Unexpected Gene-Flow in Urban Environments: The Example of the European Hedgehog

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
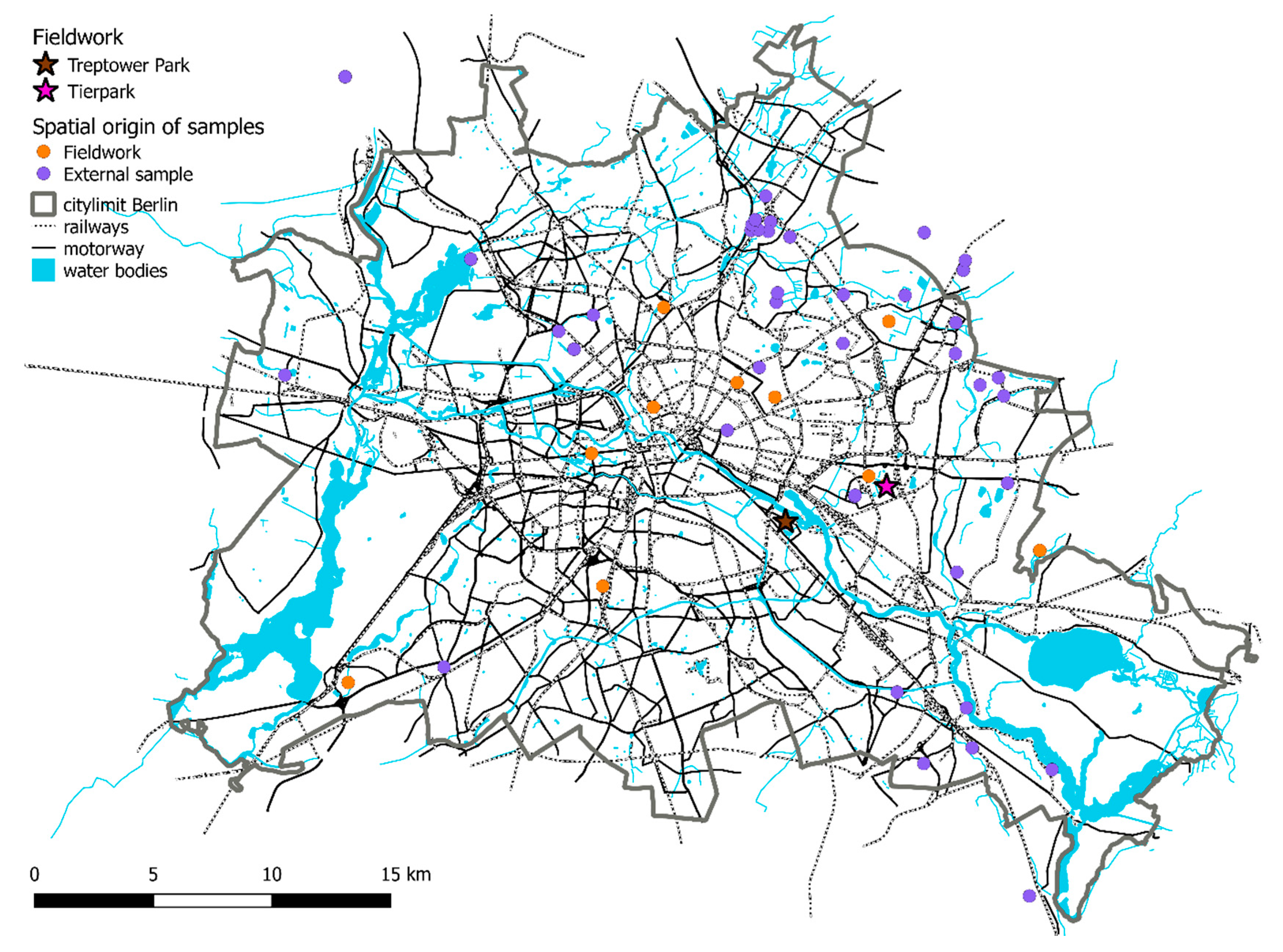
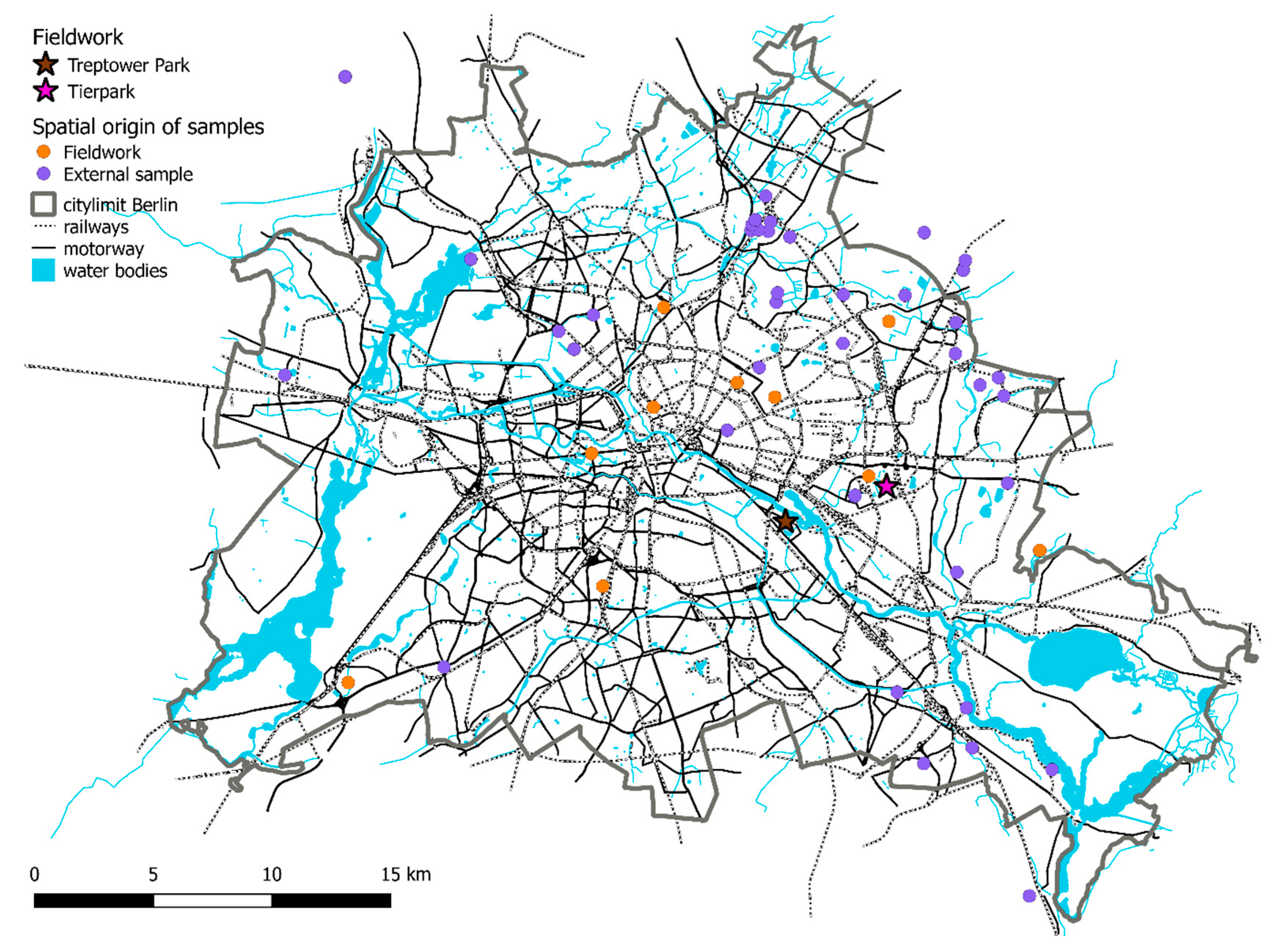
2.1. Sample Collection and Sites
2.2. DNA Extraction and Analysis of Microsatellite Loci
2.3. Data Analysis
2.4. Assignment
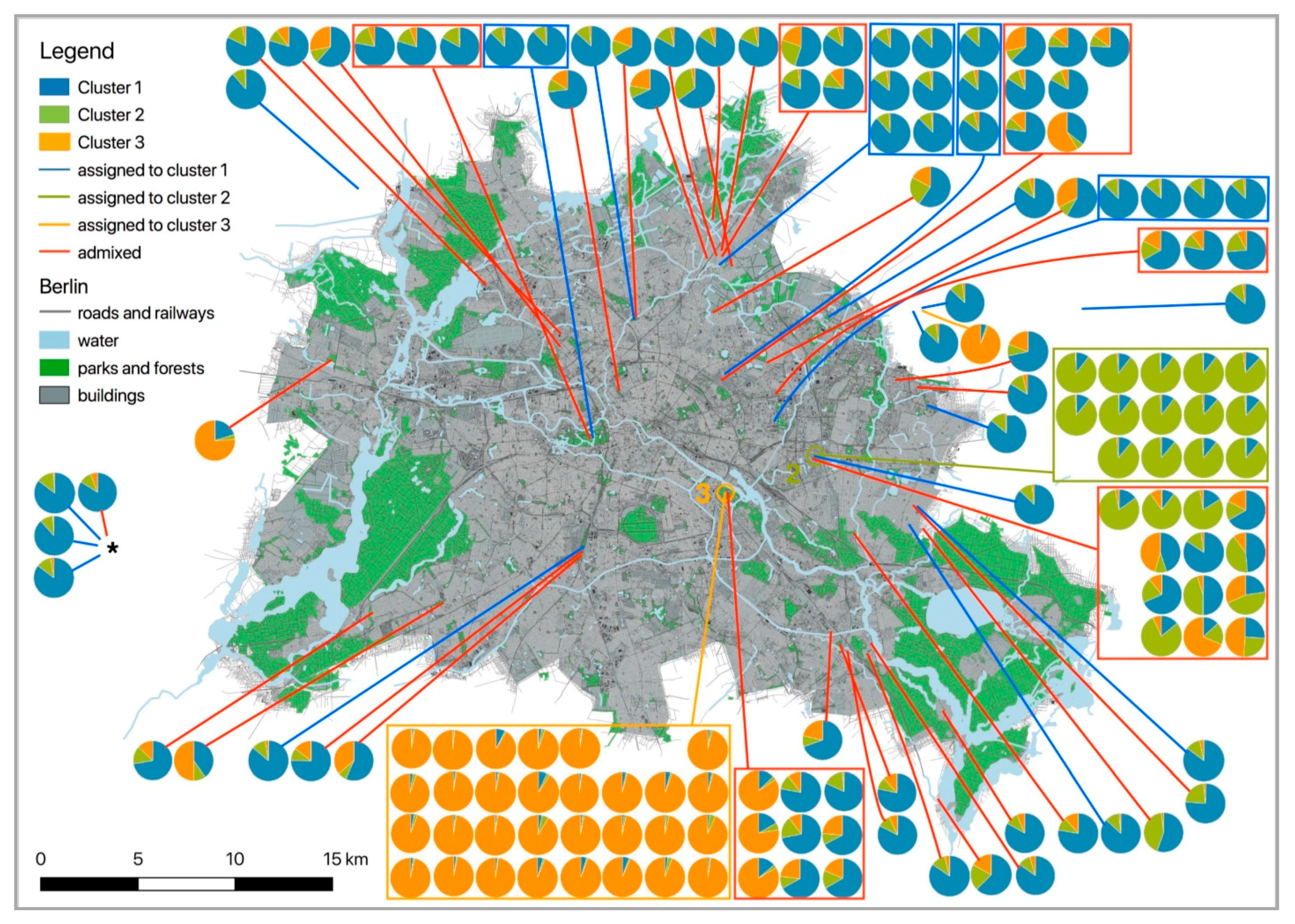
3. Results
3.1. Genotyping
3.1.1. All Individuals
3.1.2. Unrelated and Related Individuals
3.1.3. Migrants
3.2. Release of Hedgehogs after Rehabilitation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fischer, J.D.; Schneider, S.C.; Ahlers, A.A.; Miller, J.R. Categorizing wildlife responses to urbanization and conservation implications of terminology. Conserv. Biol. 2015, 29, 1246–1248. [Google Scholar] [CrossRef] [PubMed]
- Stillfried, M.; Fickel, J.; Börner, K.; Wittstatt, U.; Heddergott, M.; Ortmann, S.; Kramer-Schadt, S.; Frantz, A.C. Do cities represent sources, sinks or isolated islands for urban wild boar population structure? J. Appl. Ecol. 2017, 54, 272–281. [Google Scholar] [CrossRef]
- Pitra, C.; Lieckfeldt, D.; Frahnert, S.; Fickel, J. Phylogenetic relationships and ancestral areas of the bustards (Gruiformes: Otididae), inferred from mitochondrial DNA and nuclear intron sequences. Mol. Phylogenetics Evol. 2002, 23, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Schaub, T.; Meffert, P.J.; Kerth, G. Nest-boxes for Common Swifts Apus apus as compensatory measures in the context of building renovation: Efficacy and predictors of occupancy. Bird Conserv. Int. 2015, 26, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.T.J.; Munshi-South, J. Evolution of life in urban environments. Science 2017, 358, eaam8327. [Google Scholar] [CrossRef]
- Maclagan, S.J.; Coates, T.; Ritchie, E.G. Don’t judge habitat on its novelty: Assessing the value of novel habitats for an endangered mammal in a peri-urban landscape. Biol. Conserv. 2018, 223, 11–18. [Google Scholar] [CrossRef]
- Belasen, A.H.; Bletz, M.C.; da Silva Leite, D.; Toledo, L.F.; James, T.Y. Long-Term Habitat Fragmentation Is Associated With Reduced MHC IIB Diversity and Increased Infections in Amphibian Hosts. Front. Ecol. Evol. 2019, 6, 236. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, B.A.; Murphy, D.D. Conservation Strategy: The Effects of Fragmentation on Exinction. Am. Nat. 1985, 125, 879–887. [Google Scholar] [CrossRef]
- Munshi-South, J.; Nagy, C. Urban park characteristics, genetic variation, and historical demography of white-footed mouse (Peromyscus leucopus) populations in New York City. PeerJ 2014, 2, e310. [Google Scholar] [CrossRef] [Green Version]
