRef]
- Priolo, A.; Lanza, M.; Barbagallo, D.; Finocchiaro, L.; Biondi, L. Can the reflectance spectrum be used to trace grass feeding in ewe milk? Small Rumin. Res. 2003, 48, 103–107. [Google Scholar] [CrossRef]
- Morris, C.A.; Knight, T.W.; Newman, S.-A.N.; Hickey, S.M.; Death, A.F.; O’Neill, K.T.; Ridland, M. Genetic studies of carotenoid concentration in the plasma and milk of New Zealand dairy cattle. N. Z. J. Agric. Res. 2002, 45, 27–33. [Google Scholar] [CrossRef]
- Raty, J.A.; Peiponen, K.E. Reflectance study of milk in the UV-visible range. Appl. Spectrosc. 1999, 53, 1123–1127. [Google Scholar] [CrossRef]
- Solah, V.A.; Staines, V.; Honda, S.; Limley, H.A. Measurement of milk color and composition: Effect of dietary intervention on Western Australian Holstein-Friesian cow’s milk quality. J. Food Sci. 2007, 72, S560–S566. [Google Scholar] [CrossRef]
- Alcântara, L.A.P.; Fontan, R.C.I.; Bonomo, R.C.F.; Souza, E.C., Jr.; Sampaio, V.S.; Pereira, R.G. Density and dynamic viscosity of bovine milk affected by temperature and composition. Int. J. Food Eng. 2012, 8. [Google Scholar] [CrossRef]
- Rice, E.F.; Markley, A.L. The relation of natural acidity in milk to composition and physical properties. J. Diary Sci. 1924, 7, 468–483. [Google Scholar] [CrossRef]
- Niedźwiadek, S. Zasady hodowli królików (The principles of breeding rabbit, in Polish); PWRiL: Warszawa, Polska, 1981; pp. 21–25. [Google Scholar]
- CIE Central Bureau. CIE Recommendations on Uniform Color Spaces—Color Difference Equations, Psychometric Color Terms, CIE Technical Report, Supplement No. 2; CIE Publication: Vienna, Austria, 1978.
- SAS/STAT 9.3. User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2012. [Google Scholar]
- Vacca, G.M.; Stocco, G.; Dettori, M.L.; Pira, E.; Bittante, G. Milk yield, quality and coagulation properties of 6 breeds of goats: Environmental and individual variability. J. Diary Sci. 2018, 101, 7236–7242. [Google Scholar] [CrossRef]
- Martuzzi, F.; Summer, A.; Formaggioni, P.; Mariani, P. Milk of Italian Saddle and Halfinger nursing mares: Physico-chemical characteristics, nitrogen composition and mineral elements at the end of lactation. Ital. J. Anim. Sci. 2004, 3, 293–299. [Google Scholar] [CrossRef]
- Mariani, P.; Summer, A.; Martuzzi, F.; Formaggioni, P.; Sabbioni, A.; Catalano, A.L. Physicochemical properties, gross composition, energy value and nitrogen fractions of Halfinger nursing mare milk throughout 6 lactation months. Anim. Res. 2001, 50, 415–425. [Google Scholar] [CrossRef]
- Burrows, A. Palette of our palates: A brief history of food coloring & its regulation. Comp. Rev. Food Sci. F. 2009, 8, 394–408. [Google Scholar] [CrossRef]
- Nozière, P.; Grolier, P.; Durand, D.; Ferlay, A.; Pradel, P.; Martin, B. Variations in carotenoids, fat-soluble micronutrients, and color in cows’ plasma and milk following changes in forage and feeding level. J. Dairy Sci. 2006, 89, 2634–2648. [Google Scholar] [CrossRef]
