Porcine Digestible Peptides (PDP) in Weanling Diets Regulates the Expression of Genes Involved in Gut Barrier Function, Immune Response and Nutrient Transport in Nursery Pigs

 , , , ,
, , , ,  , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing and Diet
2.1.1. Trial 1
2.1.2. Trial 2
2.2. Data and Sample Collection
2.3. Proximate Analysis of Diets
2.4. Gene Expression Study by qPCR
2.5. Open Array Design
2.6. RNA Extraction and cDNA Preparation
2.7. Gene Expression Data Analysis
2.8. Performance Data Statistical Analysis
3. Results
3.1. Growth Performance
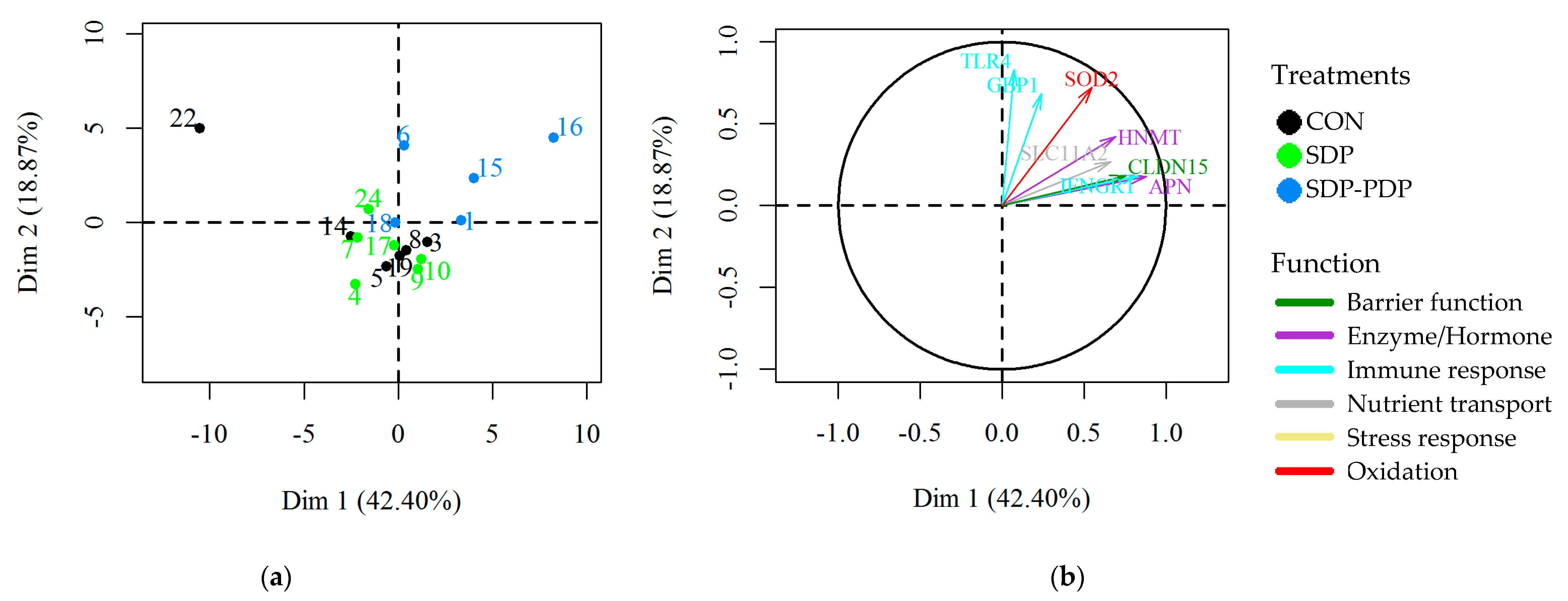
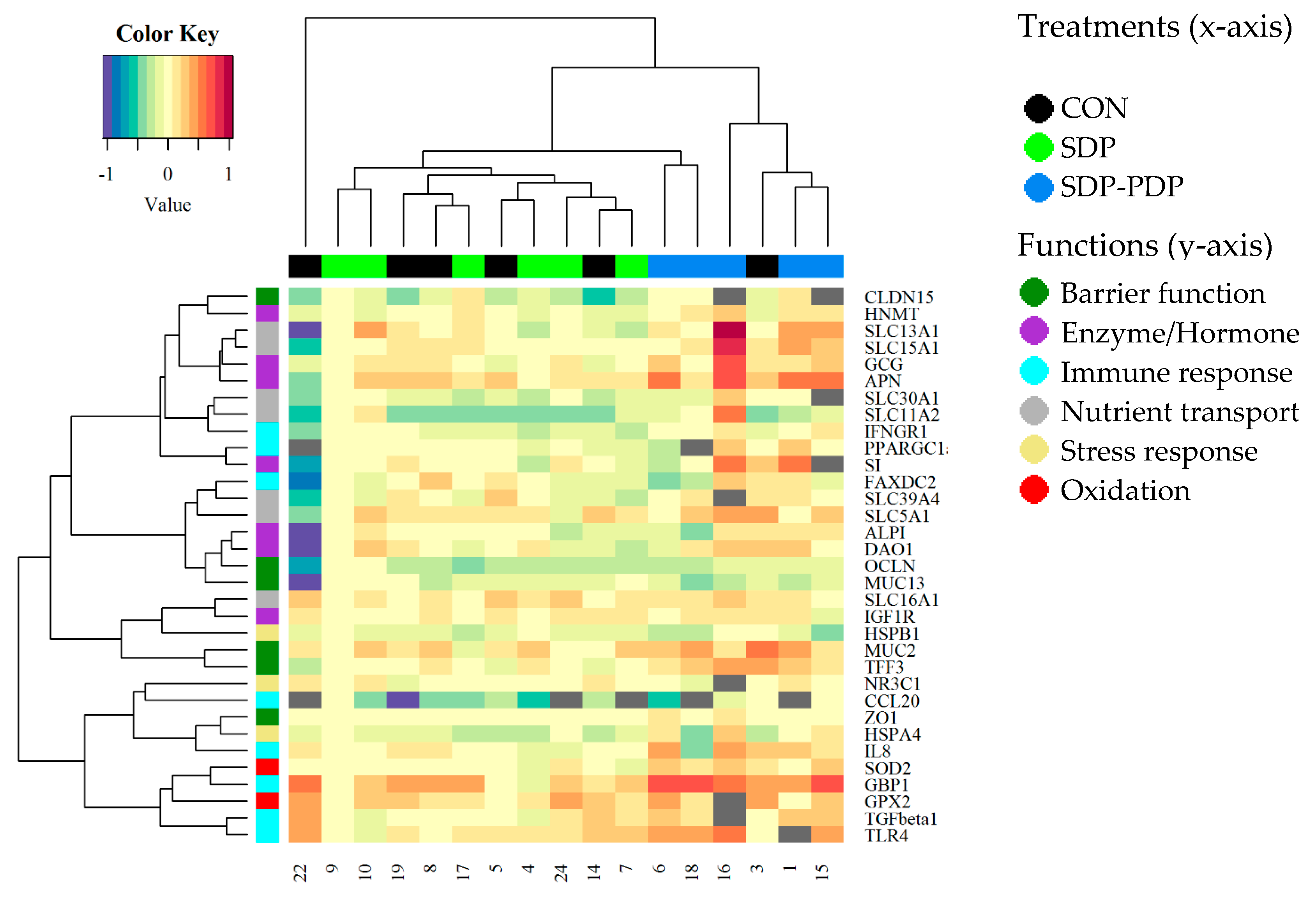
3.2. Intestinal Mucosa Gene Expression Values
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Gene | Name | Primer Forward (5′-3′) | Primer Reverse (5′-3′) | Probe (5′-3′) |
|---|---|---|---|---|
| OCLN | Occludin | CAGGTGCACCCTCCAGATTG | CAGGCCTATAAGGAGGTGGACTT | TGACATCAGCCATGTCAT |
| ZO1 | Zonula occludens 1 | GCTATGTCCAGAATCTCGGAAAA | TGCTTCTTTCAATGCTCCATACC | TCACCATCTTTTTACAACTAC |
| CLDN1 | Claudin-1 | CTTCGACTCCTTGCTGAATCTGA | CTTCCATGCACTTCATACACTTCAT | ACAGCACTTTGCAAGC |
| CLDN4 | Claudin-4 | CCTCCGTGCTGTTCCTCAA | GAGGCACAAGCCCAGCAA | CCTTGTGGCACTTTG |
| CLDN15 | Claudin-15 | GCTATCTCCTGGTATGCCTTCAA | GGGACTTCCACACTCCTTGGT | ACTTCTTCGACCCCTTGTA |
| MUC2 | Mucin 2 | AAGGACGACACCATCTACCTCACT | GGCCAGCTCGGGAATAGAC | CATGGTCAGCACCCCG |
| MUC13 | Mucin 13 | CAGTGGAGTTGGCTGTGAAAAC | ATCAAGTTCTGTTCTTCCACATTCTTG | TCCTCTCATTAAGATCAAAC |
| TFF3 | Trefoil factor 3 | AGAACCTGCCCGTGACCAT | CACACTGGTTCGCCGACAG | AGGCCAGGATGTTCT |
| TLR2 | Toll-like receptor 2 | CTCTCGTTGCGGCTTCCA | AAGACCCATGCTGTCCACAAA | CAAGGTCAACTCTCTG |
| TLR4 | Toll-like receptor 4 | CATCCCCACATCAGTCAAGATACT | TCAATTGTCTGAATTTCACATCTGG | ACAGCAATAGCTTCTCCA |
| IL1B | Interleukin 1 beta | GGTGACAACAATAATGACCTGTTATTTG | GCTCCCATTTCTCAGAGAACCA | ATGAAGTGCTGCACCC |
| IL6 | Interleukin 6 | CCAATCTGGGTTCAATCAGGAG | ACAGCCTCGACATTTCCCTTATT | AGATATACCTGGACTACCTC |
| IL8 | Interleukin 8 | GGAAAAGTGGGTGCAGAAGGT | GAGAATGGGTTTTTGCTTGTTGT | TACAGATATTTTTGAAGAGAACT |
| IL10 | Interleukin 10 | TGAGGCTGCGGCGCT | GAGCTTGCTAAAGGCACTCTTCA | AACAAGAGCAAGGCCGT |
| IL17A | Interleukin 17 | CCAGACGGCCCTCAGATTAC | ATCTTCCTTCCCTTCAGCATTG | CCATGGACTCTCCAACG |
| IL22 | Interleukin 22 | TGTTCCCCAACTCTGATAGATTCC | GTTGTTCACATTTCTCTGGATATGCT | AGCTAAGCCAATGCCGTAT |
| IFNG | Interferon gamma | TGACTTTGTGTTTTTCTGGCTCTT | CACTCTCCTCTTTCCAATTCTTCAA | ATCCTAAAGGACTATTTTAAT |
| TNF | Tumor necrosis factor alpha | CACCACGCTCTTCTGCCTACT | GACGGGCTTATCTGAGGTTTGA | CAAGGACTCAGATCATCGT |
| TGFB1 | Transforming growth factor beta 1 | GCGGCAGCTCTACATTGACTT | GACCTTGCTGTACTGAGTGTCTAGG | CCATGCCAATTTCTGCCT |
| CCL20 | Chemokine (C-C motif) ligand 20 | GACCATATTCTTCACCCCAGATTT | CACACACGGCTAACTTTTTCTTTG | ATCAATGCAATCATCTTT |
| CXCL2 | Chemokine (C-X-C motif) ligand 2 | CATGGTGAAGAAAATCATCGAGAA | GCCAGTAAGTTTCCTCCATCTCTCT | AACAAGAGCAGTGCCAAC |
| IFNGR1 | Interferon gamma receptor 1 | CATGTTACCCAAATCTTTGCTGTCT | CAGTATGCACGCTTGAAATTGTC | ATATATATCACCCATCACCTACC |
| HSPB1 | Heat shock protein 27 | CGAGGAGCTGACGGTCAAG | GCAGCGTGTATTTTCGAGTGAA | ACGGCTTCATTTCCCGGT |
| HSPA4 | Heat shock protein 70 | TCAATTGCCTGCGATTAATGAA | GAATGCCCCATGTCTACAAAAAC | CAGTTGCTCTTGCATATG |