- Lourenço, A.; Álvarez, D.; Wang, I.J.; Velo-Antón, G. Trapped within the city: Integrating demography, time since isolation and population-specific traits to assess the genetic effects of urbanization. Mol. Ecol. 2017, 26, 1498–1514. [Google Scholar] [CrossRef]
- Martins, R.F.; Fickel, J.; Le, M.; van Nguyen, T.; Nguyen, H.M.; Timmins, R.; Gan, H.M.; Rovie-Ryan, J.J.; Lenz, D.; Förster, D.W.; et al. Phylogeography of red muntjacs reveals three distinct mitochondrial lineages. BMC Evol. Biol. 2017, 17, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.; Weiss, G.H. The stepping stone model of population structure and the decrease of genetic correlation with distance. Genetics 1964, 49, 561–576. [Google Scholar] [PubMed]
- Opdam, P. Metapopulation theory and habitat fragmentation: A review of holarctic breeding bird studies. Landsc. Ecol. 1991, 5, 93–106. [Google Scholar] [CrossRef]
- Kimmig, S.E.; Beninde, J.; Brandt, M.; Schleimer, A.; Kramer-Schadt, S.; Hofer, H.; Börner, K.; Schulze, C.; Wittstatt, U.; Heddergott, M.; et al. Beyond the landscape: Resistance modelling infers physical and behavioural gene flow barriers to a mobile carnivore across a metropolitan area. Mol. Ecol. 2020, 29, 466–484. [Google Scholar] [CrossRef] [PubMed]
- Andrén, H. Effects of Habitat Fragmentation on Birds and Mammals in Landscapes with Different Proportions of Suitable Habitat: A Review. Oikos 1994, 71, 355. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.C.; Walker, F.M.; Goldingay, R.L.; Ball, T.; van der Ree, R. Degree of Landscape Fragmentation Influences Genetic Isolation among Populations of a Gliding Mammal. PLoS ONE 2011, 6, e26651. [Google Scholar] [CrossRef]
- Braaker, S.; Moretti, M.; Boesch, R.; Ghazoul, J.; Obrist, M.K.; Bontadina, F. Assessing habitat connectivity for ground-dwelling animals in an urban environment. Ecol. Appl. 2014, 24, 1583–1595. [Google Scholar] [CrossRef]
- Amori, G. Erinaceus europaeus. IUCN Red List Threat. Species 2016, e.T29650A2791303. [Google Scholar] [CrossRef]
- Doncaster, C.P.; Dickman, C.R. The ecology of small mammals in urban habitats. I. Population in a patchy environment. J. Anim. Ecol. 1987, 56, 629–640. [Google Scholar]
- Baker, P.J.; Harris, S. Urban mammals: What does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mamm. Rev. 2007, 37, 297–315. [Google Scholar] [CrossRef]
- Dowding, C.V.; Harris, S.; Poulton, S.; Baker, P.J. Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward. Anim. Behav. 2010, 80, 13–21. [Google Scholar] [CrossRef]