- Calderon, F.B.; Chaveau-Duriot, P.; Pradel, B.; Martin, B.; Graulet, M.; Doreau, P.; Noziere, P. Variations in carotenoids, vitamins A and E, and colour in cow’s plasma and milk following a shift from hay diet to diets containing increasing levels of carotenoids and vitamin E. J. Dairy Sci. 2007, 90, 5651–5664. [Google Scholar] [CrossRef]
- McDermott, A.; Visentin, G.; Mc Parland, S.; Berry, D.P.; Fenelon, M.A.; De Marchi, M. Effectiveness of mid-infrared spectroscopy to predict the colour of bovine milk and the relationship between milk color and traditional milk quality traits. J. Diary Sci. 2016, 99, 3267–3273. [Google Scholar] [CrossRef]
- Scarso, S.; McParland, S.; Vinsentin, G.; Berry, D.P.; McDermott, A.; De Marchi, M. Genetic and nongenetic factors associated with milk color in dairy cows. J. Diary Sci. 2017, 100, 7345–7361. [Google Scholar] [CrossRef]
- Lukefahr, S.; Hohenboken, W.D.; Cheeke, P.R.; Patton, N.M. Characterization of straightbred and crossbred rabbits for milk production and associative traits. J. Anim. Sci. 1983, 57, 1100–1107. [Google Scholar] [CrossRef]
- Chrastinova, L.; Sommer, A.; Rafay, J.; Svetlanska, M. Azotan exploitation in rabbit nutrition. II. Nutrient digestibility and lactation performance of does rabbit. J. Farm. Anim. Sci. 1997, 30, 80–86. [Google Scholar]
- El-Sabrout, K.; Aggag, S.; El-Raffa, A. Comparison of milk production and milk composition for an exotic and a local synthetic rabbit lines. Vet. World 2017, 10, 526–529. [Google Scholar] [CrossRef]
- Kolawole, F.S.O.; Kikelomo, A.M.; Taiwo, O.J.; Sunday, F.S. Yield, intake and chemical profile of Milk of Commercially available rabbits at first parity. Res. Rev. J. Agr. Allied. Sci. 2013, 20. [Google Scholar] [CrossRef]
- Langer, P. Differences in the composition of colostrum and milk in eutherians reflect differences in immunoglobulin transfer. J. Mammal. 2009, 90, 332–339. [Google Scholar] [CrossRef]
- Wyczling, P.; Gugołek, A.; Kowalska, D.; Kaliniewicz, J.; Strychalski, J.; Zwoliński, C. Effect of feeding diets with rapeseed cake and wheat dried distillers grain with solubles (DDGS) on reproductive performance and milk fatty acid profile in rabbit does (in Polish with English abstract). Rocz. Nauk. Zoot. 2013, 40, 65–76. [Google Scholar]
- Odegard, J.; Klemetsdal, G.; Heringstad, B. Genetic improvement of mastitis resistance: Validation of somatic cell score and clinical mastitis as selection criteria. J. Diary Sci. 2003, 86, 4129–4136. [Google Scholar] [CrossRef]
- Rainard, P.; Foucras, D.; Biochard, D.; Ruppa, R. Invited review: Low milk somatic cell count and susceptibility to mastitis. J. Diary Sci. 2018, 101, 6703–6714. [Google Scholar] [CrossRef] [PubMed]
- Raynal-Ljutovac, K.; Pirisi, A.; de Crémoux, R.; Gonzalo, C. Somatic cells of goat and sheep milk: Analytical, sanitary, productive and technological aspects. Small Rumin. Res. 2007, 68, 126–144. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Jaśkowski, J.M. Somatic cells in goat milk (in Polish with English abstract). Medycyna Wet. 2004, 60, 1263–1266. [Google Scholar]
- Zeng, S.S.; Escobar, E.M. Effect of breed and milking method on somatic cell count, standard plate count and composition of goat milk. Small Rumin. Res. 1996, 19, 169–175. [Google Scholar] [CrossRef]