| REG3G | Regenerating-islet derived protein 3 gamma | TGCCTGATGCTCCTGTCTCA | GGCATAGCAGTAGGAAGCATAGG | CCAAGGTGAAGATTC |
| PPARGC1A | Peroxisome proliferative activated receptor gamma, coactivator 1 alpha | CTCTGGAACTGCAGGCCTAA | TGGAGAAGCCCTAAAAGGGTTAT | ACCCACAACTCCTCCT |
| FAXDC2 | Fatty acid hydrolase domain containing 2 | CCATGACTACCACCATCTCAAGTT | GCAGGATCGTGTGTCTCTCGTA | TGTTCAAGCAGACCAAG |
| GBP1 | Guanylate binding protein 1 | AGAATCCATCACAGCAGACGAGTA | CGGATACAGAGTCGAGGCAGGTTAA | TCAAGCTTAAGAAGGGTACCAG |
| GPX2 | Glutathione peroxidase 2 | CAACCAATTTGGACATCAGGAG | GGGTAAAGGTGGGCTGGAAT | AGATCCTGAACAGCCTCA |
| SOD2 | Superoxide dismutase | GGGTTGGCTCGGTTTCAA | CATGCTCCCACACGTCGAT | CTGCAAGGAACAACAGGTCT |
| ALPI | Intestinal alkaline phosphatase | ATGTCTTCTCTTTTGGTGGCTACA | GGAGGTATATGGCTTGAGATCCA | AAGCTCCGTTTTTGGCCT |
| SI | Sucrase-isomaltase | CGACCCCTTTTGCATGAGTT | AAGGCTGGACCCCATAGGAA | TTTAATGAAAAGCCAACCTG |
| DAO1 | Diamine oxidase | GGAACCAACAGACCTTCAACTATCTC | TTCGGAATCCCAGGACCAT | CCGGACCCTTACTGGAAA |
| HNMT | Histamine N-methyltransferase | TGTTGAACCAAGTGCTGAACAAAT | ACTTTATGTTCTCGAGGTTTGATGTCTT | ACCAAGTACAAAGAGCTT |
| APN | Aminopeptidase-N | AGGGCAACGTCAAAAAGGTG | GTCAAAGCATGGGAAGGATTTC | ACACAGATGCAGTCTACAG |
| IDO1 | Indoleamine 2,3 dioxygenase | TTGGCAAATTGGAAGAAAAAGG | CCGGAAATGAGAAGAGAATATCCAT | CCAGTGGGCCCATGACTTAC |
| GCG | Glucagon | AGGCGTGCCCAGGATTTT | CATCGTGACGTTTGGCAATG | CACCAAGAGGAACAAGAA |
| CCK | Cholecystokinin | CAGCAGGCTCGAAAAGCAC | AATCCATCCAGCCCATGTAGTC | CAGCCACAGAATAAGTGA |
| IGF1R | Insulin-like growth factor 1 receptor | CCGACGCGGCAACAAC | TCAGGAAGGACAAGGAGACCAA | CTACGTGAAGATCCGCCA |
| PYY | Peptide tyrosine tyrosine | CAGAGGTATGGGAAACGTGACA | CCTTCTGGCCACGACTTGAC | CAAACTGCTCTTCCCTGAA |
| SLC5A1 | Solute carrier family 5 (sodium/glucose cotransporter) member 1 | GGCCATCTTTCTCTTACTGGCA | TCCCACTTCATGAAAAGCAAAC | TTTATACGGATACCTTGCAGAC |
| SLC16A1 | Monocarboxylate transporter 1 | CCTTGTTGGACCTCAGAGATTCTC | CCAGTATGTGTATTTATAGTCTCCGTATATGTC | CCACCACTTTTAGGTCGTC |
| SLC7A8 | Solute carrier family 7 (amino acid transporter light chain, L System) member 8 | TGTCGCTTATGTCACTGCAATGT | GACAGGGCGACGGAAATG | CTGTGACTTTTGGAGAGAA |
| SLC15A1 | Solute carrier family 15 (oligopeptide transporter) member 1 | GGTTATCCCTTGAGCATCTTCTTC | AGTGCTCTCATTCCATAGTAGGAAAA | TCAACGAGTTCTGTGAAAG |
| SLC13A1 | Solute carrier family 13 (sodium/sulfate symporters) member 1 | GGTACCTCCACCAACTTGATCTTC | ATCCAAAGTTGATGCAGTGACAAT | ATTTCAATATGCGCTACCC |
| SLC11A2 | Solute carrier family 11 (proton-coupled divalent metal ion transporter) member 2 | GTCTTTGCCGAAGCGTTTTTT | ACCACGCCCCCTTTGTAGA | CCAACCAGCAGGTGGT |
| MT1A | Metallothionein 1A | TGAATCCGCGTTGCTCTCT | CAGGAGCAGCAGCTCTTCTT | ACGTGCAAAACCTGCAGA |
| SLC30A1 | Solute carrier family 30 (zinc transporter) member 1 | AATTGGACCGGACAGATCCA | TCTCTGATAAGATTCCCATTCACTTG | AAAAGTCCAGAAGTGATGC |
| SLC39A4 | Solute carrier family 39 (zinc transporter) member 4 | ATCTTTGGGCTCTTGCTCCTT | GCAGCCCCAGCACCTTAG | CTGCTACCCACTACGTCA |
| CRHR1 | Corticotropin releasing hormone receptor 1 | CAGGGCCCCATGATATTGG | CCGGAGTTTGGTCATGAGGAT | CTGATCAACTTTATCTTCC |
| NR3C1 | Glucocorticoid receptor | GGCAATACCAGGATTCAGGAACT | CCATGAGAAACATCCATGAATACTG | TGACCAAATGACCCTCCT |