- Morris, P.A. A study of home range and movements in the hedgehog (Erinaceus europaeus). J. Zool. 1988, 214, 433–449. [Google Scholar] [CrossRef]
- Rautio, A.; Valtonen, A.; Kunnasranta, M. The effects of sex and season on home range in European hedgehogs at the northern edge of the species range. Ann. Zool. Fenn. 2013, 50, 107–123. [Google Scholar] [CrossRef]
- Zingg, R. Aktivität sowie Habitat- und Raumnutzung von Igeln (Erinaceus europaeus) in einem ländlichen Siedlungsgebiet. Ph.D. Thesis, Universität Zürich, Zurich, Swiss, 1994. (In German). [Google Scholar]
- Hubert, P.; Julliard, R.; Biagianti, S.; Poulle, M.-L.L.M. Ecological factors driving the higher hedgehog (Erinaceus europaeus) density in an urban area compared to the adjacent rural area. Landsc. Urban Plan. 2011, 103, 34–43. [Google Scholar] [CrossRef]
- Hof, A.R.; Snellenberg, J.; Bright, P.W. Food or fear? Predation risk mediates edge refuging in an insectivorous mammal. Anim. Behav. 2012, 83, 1099–1106. [Google Scholar] [CrossRef]
- Huijser, M.P.; Bergers, P.J.M. The effect of roads and traffic on hedgehog (Erinaceus europaeus) populations. Biol. Conserv. 2000, 95, 111–116. [Google Scholar] [CrossRef]
- Krange, M. Change in the occurrence of the West European Hedgehog (Erinaceus europaeus) in Western Sweden during 1950–2010; Karlstad University: Karlstad, Sweden, 2015. [Google Scholar]
- Hof, A.R.; Bright, P.W. Quantifying the long-term decline of the West European hedgehog in England by subsampling citizen-science datasets. Eur. J. Wildl. Res. 2016, 62, 407–413. [Google Scholar] [CrossRef]
- Taucher, A.L.; Gloor, S.; Dietrich, A.; Geiger, M.; Hegglin, D.; Bontadina, F. Decline in distribution and abundance: Urban hedgehogs under pressure. Animals 2020, 10, 1606. [Google Scholar] [CrossRef]
- Müller, F. Langzeit-Monitoring der Straßenverkehrsopfer beim Igel (Erinaceus europaeus L.) zur Indikation von Populationsdichteveränderungen entlang zweier Teststrecken im Landkreis Fulda. Beiträge zur Naturkunde Osthessen 2018, 54, 21–26. (In German) [Google Scholar]
- Hof, A.R.A.; Bright, P.P.W. The value of green-spaces in built-up areas for western hedgehogs. Lutra 2009, 52, 69–82. [Google Scholar]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intra-urban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef]