- Paape, M.J.; Wiggans, G.R.; Bannerman, D.D.; Thomas, D.L.; Sanders, A.H.; Contreras, A.; Moroni, P.; Miller, R.H. Monitoring goat and milk sheep somatic cell counts. Small Rumin. Res. 2007, 68, 115–125. [Google Scholar] [CrossRef]
- Volek, Z.; Ebeid, T.A.; Uhlířová, L. The impact of substituting soybean and sunflower meal with a mixture of white lupine seeds and rapeseed meal on rabbit doe milk yield and composition, and the growth performance and carcass traits of their litters. Anim. Feed Sci. Tech. 2018, 236, 187–195. [Google Scholar] [CrossRef]
- Szendrő, Zs.; Gyarmati, T.; Maertens, L.; Radnai, I.; Biró-Németh, E.; Matics, Zs. Trials to produce rabbit young nursed by two does. 1. Milk production of does and milk intake of young. In Proceedings of the 7th World Rabbit Congress, Valencia, Spain, 4–7 July 2000; World Rabbit Science: Valencia, Spain, 2000; pp. 453–460. [Google Scholar]
- Fortun-Lamothe, L.; de Rochambeau, H.; Lebas, F.; Tudela, F. Influence of the number of suckling young on reproductive performance in intensively reared rabbit does. In Proceedings of the 7th World Rabbit Congress, Valencia, Spain, 4–7 July 2000; World Rabbit Science: Valencia, Spain; pp. 126–132. [Google Scholar]
- Szendrő, Zs.; Gyarmati, T.; Maertens, L.; Radnai, I.; Biró-Németh, E.; Matics, Zs. Trials to produce rabbit young nursed by two does. 2. Weight gain, body weight and mortality of young. In Proceedings of the 7th World Rabbit Congress, Valencia, Spain, 4–7 July 2000; World Rabbit Science: Valencia, Spain, 2000; pp. 461–467. [Google Scholar]
- Pasupathi, K.; Sakthivel, M.; Gopi, H.; Balasubramanyam, D.; Babu, M. Factors influencing pre-weaning growth of rabbits under farm condition. Int. J. Sci. Ent. 2014, 3, 2301–2302. [Google Scholar]
- Pałka, S.; Kmiecik, M.; Kozioł, K.; Otwinowska-Mindur, A.; Migdał, Ł.; Bieniek, J. The effect of breed on the litter size and milk yield in rabbits. Sci. Ann. PSAP 2017, 13, 25–29. [Google Scholar] [CrossRef]
- Kowalska, D.; Bielański, P. Effect of supplemental dietary fat for rabbits on milk composition and rearing performance of young rabbits. In Proceedings of the 8th World Rabbit Congress, Puebla, Mexico, 7–10 September 2004; World Rabbit Science: Valencia, Spain, 2004; pp. 869–873. [Google Scholar]
- Górska, A.; Mróz, B. Effect of somatic cell count on the amount of daily milk yield and chemical composition of milk from cows kept in the region of Southern Podlasie. Acta Sci. Pol. Zootechnica 2011, 10, 19–28. [Google Scholar]
- Bondan, C.; Folchini, J.A.; Noro, M.; Quadros, D.L.; Machado, K.M.; González, F.H.D. Milk composition of Holstein cows: A retrospective study. Cienc. Rural 2018, 48, e20180123. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Day | 0 | 2 | 6 | 12 | 17 | 21 | SEM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item/Litter Size | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | Day | Litter | |
| pH | 6.62 A | 6.61 A | 6.94 B | 6.87 B | 6.96 B | 6.93 B | 7.18 C | 7.16 C | 7.41 C | 7.40 C | 7.46 C | 7.44 C | 0.02 | 0.001 | 0.542 |