| HSD11B1 | Hydroxysteroid (11-beta) dehydrogenase 1 | GGTCAGAAGAAACTCTCAAGAAGGTG | GCGAAGGTCATGTCCTCCAT | TCTTCAGCACACTACGTTG |
| GAPDH | Glyceraldehyde-phosphate-dehydrogenase | TTCGTCAAGCTCATTTCCTGGTA | TCCTCGCGTGCTCTTGCT | AATTTGGCTACAGCAACAG |
| ACTB | Actin, beta | CAAGGACCTCTACGCCAACAC | TGGAGGCGCGATGATCTT | CACCACCATGTACCCAGG |
| TBP | TATA-box binding protein | CAGAATGATCAAACCGAGAATTGT | CTGCTCTGACTTTAGCACCTGTTAA | TTTGTCTCTGGAAAAGTTGT |
| B2M | Beta-2-microglobulin | TCACTCCTAACGCTGTGGATCA | CGGTTAGTGGTCTCGATCCC | AGCACGTGACTCTCGATA |
References
- Lallès, J.P.; Bosi, P.; Smidt, H.; Stokes, C.R. Weaning—A Challenge to Gut Physiologists. Livest. Sci. 2007, 108, 82–93. [Google Scholar] [CrossRef]
- Domeneghini, C.; Di Giancamillo, A.; Arrighi, S.; Bosi, G. Gut-Trophic Feed Additives and Their Effects upon the Gut Structure and Intestinal Metabolism. State of the Art in the Pig, and Perspectives towards Humans. Histol. Histopathol. 2006, 21, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 851–873. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R. Feed- and Feed Additives-Related Aspects of Gut Health and Development in Weanling Pigs. J. Anim. Sci. Biotechnol. 2013, 4, 1. [Google Scholar] [CrossRef]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive Peptides of Animal Origin: A Review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef] [PubMed]
- Bosi, P.; Casini, L.; Finamore, A.; Cremokolini, C.; Merialdi, G.; Trevisi, P.; Nobili, F.; Mengheri, E. Spray-Dried Plasma Improves Growth Performance and Reduces Inflammatory Status of Weaned Pigs Challenged with Enterotoxigenic Escherichia Coli K88. J. Anim. Sci 2004, 82, 1764–1772. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Miró, L.; Polo, J.; Russell, L.; Campbell, J.; Weaver, E.; Crenshaw, J.; Moretó, M. Dietary Plasma Proteins Modulate the Immune Response of Diffuse Gut-Associated Lymphoid Tissue in Rats Challenged with Staphylococcus Aureus Enterotoxin B. J. Nutr. 2018, 138, 533–537. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Amat, C.; Polo, J.; Campbell, J.M.; Crenshaw, J.; Russell, L.; Moretó, M. Spray-Dried Animal Plasma Prevents the Effects of Staphylococcus Aureus Enterotoxin B on Intestinal Barrier Function in Weaned Rats. J. Nutr. 2018, 136, 2838–2843. [Google Scholar] [CrossRef]
- Figueroa, J.; Solà-Oriol, D.; Guzmán-Pino, S.A.; Chetrit, C.; Borda, E.; Pérez, J.F. The Use of Porcine Digestible Peptides and Their Continuity Effect in Nursery Pigs. J. Anim. Sci. 2016, 94, 1531–1540. [Google Scholar] [CrossRef][Green Version]
- Solà-Oriol, D.; Roura, E.; Torrallardona, D. Feed Preference in Pigs: Effect of Selected Protein, Fat, and Fiber Sources at Different Inclusion Rates. J. Anim. Sci. 2011, 89, 3219–3227. [Google Scholar] [CrossRef]
- Xin, G.; QiuGang, M.; Cheng, J.; ShaoChen, G.; Keller, G. Evaluation of DPS Supplement in Early-Weaned Pig Diets. Acta Zoonutrimenta Sin. 2001, 13, 15–19. [Google Scholar]
- FEDNA. Necesidades Nutricionales Para Ganado Porcino, 2nd ed.; FEDNA: Madrid, Spain, 2013. [Google Scholar]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Official Methods of Analysis of the AOAC, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- AOAC International. Official Methods of Analysis of AOAC International, 18th ed.; Official Methods of Analysis of AOAC International: Gaithersburg, MI, USA, 2007. [Google Scholar]