- Bolfíková, B.Č.; Hulva, P. Microevolution of sympatry: Landscape genetics of hedgehogs Erinaceus europaeus and E. roumanicus in Central Europe. Heredity (Edinb.) 2012, 108, 248–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becher, S.A.; Griffiths, R. Isolation and characterization of six polymorphic microsatellite loci in the European hedgehog Erinaceus europaeus. Mol. Ecol. 1997, 6, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Henderson, M.; Becher, S.A.; Doncaster, C.P.; Maclean, N. Five new polymorphic microsatellite loci in the European hedgehog Erinaceus europaeus. Mol. Ecol. 2000, 9, 1949–1951. [Google Scholar] [CrossRef]
- Miller, C.R.; Joyce, P.; Waits, L.P. Assessing allelic dropout and genotype reliability using maximum likelihood. Genetics 2002, 160, 357–366. [Google Scholar]
- Guo, S.W.; Thompson, E.A. Performing the Exact Test of Hardy-Weinberg Proportion for Multiple Alleles. Biometrics 1992, 48, 361. [Google Scholar] [CrossRef] [PubMed]
- Marshall, T.C.; Slate, J.; Kruuk, L.E.B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Rodríguez-Ramilo, S.T.; Wang, J. The effect of close relatives on unsupervised Bayesian clustering algorithms in population genetic structure analysis. Mol. Ecol. Resour. 2012, 12, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Queller, D.C.; Goodnight, K.F. Estimating Relatedness Using Genetic Markers. Evolution 1989, 43, 258–275. [Google Scholar] [CrossRef]
- Wang, J. Coancestry: A program for simulating, estimating and analysing relatedness and inbreeding coefficients. Mol. Ecol. Resour. 2011, 11, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [PubMed]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Corander, J.; Sirén, J.; Arjas, E. Bayesian Spatial Modelling of Genetic Population Structure. Comput. Stat. 2008, 23, 111–129. [Google Scholar] [CrossRef]
- Cheng, L.; Connor, T.R.; Sirén, J.; Aanensen, D.M.; Corander, J. Hierarchical and spatially explicit clustering of DNA sequences with Baps software. Mol. Biol. Evol. 2013, 30, 1224–1228. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Moßbrucker, A.M.; Apriyana, I.; Fickel, J.; Imron, M.A.; Pudyatmoki, S.; Sumardi; Suryadi, H. Non-invasive genotyping of Sumatran elephants: Implications for conservation. Trop. Conserv. Sci. 2015, 8, 745–759. [Google Scholar]
- Schmidt, K.; Kowalczyk, R.; Ozolins, J.; Männil, P.; Fickel, J. Genetic structure of the Eurasian lynx population in north-eastern Poland and the Baltic states. Cons. Genet. 2009, 10, 497–501. [Google Scholar] [CrossRef]
- Coombs, J.A.; Letcher, B.H.; Nislow, K.H. Create: A software to create input files from diploid genotypic data for 52 genetic software programs. Mol. Ecol. Resour. 2008, 8, 578–580. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 1 April 2017).