| L* | 68.37 A | 68.75 A | 69.98 A | 69.87 A | 70.07 AB | 70.12 AB | 70.38 B | 70.49 B | 71.31 B | 71.43 B | 70.88 B | 70.70 B | 0.13 | <0.001 | 0.865 |
| a* | −0.40 A | −0.43 A | −0.45 A | −0.48 A | −0.62 B | −0.58 B | −0.47 A | −0.48 A | −0.98 C | −0.86 C | −1.34 D | −1.26 D | 0.03 | <0.001 | 0.534 |
| b* | 4.68 A | 4.53 A | 3.84 B | 3.67 B | 3.96 B | 4.07 B | 4.57 A | 4.62 A | 4.70 A | 4.42 A | 5.17 C | 5.25 C | 0.03 | <0.001 | 0.202 |
| SCC (103 mL−1) | 523.67 A | 536.57 A | 435.67 B | 421.31 B | 184.00 C | 196.55 C | 155.67 D | 162.78 D | 104.50 E | 98 E | 71.28 F | 77.58 F | 11.1 | <0.001 | 0.397 |
| Acidity | 25.17 A | 24.81 A | 20.51 A | 20.78 A | 17.23 AB | 17.42 AB | 18.94 AB | 18.64 AB | 14.67 B | 14.57 B | 20.02 A | 19.79 A | 0.29 | <0.001 | 0.352 |
| Density | 1.027 | 1.033 | 1.033 | 1.035 | 1.039 | 1.038 | 1.042 | 1.045 | 1.043 | 1.043 | 1.045 | 1.049 | 0.005 | 0.151 | 0.348 |
| Day | 0 | 2 | 6 | 12 | 17 | 21 | SEM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item/Litter Size | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | Day | Litter | |
| TS (g kg−1) | 31.54 A | 31.80 A | 28.31 B | 28.15 B | 29.88 C | 29.63 C | 28.53 B | 28.12 B | 26.39 D | 26.41 D | 27.63 E | 27.96 E | 0.24 | <0.001 | 0.496 |
| Protein (g kg−1) | 11.01 A | 11.13 A | 11.07 A | 11.06 A | 11.05 A | 11.02 A | 11.41 B | 11.36 B | 10.93 EC | 10.89 C | 12.40 D | 12.30 D | 0.12 | <0.001 | 0.172 |
| Casein (g kg−1) | 8.43 A | 8.34 Aa | 8.60 A | 8.81 Ab | 8.98 AB | 8.79 Ab | 9.33 C | 9.14 Aa | 8.26 A | 8.85 Ab | 9.86 D | 9.75 D | 0.23 | 0.002 | 0.379 |
| Fat (g kg−1) | 15.73 A | 15.69 A | 13.66 B | 13.55 B | 15.58 C | 15.71 C | 12.82 Da | 12.92 Da | 11.63 E | 11.26 E | 12.16 Db | 12.09 Db | 0.20 | <0.001 | 0.115 |
| SNF (g kg−1) | 14.79 A | 14.87 A | 15.81 B | 15.75 B | 16.41 C | 16.94 C | 17.55 D | 17.82 Da | 17.31 Db | 17.93 Da | 18.22 E | 18.11 E | 0.15 | <0.001 | 0.162 |
| Lactose (g kg−1) | 1.99 A | 2.00 A | 2.01 A | 2.02 A | 2.26 B | 2.24 B | 2.69 C | 2.64 C | 2.72 C | 2.69 C | 2.30 B | 2.24 B | 0.02 | <0.001 | 0.251 |
| Days | 2 | 6 | 12 | 17 | 21 | SEM | p-value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item/Litter Size | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | 8 | 10 | Day | Litter Size | |
| Body Weight of the Doe (g) | 4629 a | 4663 a | 4977 ab | 4981 ab | 5024 b | 5067 b | 5117 b | 5192 b | 4880 ab | 4928 ab | 24.1 | 0.013 | 0.207 |
| Daily Milk Production (g) | 100.4 A | 120.0 B | 198.0 C | 224.0 Da | 237.1 Db | 242.7 DE | 303.6 F | 332.3 G | 246.0 H | 270.9 I | 4.8 | <0.001 | <0.001 |
| Intake by One Marked kit (g) | 11.8 A | 12.2 A | 26.0 Ba | 25.6 Bb | 26.5 Bb | 25.0 C | 28.6 D | 29.0 D | 26.8 Bb | 25.1 C | 0.6 | <0.001 | 0.007 |
| Item/Litter Size | 8 | 10 | SEM |
|---|---|---|---|
| Litter Weight Gain (g) | 2207 A | 2794 B | 32 |
| Weight gain of the Marked Kit (g) | 301.8 | 297.8 | 4.5 |
| Milk Yield Coefficient | 3.65 A | 3.82 B | 0.01 |
| Trait | pH | L* | a* | b* | SCC | Protein | Casein | Fat | TS | SNF | Lactose | Acidity |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L* | 0.750 (0.003) | |||||||||||