- AOAC International. Official Methods of Analysis of AOAC International, 18th ed.; Official Methods of Analysis of AOAC International: Washington, DC, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Garrick, M.D.; Dolan, K.G.; Horbinski, C.; Ghio, A.J.; Higgins, D.; Porubcin, M.; Moore, E.G.; Hainsworth, L.N.; Umbreit, J.N.; Conrad, M.E.; et al. DMT1: A Mammalian Transporter for Multiple Metals. BioMetals 2003, 16, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like Receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Liu, P.; Pieper, R.; Rieger, J.; Vahjen, W.; Davin, R.; Plendl, J.; Meyer, W.; Zentek, J. Effect of Dietary Zinc Oxide on Morphological Characteristics, Mucin Composition and Gene Expression in the Colon of Weaned Piglets. PLoS ONE 2014, 9, e91091. [Google Scholar] [CrossRef]
- Smith, S.H.; Wilson, A.D.; Van Ettinger, I.; Macintyre, N.; Archibald, A.L.; Ait-Ali, T. Down-Regulation of Mechanisms Involved in Cell Transport and Maintenance of Mucosal Integrity in Pigs Infected with Lawsonia Intracellularis. Vet. Res. 2014, 45. [Google Scholar] [CrossRef]
- Khan, N.; Asif, A.R. Transcriptional Regulators of Claudins in Epithelial Tight Junctions. Mediat. Inflamm. 2015, 2015. [Google Scholar] [CrossRef]
- Xu, S.; Wang, D.; Zhang, P.; Lin, Y.; Fang, Z.; Che, L.; Wu, D. Oral Administration of Lactococcus lactis-Expressed Recombinant Porcine Epidermal Growth Factor Stimulates the Development and Promotes the Health of Small Intestines in Early-Weaned Piglets. J. Appl. Microbiol. 2015, 119, 225–235. [Google Scholar] [CrossRef]
- Taupin, D.; Podolsky, D.K. Erratum: Trefoil Factors: Initiators of Mucosal Healing. Nat. Rev. Mol. Cell Biol. 2003, 4, 721–732. [Google Scholar] [CrossRef]
- Tamura, A.; Kitano, Y.; Hata, M.; Katsuno, T.; Moriwaki, K.; Sasaki, H.; Hayashi, H.; Suzuki, Y.; Noda, T.; Furuse, M.; et al. Megaintestine in Claudin-15-Deficient Mice. Gastroenterology 2008, 134. [Google Scholar] [CrossRef]
- Hansen, S.L.; Trakooljul, N.; Liu, H.-C.; Moeser, A.J.; Spears, J.W. Iron Transporters Are Differentially Regulated by Dietary Iron, and Modifications Are Associated with Changes in Manganese Metabolism in Young Pigs. J. Nutr. 2009, 139, 1474–1479. [Google Scholar] [CrossRef] [PubMed]
- Schnoor, M.; Betanzos, A.; Weber, D.A.; Parkos, C.A. Guanylate-Binding Protein-1 Is Expressed at Tight Junctions of Intestinal Epithelial Cells in Response to Interferon-Gamma and Regulates Barrier Function through Effects on Apoptosis. Mucosal Immunol. 2009, 2, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Badia, R.; Lizardo, R.; Martinez, P.; Badiola, I.; Brufau, J. The Influence of Dietary Locust Bean Gum and Live Yeast on Some Digestive Immunological Parameters of Piglets Experimentally Challenged with Escherichia Coli1. J. Anim. Sci. 2012, 90, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, Z.; Jiang, J.; Yu, Y.; Zhang, Q. Differential Gene Expression Profiling of Porcine Epithelial Cells Infected with Three Enterotoxigenic Escherichia Coli Strains. BMC Genom. 2012, 13, 330. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, M.; Łosiewicz, K.; Podlasz, P.; Wasowicz, K. The Expression of Mitochondrial, Cytoplasmic and Extracellular Superoxide Dismutase in the Colonic Wall of Pigs Suffering from Swine Dysenteria. Pol. J. Vet. Sci. 2013, 16, 463–467. [Google Scholar] [CrossRef]