- Becher, S.A.; Griffiths, R. Genetic differentiation among local populations of the European hedgehog (Erinaceus europaeus) in mosaic habitat. Mol. Ecol. 1998, 7, 1599–1604. [Google Scholar] [CrossRef] [PubMed]
- Wahlund, S. Zusammensetzung von Populationen und Korrelationserscheinungen vom Standpunkt der Vererbungslehre aus betrachtet. Hereditas 1928, 11, 65–106. (In German) [Google Scholar] [CrossRef]
- Braaker, S.; Kormann, U.; Bontadina, F.; Obrist, M.K.K. Prediction of genetic connectivity in urban ecosystems by combining detailed movement data, genetic data and multi-path modelling. Landsc. Urban Plan 2017, 160, 107–114. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Nielsen, J.L.; Jones, O.R.; Berg, T.B.; Pertoldi, G. Genetic structure of the European hedgehog (Erinaceus europaeus) in Denmark. PLoS ONE 2020, 15, e0227205. [Google Scholar] [CrossRef]
- Bolfíková, B.; Konečný, A.; Pfäffle, M.; Skuballa, J.; Hulva, P. Population biology of establishment in New Zealand hedgehogs inferred from genetic and historical data: Conflict or compromise? Mol. Ecol. 2013, 22, 3709–3720. [Google Scholar] [CrossRef]
- Morris, P.A.; Reeve, N.J. Hedgehog Erinaceus europaeus. In Mammals of the British Isles: Handbook, 4th ed.; Harris, S., Yalden, D.W., Eds.; The Mammal Society: Southampton, UK, 2008; pp. 241–248. [Google Scholar]
- Rasmussen, S.L.; Berg, T.B.; Dabelsteen, T.; Jones, O.R. The ecology of suburban juvenile European hedgehogs (Erinaceus erinaceus) in Denmark. Ecol. Evol. 2019, 9, 13174–13187. [Google Scholar] [CrossRef] [Green Version]
- Doncaster, C.P.; Rondinini, C.; Johnson, P.C.D. Field test for environmental correlates of dispersal in hedgehogs Erinaceus europaeus. J. Anim. Ecol. 2001, 70, 33–46. [Google Scholar]
- Rondinini, C.; Doncaster, C.P. Roads as barriers to movement for hedgehogs. Funct. Ecol. 2002, 16, 504–509. [Google Scholar] [CrossRef]
- Herter, K. Gefangenschaftsbeobachtungen an europäischen Igeln II. Mamm. Biol. (formerly Zeitschrift für Säugetierkunde) 1933, 8, 195–218. [Google Scholar]
- SenUVK: Senatsverwaltung für Umwelt, Verkehr und Klimaschutz Berlin, Referat Freiraumplanung und Stadtgrün, Grünflacheninformationssystem GRIS. 2018. Available online: https://www.berlin.de/senuvk/umwelt/stadtgruen/gruenanlagen/de/daten_fakten/downloads/ausw_5.pdf (accessed on 31 January 2019).
- Ploi, K.; Curto, M.; Bolfíková, B.Č.; Loudová, M.; Hulva, P.; Seiter, A.; Fuhrmann, M.; Winter, S.; Meimberg, H. Evaluating the impact of wildlife shelter management on the genetic diversity of Erinaceus europaeus and E. roumanicus in their contact zone. Animals 2020, 10, 1452. [Google Scholar] [CrossRef] [PubMed]
- Molony, S.E.; Dowding, C.V.; Baker, P.J.; Cuthill, I.C.; Harris, S. The effect of translocation and temporary captivity on wildlife rehabilitation success: An experimental study using European hedgehogs (Erinaceus europaeus). Biol. Conserv. 2006, 130, 530–537. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Locus | Ntyped | NA | Allele Size Range (bp) | HO | HE | HWE | fNull |
|---|---|---|---|---|---|---|---|
| EEU1 | 143 | 8 | 129–143 | 0.671 | 0.773 | + | 0.062 |
| EEU2 | 141 | 13 | 257–281 | 0.752 | 0.863 | + | 0.064 |
| EEU3 | 142 | 15 | 131–181 | 0.754 | 0.868 | + | 0.064 |
| EEU4 | 143 | 14 | 144–170 | 0.699 | 0.785 | + | 0.052 |
| EEU5 | 143 | 13 | 107–139 | 0.678 | 0.711 | + | 0.011 |
| EEU6 | 143 | 6 | 145–159 | 0.350 | 0.331 | + | −0.049 |
| EEU12H | 143 | 4 | 91–97 | 0.497 | 0.615 | + | 0.098 |
| EEU37H | 142 | 16 | 236–280 | 0.676 | 0.839 | - | 0.095 |
| EEU43H | 143 | 12 | 146–172 | 0.657 | 0.730 | + | 0.047 |
| EEU54H | 142 | 8 | 276–296 | 0.479 | 0.551 | + | 0.067 |
| Mean SD | 142.5 0.71 | 10.9 4.09 | 0.621 0.133 | 0.707 0.167 | |||
| Cluster assignment (Q > 0.85) | mean NA | HO | HE | HWE | |||