| a* | −0.665 (0.010) | −0.564 (0.025) | ||||||||||
| b* | 0.510 (0.034) | 0.370 (0.045) | −0.371 (0.045) | |||||||||
| SCC | −0.573 (0.022) | −0.486 (0.037) | 0.360 (0.051) | −0.319 (0.083) | ||||||||
| Protein | 0.258 (0.220) | 0.094 (0.769) | −0.329 (0.084) | 0.331 (0.078) | −0.093 (0.769) | |||||||
| Casein | 0.318 (0.083) | 0.227 (0.220) | −0.301 (0.093) | 0.358 (0.070) | −0.215 (0.223) | 0.916 (<0.001) | ||||||
| Fat | −0.652 (0.013) | −0.412 (0.038) | 0.548 (0.028) | −0.322 (0.082) | 0.279 (0.221) | −0.484 (0.038) | −0.293 (0.110) | |||||
| TS | −0.614 (0.018) | −0.431 (0.037) | 0.496 (0.032) | −0.225 (0.220) | 0.273 (0.199) | −0.085 (0.771) | 0.088 (0.772) | 0.886 (0.001) | ||||
| SNF | 0.456 (0.030) | 0.337 (0.067) | −0.311 (0.085) | 0.351 (0.069) | −0.343 (0.075) | 0.790 (0.002) | 0.838 (0.001) | −0.484 (0.036) | −0.037 (0.812) | |||
| Lactose | 0.393 (0.039) | 0.370 (0.047) | −0.085 (0.771) | 0.200 (0.228) | −0.387 (0.045) | 0.067 (0.773) | 0.285 (0.217) | −0.082 (0.755) | 0.063 (0.773) | 0.491 (0.031) | ||
| Acidity | −0.425 (0.042) | −0.451 (0.039) | 0.182 (0.360) | −0.038 (0.812) | 0.389 (0.045) | 0.569 (0.024) | 0.450 (0.041) | 0.156 (0.376) | 0.480 (0.038) | 0.304 (0.093) | −0.181 (0.368) | |
| Daily milk production | 0.377 (0.047) | 0.425 (0.042) | −0.126 (0.411) | 0.311 (0.085) | −0.331 (0.078) | −0.120 (0.409) | −0.110 (0.442) | −0.170 (0.407) | −0.229 (0.239) | 0.075 (-0.769) | 0.369 (0.064) | −0.364 (0.064) |
| Trait | DM | LB | IM | BWD | LW | MW | LG | MG |
|---|---|---|---|---|---|---|---|---|
| LB | 0.114 (0.441) | |||||||
| IM | 0.384 (0.051) | −0.123 (0.320) | ||||||
| BWD | 0.451 (0.040) | 0.139 (0.415) | 0.034 (0.813) | |||||
| LW | 0.538 (0.028) | 0.093 (0.734) | 0.278 (0.234) | 0.393 (0.050) | ||||
| MW | 0.467 (0.036) | −0.004 (0.991) | 0.812 (0.001) | 0.284 (0.240) | 0.898 (0.001) | |||
| LG | 0.441 (0.045) | 0.320 (0.091) | 0.074 (0.768) | 0.346 (0.068) | 0.969 (0.001) | 0.560 (0.020) | ||
| MG | 0.353 (0.053) | 0.284 (0.239) | 0.095 (0.734) | 0.318 (0.090) | 0.531 (0.029) | 0.956 (0.001) | 0.546 (0.021) | |
| MY | 0.038 (0.815) | 0.282 (0.239) | 0.128 (0.320) | 0.102 (0.449) | 0.673 (0.012) | 0.320 (0.090) | 0.831 (0.001) | 0.449 (0.040) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ludwiczak, A.; Składanowska-Baryza, J.; Kuczyńska, B.; Stanisz, M. Hycole Doe Milk Properties and Kit Growth. Animals 2020, 10, 214. https://doi.org/10.3390/ani10020214
Ludwiczak A, Składanowska-Baryza J, Kuczyńska B, Stanisz M. Hycole Doe Milk Properties and Kit Growth. Animals. 2020; 10(2):214. https://doi.org/10.3390/ani10020214
Chicago/Turabian StyleLudwiczak, Agnieszka, Joanna Składanowska-Baryza, Beata Kuczyńska, and Marek Stanisz. 2020. "Hycole Doe Milk Properties and Kit Growth" Animals 10, no. 2: 214. https://doi.org/10.3390/ani10020214
APA StyleLudwiczak, A., Składanowska-Baryza, J., Kuczyńska, B., & Stanisz, M. (2020). Hycole Doe Milk Properties and Kit Growth. Animals, 10(2), 214. https://doi.org/10.3390/ani10020214