- Kröger, S.; Pieper, R.; Schwelberger, H.G.; Wang, J.; Villodre Tudela, C.; Aschenbach, J.R.; Van Kessel, A.G.; Zentek, J. Diets High in Heat-Treated Soybean Meal Reduce the Histamine-Induced Epithelial Response in the Colon of Weaned Piglets and Increase Epithelial Catabolism of Histamine. PLoS ONE 2013, 8, e80612. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. [2.12.2]; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open Software Development for Computational Biology and Bioinformatics. Genome Biol. 2004, 5. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate. J. R. Stat. Soc. B 1995, 289–300. [Google Scholar] [CrossRef]
- Tukey, J.W. Comparing Individual Means in the Analysis of Variance. Biometrics 1949, 5, 99. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Agglomerative Clustering Method: Which Algorithms Implement Ward’s Criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef]
- Everitt, B. Cluster Analysis; Heinemann Educational Books: London, UK, 1974. [Google Scholar] [CrossRef]
- Warnes, G.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. Gplots: Various R Programming Tools for Plotting Data. R Package Version 3.0.1.1; Science Open, Inc.: Berlin, Germany, 2015. [Google Scholar]
- Carbonaro, M.; Maselli, P.; Nucara, A. Relationship between Digestibility and Secondary Structure of Raw and Thermally Treated Legume Proteins: A Fourier Transform Infrared (FT-IR) Spectroscopic Study. Amino Acids 2012, 43, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Villodre Tudela, C.; Taciak, M.; Bindelle, J.; Pérez, J.F.; Zentek, J. Health Relevance of Intestinal Protein Fermentation in Young Pigs. Anim. Heal. Res. Rev. 2016, 17, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Li, D.F.; Nelssen, J.L.; Reddy, P.G.; Blecha, F.; Klemm, R.D.; Giesting, D.W.; Hancock, J.D.; Allee, G.L.; Goodband, R.D. Measuring Suitability of Soybean Products for Early-Weaned Pigs with Immunological Criteria. J. Anim. Sci. 1991, 69, 3299–3307. [Google Scholar] [CrossRef] [PubMed]
- Salgado, P.; Freire, J.P.B.; Mourato, M.; Cabral, F.; Toullec, R.; Lallès, J.P. Comparative Effects of Different Legume Protein Sources in Weaned Piglets: Nutrient Digestibility, Intestinal Morphology and Digestive Enzymes. Livest. Prod. Sci. 2002, 74, 191–202. [Google Scholar] [CrossRef]
- Prohászka, L.; Baron, F. The Predisposing Role of High Dietary Protein Supplies in Enteropathogenic E. coli Infections of Weaned Pigs. Zent. Veterinärmedizin R. B 1980, 27, 222–232. [Google Scholar] [CrossRef]
- Solà-Oriol, D.; Figueroa, J.; Borda, E.; Chetrit, C.; Pérez, J.F. The Effect of Different Animal and Vegetable Protein Sources on the Feed Intake and Weight Gain of Piglets. J. Anim. Sci. 2010, 88, 92. [Google Scholar]
- Martínez-Puig, D.; Anguita, M.; Baucells, F.; Borda, E.; Pérez, J.; Chetrit, C. Evidence of a Preference in Piglets for an Animal Protein Hydrolysate. In Proceedings of the Joint Annual Meeting ADSA-PSA-AMPA-ASAS, San Antonio, TX, USA, 8–12 July 2007; Journal of Animal Science: San Antonio, TX, USA, 2007; Volume 85, p. 310. [Google Scholar]