| 1 | 29 | 8.6 | 0.623 | 0.685 | + | ||
| 2 (“Tierpark”) | 14 | 3.3 | 0.557 | 0.524 | + | ||
| 3 (“Trepower Park”) | 31 | 6.0 | 0.578 | 0.651 | + | ||
| single cluster (N = 65) | 65 | 10.6 | 0.672 | 0.731 | + | ||
| no cluster, all (N = 143) | 143 | 10.9 | 0.621 | 0.706 | - * |
| Internal ID | Locality | STRUCTURE Q-Value for Cluster 1 | STRUCTURE Q-Value for Cluster 2 | STRUCTURE Q-Value for Cluster 3 | STRUCTURE Cluster Assignment (Q ≥ 0.85) | BAPS Cluster Assignment * |
|---|---|---|---|---|---|---|
| 147 | Tiergarten, Berlin | 0.87559 | 0.10548 | 0.01895 | 1 | 4 |
| 167 | Tiergarten, Berlin | 0.88068 | 0.10901 | 0.01031 | 1 | 4 |
| 176 | Eisenhuettenstadt | 0.85406 | 0.11966 | 0.02629 | 1 | 1 |
| 161 | Hans-Baluschek-Park, | 0.86127 | 0.11218 | 0.02655 | 1 | 1 |
| 311 | Tierpark, Berlin | 0.87213 | 0.10962 | 0.01824 | 1 | 1 |
| 334 | 12623 Berlin | 0.86935 | 0.11628 | 0.01438 | 1 | 1 |
| 335 | 12623 Berlin | 0.85235 | 0.11496 | 0.03271 | 1 | 1 |
| 220 | Friedenstr., Berlin Near Volkspark Friedrichshain | 0.87103 | 0.11977 | 0.00921 | 1 | 1 |
| 175 | Volkspark Prenzlauerberg, Berlin | 0.86135 | 0.105 | 0.03368 | 1 | 1 |
| 199 | Hellersdorf, Berlin | 0.86134 | 0.12691 | 0.01175 | 1 | 1 |
| 159 | Prenzlauerberg | 0.85732 | 0.10238 | 0.04031 | 1 | 1 |
| 338 | Park am Weidengrund | 0.87595 | 0.11224 | 0.01182 | 1 | 1 |
| 117 | Near graveyard | 0.88408 | 0.10591 | 0.01 | 1 | 1 |
| 326 | Zum Erlenbruch, 15344 Strausberg | 0.86423 | 0.11255 | 0.02321 | 1 | 1 |
| 156 | Buergerpark Pankow-Berlin | 0.86845 | 0.11301 | 0.01854 | 1 | 1 |
| 341 | Kleingartenanlage 750 Jahre Berlin, 13057 Berlin | 0.85329 | 0.10368 | 0.04302 | 1 | 1 |
| 328 | Warnemuender Str. 18, 13059 Berlin | 0.86432 | 0.11747 | 0.01821 | 1 | 1 |
| 329 | Warnemuender Str. 18, 13059 Berlin | 0.86724 | 0.1151 | 0.01767 | 1 | 1 |
| 330 | Warnemuender Str. 18, 13059 Berlin | 0.87485 | 0.1077 | 0.01746 | 1 | 1 |
| 337 | KGA Maerchenland, 13089 Berlin | 0.87728 | 0.10819 | 0.01456 | 1 | 1 |
| 231 | Friedenstr. 8, 16356 Ahrensfelde | 0.87273 | 0.10608 | 0.02116 | 1 | 1 |
| 257 | Dietrichstr. 5, 16356 Ahrensfelde | 0.87167 | 0.10645 | 0.0219 | 1 | 1 |
| 193 | Jungbornstr., 13129 Berlin | 0.85709 | 0.12218 | 0.02072 | 1 | 1 |
| 189 | Strasse 7, 13129 Berlin | 0.85836 | 0.11349 | 0.02814 | 1 | 1 |
| 194 | Schwarzwaldstr./Ilsenstr., 13129 Berlin | 0.85756 | 0.11789 | 0.02457 | 1 | 1 |
| 185 | Gutenfelsstr. 14, 13129 Berlin | 0.8736 | 0.11369 | 0.01271 | 1 | 1 |
| 187 | Gutenfelsstr. 14, 13129 Berlin | 0.8837 | 0.1066 | 0.0097 | 1 | 1 |
| 113 | Choise-le-Roi-Str. 3, Berlin | 0.88496 | 0.10623 | 0.00881 | 1 | 1 |
| 118 | Vielitzsee Ortsteil Strubensee, 16835 green area | 0.85436 | 0.13413 | 0.01153 | 1 | 1 |
| 179 | Eisenhuettenstadt | 0.84074 | 0.10226 | 0.05701 | admixed | 1 |
| 243 | Zeuthen | 0.6249 | 0.20766 | 0.16744 | admixed | 1 |
| 129 | Rohrwallallee 10. 12527 Berlin | 0.83142 | 0.10412 | 0.06446 | admixed | 1 |
| 120 | Altglienike Feldweg | 0.84766 | 0.10475 | 0.0476 | admixed | 1 |
| 137 | Kablower Weg 89, 12526 Berlin | 0.78369 | 0.0984 | 0.1179 | admixed | 1 |
| 138 | Kablower Weg 89, 12526 Berlin | 0.8267 | 0.1031 | 0.0702 | admixed | 1 |
| 125 | Riesserseestr. 10. 12527 Berlin | 0.84665 | 0.11413 | 0.03923 | admixed | 1 |
| 135 | Korkedamm 73, 12524 Berlin | 0.69588 | 0.08972 | 0.21436 | admixed | 1 |
| 127 | Rehwiese, Gerkrathstraße 2 Park | 0.72131 | 0.14963 | 0.12906 | admixed | 1 |
| 182 | Zehlendorf, Berlin | 0.40151 | 0.09898 | 0.49948 | admixed | 1 |
| 158 | Hans-Baluschek-Park, 10829 Berlin | 0.56664 | 0.07147 | 0.36189 | admixed | 1 |
| 169 | Hans-Baluschek-Park, 10829 Berlin | 0.75432 | 0.10163 | 0.14405 | admixed | 1 |
| 235 | Glasberger Str. 43, 12555 Berlin | 0.77728 | 0.10906 | 0.11368 | admixed | 1 |
| 110 | Trainierbahn Hoppegarten | 0.55419 | 0.42731 | 0.01849 | admixed | 1 |
| A35_088 | Treptower Park | 0.66656 | 0.14738 | 0.18608 | admixed | 1 |
| A4_317 | Treptower Park | 0.81876 | 0.16614 | 0.01512 | admixed | 1 |
| A61_108 | Treptower Park | 0.67049 | 0.09899 | 0.23051 | admixed | 1 |
| A68_108 | Treptower Park | 0.71856 | 0.17835 | 0.10308 | admixed | 1 |
| 126 | Moldaustr. 30. 10319 Berlin near Tierpark | 0.68485 | 0.2057 | 0.10944 | admixed | 1 |
| 128 | Moldaustr. 24, 10319 Berlin near Tierpark | 0.13048 | 0.18259 | 0.68692 | admixed | 1 |
| 136 | Moldaustr. 24, 10319 Berlin near Tierpark | 0.2581 | 0.25296 | 0.48897 | admixed | 1 |
| 174 | Tierpark, Berlin | 0.48697 | 0.40986 | 0.10316 | admixed | 1 |
| 308 | Tierpark, Berlin | 0.8438 | 0.13492 | 0.02127 | admixed | 1 |
| 310 | Tierpark, Berlin | 0.45013 | 0.09578 | 0.45409 | admixed | 1 |
| 314 | Tierpark, Berlin | 0.50102 | 0.45028 | 0.0487 | admixed | 1 |
| 317 | Tierpark, Berlin | 0.66602 | 0.15881 | 0.17519 | admixed | 1 |
| 320 | Tierpark, Berlin | 0.22576 | 0.45684 | 0.3174 | admixed | 1 |
| 333 | 12623 Berlin | 0.76214 | 0.22126 | 0.01663 | admixed | 1 |
| 143 | Tiergarten, Berlin | 0.82986 | 0.15939 | 0.01076 | admixed | 1 |
| 152 | Tiergarten, Berlin | 0.79801 | 0.14577 | 0.05625 | admixed | 1 |
| 166 | Tiergarten, Berlin | 0.78003 | 0.16683 | 0.05316 | admixed | 1 |
| 309 | Nordbahnhof park | 0.72988 | 0.1093 | 0.16079 | admixed | 1 |
| 134 | Volkspark Prenzlauerberg, Berlin | 0.84225 | 0.1053 | 0.05244 | admixed | 1 |
| 142 | Volkspark Prenzlauerberg, Berlin | 0.62693 | 0.08785 | 0.28521 | admixed | 1 |
| 153 | Volkspark Prenzlauerberg, Berlin | 0.77372 | 0.0954 | 0.13087 | admixed | 1 |
| 168 | Volkspark Prenzlauerberg, Berlin | 0.75805 | 0.0945 | 0.14747 | admixed | 1 |
| 170 | Volkspark Prenzlauerberg, Berlin | 0.35675 | 0.0564 | 0.58687 | admixed | 1 |
| 172 | Volkspark Prenzlauerberg, Berlin | 0.83185 | 0.10633 | 0.06181 | admixed | 1 |
| 324 | Eisenacher Str.,12629 Berlin near park | 0.71554 | 0.09224 | 0.19223 | admixed | 1 |
| 300 | Kastanienallee 122/126, 12627 Berlin near Teupitzer Park | 0.83854 | 0.12196 | 0.03952 | admixed | 1 |
| 261 | Wolfshorststr. 25, 13591 Berlin | 0.20168 | 0.04025 | 0.75806 | admixed | 1 |
| 340 | Mahlerstraße, 13088 Berlin | 0.75765 | 0.09987 | 0.14248 | admixed | 1 |
| 241 | Glambecker Ring 4, 12679 Berlin | 0.66759 | 0.16157 | 0.17085 | admixed | 1 |
| 114 | Togostr. 45, 13351 Berlin near Volkspark Rehberge | 0.61322 | 0.10559 | 0.28119 | admixed | 1 |
| 248 | 13053 Berlin | 0.58362 | 0.0891 | 0.32725 | admixed | 1 |
| 119 | Ghanastr. 27, 13351 Berlin near Volkspark Rehberge | 0.7999 | 0.09627 | 0.10382 | admixed | 1 |
| 139 | Falkenberger Krugwiesen, 13057 Berlin | 0.78143 | 0.11325 | 0.10533 | admixed | 1 |
| 140 | Falkenberger Krugwiesen, 13057 Berlin | 0.73941 | 0.18507 | 0.0755 | admixed | 1 |
| 150 | Buergerpark Pankow-Berlin | 0.66583 | 0.13902 | 0.19514 | admixed | 1 |
| 188 | Schwarzelfenweg 19, 13088 Berlin | 0.59525 | 0.23319 | 0.17157 | admixed | 1 |
| 116 | Alt-Tegel 47c, 13507 Berlin | 0.82319 | 0.14861 | 0.0282 | admixed | 1 |
| 191 | Strasse 26 Nr. 30. 13129 Berlin near green area | 0.65149 | 0.31845 | 0.03009 | admixed | 1 |
| 196 | Schwarzwaldstr., 13129 Berlin | 0.67859 | 0.10107 | 0.22037 | admixed | 1 |
| 186 | Gutenfelsstr. 14, 13129 Berlin | 0.81717 | 0.1626 | 0.02022 | admixed | 1 |
| 198 | Gutenfelsstr. 14, 13129 Berlin | 0.76845 | 0.1193 | 0.11227 | admixed | 1 |
| 200 | Gutenfelsstr. 14, 13129 Berlin | 0.8493 | 0.10802 | 0.04267 | admixed | 1 |
| 192 | Urbacher Str., 13129 Berlin | 0.54983 | 0.25383 | 0.19634 | admixed | 1 |
| 184 | Freischuetzstr., 13129 Berlin | 0.81933 | 0.14711 | 0.03358 | admixed | 1 |
| 203 | Freischuetzstr., 13129 Berlin | 0.8327 | 0.13544 | 0.03187 | admixed | 1 |