- Torrallardona, D. Spray Dried Animal Plasma as an Alternative to Antibiotics in Weanling Pigs. Asian-Australas. J. Anim. Sci. 2010, 23, 131–148. [Google Scholar] [CrossRef]
- Ermer, P.M.; Miller, P.S.; Lewis, A.J. Diet Preference and Meal Patterns of Weanling Pigs Offered Diets Containing Either Spray-Dried Porcine Plasma or Dried Skim Milk. J. Anim. Sci. 1994, 72, 1548–1554. [Google Scholar] [CrossRef]
- Pérez-Bosque, A.; Polo, J.; Torrallardona, D. Spray Dried Plasma as an Alternative to Antibiotics in Piglet Feeds, Mode of Action and Biosafety. Porc. Health Manag. 2016, 2, 1–10. [Google Scholar] [CrossRef]
- Myers, A.J.; Goodband, R.D.; Tokach, M.D.; Dritz, S.S.; DeRouchey, J.M.; Nelssen, J.L. The Effects of Porcine Intestinal Mucosa Protein Sources on Nursery Pig Growth Performance. J. Anim. Sci. 2014, 92, 783–792. [Google Scholar] [CrossRef]
- Borda, E.; Martínez-Puig, D.; Pérez, J. An Hydrolyzed Protein Concentrate (Palbio 62) Increases Feed Intake and Villus Height in Early Weaning Pigs. In Book of Abstracts of the 56th Annual Meeting of the EAAP; Wageningen Academic Publishers: Uppsala, Sweden, 2005. [Google Scholar]
- Gilbert, E.R.; Wong, E.A.; Webb, K.E. Board-Invited Review: Peptide Absorption and Utilization: Implications for Animal Nutrition and Health. J. Anim. Sci. 2008, 86, 2135–2155. [Google Scholar] [CrossRef] [PubMed]
- Rérat, A.; Nunes, C.S.; Mendy, F.; Roger, L. Amino Acid Absorption and Production of Pancreatic Hormones in Non-Anaesthetized Pigs after Duodenal Infusions of a Milk Enzymic Hydrolysate or of Free Amino Acids. Br. J. Nutr. 1988, 60, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Jiang, C.; Liang, S.; Kang, L.; Jiang, Y. Genomic Structure and Characterization of MRNA Expression Pattern of Porcine Interferon Gamma Receptor 1 Gene. Int. J. Immunogenet. 2010, 37, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Bank, U.; Bohr, U.R.M.; Reinhold, D.; Lendeckel, U.; Ansorge, S.; Malfertheiner, P.; Täger, M. Inflammatory Bowel Diseases: Multiple Benefits from Therapy with Dipepridyl- and Alanyl-Aminopeptidase Inhibitors. Front. Biosci. 2008, 3699–3713. [Google Scholar] [CrossRef]
- Zhao, L.; Bartnikas, T.; Chu, X.; Klein, J.; Yun, C.; Srinivasan, S.; He, P. Hyperglycemia Promotes Microvillus Membrane Expression of DMT1 in Intestinal Epithelial Cells in a PKCα-Dependent Manner. FASEB J. 2019, 33, 3549–3561. [Google Scholar] [CrossRef]
- Li, C.; Zhou, H.M. The Role of Manganese Superoxide Dismutase in Inflammation Defense. Enzym. Res. 2011, 2011, 387176. [Google Scholar] [CrossRef]
- Visner, G.A.; Dougall, W.C.; Wilson, J.M.; Burr, I.A.; Nick, H.S. Regulation of Manganese Superoxide Dismutase by Lipopolysaccharide, Interleukin-1, and Tumor Necrosis Factor. Role in the Acute Inflammatory Response. J. Biol. Chem. 1990, 265, 2856–2864. [Google Scholar]


| Experimental Diets 1 | |||
|---|---|---|---|
| Item | CON | SDP | PDP |
| Ingredient, % | |||
| Maize | 36.9 | 38.7 | 38.8 |
| Wheat | 16.0 | 16.0 | 16.0 |
| Extruded Soybeans | 15.0 | 11.4 | 11.2 |
| Barley | 13.0 | 13.0 | 13.0 |
| Soybean meal 44% crude protein (CP) | 6.8 | 6.8 | 6.8 |
| Soybean protein concentrate 56% CP | 5.6 | 5.6 | 5.6 |
| Sweet milk whey | 2.5 | 2.5 | 2.5 |
| Spray-dried plasma 80% CP | - | 2.0 | - |
| Porcine digestible peptides 62% CP | - | - | 2.0 |
| Mono calcium phosphate | 1.34 | 1.37 | 1.30 |
| Calcium carbonate | 0.62 | 0.64 | 0.67 |
| L-Lysine HCl | 0.55 | 0.49 | 0.52 |
| Salt | 0.53 | 0.40 | 0.16 |
| Vitamin-Mineral premix 2 | 0.40 | 0.40 | 0.40 |
| DL-Methionine | 0.27 | 0.23 | 0.57 |
| L-Threonine | 0.25 | 0.21 | 0.23 |
| L-Valine | 0.15 | 0.11 | 0.18 |
| L-Tryptophan | 0.09 | 0.08 | 0.10 |
| Calculated composition | |||
| Net Energy (NE), kcal/kg | 2470 | 2470 | 2470 |
| Dry Matter, % | 89.1 | 89.0 | 89.1 |
| Ash, % | 5.4 | 5.3 | 5.4 |
| Crude Protein, % | 19.5 | 19.7 | 19.5 |
| Calcium, % | 0.650 | 0.655 | 0.650 |
| Total P, % | 0.678 | 0.671 | 0.671 |
| Digestible amino acids | |||
| Lys, % | 1.280 | 1.280 | 1.280 |
| Met, % | 0.509 | 0.470 | 0.829 |
| Cys, % | 0.222 | 0.262 | 0.213 |
| Met+Cys, % | 0.768 | 0.768 | 1.078 |
| Thr, % | 0.832 | 0.832 | 0.832 |
| Trp, % | 0.282 | 0.282 | 0.282 |
| Val, % | 0.896 | 0.896 | 0.934 |
| Analyzed composition | |||
| Dry Matter, % | 89.1 | 88.8 | 88.8 |
| Ether Extract, % | 5.1 | 4.2 | 4.3 |
| Neutral Detergent Fiber, % | 10.5 | 10.2 | 10.0 |
| Crude Protein, % | 17.9 | 18.2 | 18.4 |
| Experimental Diets 1 | ||||
|---|---|---|---|---|
| Item | CON | SDP | SDP-PDP | Starter |
| Ingredients, % | ||||
| Maize | 29.68 | 32.53 | 32.11 | 29.79 |
| Soybean Meal 47% crude protein (CP) | 25.82 | 20.6 | 21.34 | 21.34 |
| Wheat | 16 | 16 | 16 | 15 |
| Barley | 6.5 | 6.5 | 6.5 | 20 |
| Dextrose | 6.5 | 6.5 | 6.5 | - |
| Sweet Milk Whey | 6.5 | 6.5 | 6.5 | - |
| Potato Protein | 2.5 | 2.5 | 2.5 | - |
| Lard | 2.67 | 2.38 | 2.31 | 6.53 |
| Di-calcium phosphate | 1.72 | 1.77 | 1.67 | 1.56 |
| Spray-dried plasma (80% CP) | - | 3 | 1 | - |
| Porcine digestible peptides 62% PB | - | - | 2 | - |
| Salt | 0.47 | 0.27 | 0.03 | 0.48 |
| Vitamin-Mineral Premix 2 | 0.40 | 0.40 | 0.40 | 0.40 |
| L-Lysine HCL (78) | 0.46 | 0.39 | 0.39 | 0.51 |
| DL-Methionine | 0.27 | 0.24 | 0.25 | 0.25 |
| L-Threonine | 0.21 | 0.16 | 0.17 | 0.24 |
| Calcium Carbonate | 0.11 | 0.11 | 0.17 | 0.55 |
| L-Valine | 0.11 | 0.09 | 0.08 | 0.14 |
| L-Tryptophan | 0.08 | 0.07 | 0.09 | 0.07 |
| Calculated composition | ||||
| Net Energy (NE), kcal/kg | 2470 | 2470 | 2470 | 2653 |
| Ether Extract, % | 4.69 | 4.43 | 4.39 | 8.61 |
| Neutral Detergent Fiber, % | 7.87 | 7.57 | 7.61 | 9.92 |
| Crude Protein, % | 20.51 | 20.50 | 20.50 | 18.50 |
| Calcium, % | 0.65 | 0.65 | 0.65 | 0.73 |
| Total P, % | 0.70 | 0.68 | 0.68 | 0.65 |
| Digestible amino acids | ||||
| Lys, % | 1.350 | 1.350 | 1.350 | 1.230 |
| Met, % | 0.545 | 0.504 | 0.539 | 0.478 |
| Met + Cys, % | 0.810 | 0.810 | 0.810 | 0.720 |
| Thr, % | 0.878 | 0.878 | 0.878 | 0.780 |
| Trp, % | 0.297 | 0.297 | 0.297 | 0.264 |
| Analyzed composition | ||||
| Dry Matter, % | 90.3 | 92.3 | 91.8 | 89.3 |
| Ether Extract, % | 4.6 | 4.3 | 4.1 | 8.4 |
| Neutral Detergent Fiber, % | 9.7 | 7.7 | 8.2 | - |
| Crude Protein, % | 19.7 | 18.7 | 19.6 | 17.7 |
| Experimental Diets 3 | |||||
|---|---|---|---|---|---|
| Item 2 | CON | SDP | PDP | SEM | p-Value 1 |
| BW, day 0, g | 7505 | 7517 | 7534 | 12.0 | 0.241 |
| day 14, g | 9438 b | 10,381 a | 9990 a | 138.9 | 0.001 |
| ADFI 0–14 d, g/d | 236 b | 295 a | 250 b | 11.3 | 0.002 |
| ADG 0–14 d, g/d | 138 b | 205 a | 175 a | 11.8 | 0.002 |
| G:F 0–14 d | 0.554 b | 0.695 a | 0.700 a | 0.0413 | 0.034 |
| Experimental Diets 3 | |||||
|---|---|---|---|---|---|
| Items 2 | CON | SDP | SDP-PDP | SEM | p-Value 1 |
| BW, day 0, g | 5724 | 5720 | 5721 | 46.9 | 0.859 |
| day 7, g | 5956 b | 6344 a | 6254 ab | 88.3 | 0.023 |
| day 14, g | 7165 b | 7894 a | 7871 a | 152.7 | 0.008 |
| day 35, g | 15,270 | 16,252 | 15,960 | 460.3 | 0.335 |
| ADFI, 0–14 d, g | 267 | 282 | 283 | 5.96 | 0.139 |
| 14–35 d, g | 483 | 508 | 525 | 25.9 | 0.532 |
| ADG, 0–14 d, g | 103 b | 155 a | 153 a | 10.7 | 0.007 |
| 14–35 d, g | 364 | 398 | 382 | 22.0 | 0.571 |
| G:F, 0–14 d | 0.380 b | 0.548 a | 0.540 a | 0.0383 | 0.046 |
| 14–35 d | 0.761 | 0.782 | 0.732 | 0.0501 | 0.840 |
| Function | Genes 1 | Experimental Diets 2 | Contrast Statistic | p-Value | q-Value (FDR) | ||
|---|---|---|---|---|---|---|---|
| CON | SDP | SDP-PDP | |||||
| Barrier function | CLDN15 | 0.51 b | 0.81 ab | 1.05 a | 4.984 | 0.027 | 0.139 |
| TFF3 | 1.15 b | 1.07 ab | 1.83 a | 4.290 | 0.035 | 0.145 | |
| Immune response | TLR4 | 1.54 b | 1.31 b | 2.86 a | 8.052 | 0.005 | 0.059 |
| GBP1 | 2.40 ab | 1.80 b | 4.20 a | 5.711 | 0.015 | 0.127 | |
| IFNGR1 | 0.69 b | 0.74 ab | 1.10 a | 4.661 | 0.028 | 0.139 | |
| Nutrient transport | SLC11A2/DMT1 | 0.37 b | 0.71 ab | 1.24 a | 3.862 | 0.046 | 0.169 |
| Enzyme/Hormone | HNMT | 0.86 b | 0.91 b | 1.56 a | 14.111 | 4 × 10−4 | 0.015 |
| APN | 1.59 b | 1.40 b | 3.40 a | 4.577 | 0.030 | 0.139 | |
| Oxidation | SOD2 | 1.04 b | 0.95 b | 1.71 a | 11.030 | 0.001 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Solé, F.; Criado-Mesas, L.; Villodre, C.; García, W.C.; Farré, M.; Borda, E.; Pérez-Cano, F.J.; Folch, J.M.; Solà-Oriol, D.; Pérez, J.F. Porcine Digestible Peptides (PDP) in Weanling Diets Regulates the Expression of Genes Involved in Gut Barrier Function, Immune Response and Nutrient Transport in Nursery Pigs. Animals 2020, 10, 2368. https://doi.org/10.3390/ani10122368
González-Solé F, Criado-Mesas L, Villodre C, García WC, Farré M, Borda E, Pérez-Cano FJ, Folch JM, Solà-Oriol D, Pérez JF. Porcine Digestible Peptides (PDP) in Weanling Diets Regulates the Expression of Genes Involved in Gut Barrier Function, Immune Response and Nutrient Transport in Nursery Pigs. Animals. 2020; 10(12):2368. https://doi.org/10.3390/ani10122368
Chicago/Turabian StyleGonzález-Solé, Francesc, Lourdes Criado-Mesas, Carmen Villodre, Wellington C. García, Mercè Farré, Elisabet Borda, Francisco J. Pérez-Cano, Josep M. Folch, David Solà-Oriol, and José F. Pérez. 2020. "Porcine Digestible Peptides (PDP) in Weanling Diets Regulates the Expression of Genes Involved in Gut Barrier Function, Immune Response and Nutrient Transport in Nursery Pigs" Animals 10, no. 12: 2368. https://doi.org/10.3390/ani10122368
APA StyleGonzález-Solé, F., Criado-Mesas, L., Villodre, C., García, W. C., Farré, M., Borda, E., Pérez-Cano, F. J., Folch, J. M., Solà-Oriol, D., & Pérez, J. F. (2020). Porcine Digestible Peptides (PDP) in Weanling Diets Regulates the Expression of Genes Involved in Gut Barrier Function, Immune Response and Nutrient Transport in Nursery Pigs. Animals, 10(12), 2368. https://doi.org/10.3390/ani10122368