| 197 | Krontalerstr., 13125 Berlin | 0.84704 | 0.10899 | 0.04399 | admixed | 1 |
| A3_317 | Treptower Park | 0.77842 | 0.11654 | 0.10505 | admixed | admixed |
| A34_078 | Treptower Park | 0.67359 | 0.09012 | 0.23627 | admixed | admixed |
| 144 | Tierpark, Berlin | 0.10456 | 0.88882 | 0.00662 | 2 | 2 |
| 146 | Tierpark, Berlin | 0.10652 | 0.8851 | 0.00836 | 2 | 2 |
| 154 | Tierpark, Berlin | 0.10959 | 0.88179 | 0.00862 | 2 | 2 |
| 165 | Tierpark, Berlin | 0.10523 | 0.88487 | 0.00991 | 2 | 2 |
| 305 | Tierpark, Berlin | 0.10535 | 0.88925 | 0.00539 | 2 | 2 |
| 306 | Tierpark, Berlin | 0.10434 | 0.89067 | 0.005 | 2 | 2 |
| 307 | Tierpark, Berlin | 0.12173 | 0.85912 | 0.01916 | 2 | 2 |
| 312 | Tierpark, Berlin | 0.10548 | 0.88768 | 0.00684 | 2 | 2 |
| 313 | Tierpark, Berlin | 0.10518 | 0.88331 | 0.0115 | 2 | 2 |
| 315 | Tierpark, Berlin | 0.11634 | 0.86767 | 0.01601 | 2 | 2 |
| 318 | Tierpark, Berlin | 0.11617 | 0.87305 | 0.01078 | 2 | 2 |
| 319 | Tierpark, Berlin | 0.11028 | 0.87549 | 0.01424 | 2 | 2 |
| 342 | Tierpark, Berlin | 0.11259 | 0.87716 | 0.01028 | 2 | 2 |
| 344 | IZW Garten, Berlin (bordering with Tierpark) | 0.10948 | 0.87852 | 0.01199 | 2 | 2 |
| 141 | Tierpark, Berlin | 0.14033 | 0.79122 | 0.06844 | admixed | 2 |
| 149 | Tierpark, Berlin | 0.10468 | 0.79688 | 0.09844 | admixed | 2 |
| 321 | Tierpark, Berlin | 0.15174 | 0.82023 | 0.02802 | admixed | 2 |
| 322 | Tierpark, Berlin | 0.1618 | 0.8123 | 0.02591 | admixed | 2 |
| 157 | Treptower Park | 0.02233 | 0.01133 | 0.96636 | 3 | 3 |
| 345 | Treptower Park | 0.01293 | 0.01151 | 0.97556 | 3 | 3 |
| 346 | Treptower Park | 0.02053 | 0.00877 | 0.97068 | 3 | 3 |
| 348 | Treptower Park | 0.07228 | 0.0132 | 0.9145 | 3 | 3 |
| 349 | Treptower Park | 0.0197 | 0.00997 | 0.97034 | 3 | 3 |
| 350 | Treptower Park | 0.01276 | 0.01003 | 0.97721 | 3 | 3 |
| A1_317 | Treptower Park | 0.01622 | 0.00786 | 0.97591 | 3 | 3 |
| A10_028 | Treptower Park | 0.02804 | 0.02522 | 0.94674 | 3 | 3 |
| A11_028 | Treptower Park | 0.06713 | 0.02515 | 0.90771 | 3 | 3 |
| A12_028 | Treptower Park | 0.03142 | 0.0546 | 0.914 | 3 | 3 |
| A13_028 | Treptower Park | 0.04447 | 0.01617 | 0.93935 | 3 | 3 |
| A14_028 | Treptower Park | 0.01659 | 0.00572 | 0.97765 | 3 | 3 |
| A15_028 | Treptower Park | 0.01453 | 0.01432 | 0.97112 | 3 | 3 |
| A16_028 | Treptower Park | 0.04624 | 0.01015 | 0.94359 | 3 | 3 |
| A2_317 | Treptower Park | 0.03028 | 0.01088 | 0.95885 | 3 | 3 |
| A20_038 | Treptower Park | 0.01454 | 0.01811 | 0.96735 | 3 | 3 |
| A21_038 | Treptower Park | 0.05401 | 0.01349 | 0.9325 | 3 | 3 |
| A22_038 | Treptower Park | 0.04167 | 0.01371 | 0.94463 | 3 | 3 |
| A25_078 | Treptower Park | 0.01262 | 0.02414 | 0.96325 | 3 | 3 |
| A27_078 | Treptower Park | 0.02487 | 0.03366 | 0.94146 | 3 | 3 |
| A28_078 | Treptower Park | 0.01631 | 0.01948 | 0.96424 | 3 | 3 |
| A30_078 | Treptower Park | 0.02939 | 0.01268 | 0.95794 | 3 | 3 |
| A31_078 | Treptower Park | 0.01652 | 0.04733 | 0.93614 | 3 | 3 |
| A32_078 | Treptower Park | 0.01376 | 0.01006 | 0.97617 | 3 | 3 |
| A37_088 | Treptower Park | 0.01249 | 0.01269 | 0.97483 | 3 | 3 |
| A43_088 | Treptower Park | 0.02567 | 0.02771 | 0.94662 | 3 | 3 |
| A47_098 | Treptower Park | 0.01487 | 0.01031 | 0.97481 | 3 | 3 |
| A5_317 | Treptower Park | 0.02926 | 0.0273 | 0.94345 | 3 | 3 |
| A59_108 | Treptower Park | 0.0119 | 0.00944 | 0.97865 | 3 | 3 |
| A9_028 | Treptower Park | 0.00902 | 0.00582 | 0.98515 | 3 | 3 |
| 252 | Friedenstr., 16356 Ahrensfelde | 0.04766 | 0.01531 | 0.93704 | 3 | 3 |
| A56_098 | Treptower Park | 0.12642 | 0.03009 | 0.84349 | admixed | 3 |
| A62_108 | Treptower Park | 0.13874 | 0.02767 | 0.83359 | admixed | 3 |
| 343 | Tierpark, Berlin | 0.16607 | 0.05859 | 0.77535 | admixed | admixed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barthel, L.M.F.; Wehner, D.; Schmidt, A.; Berger, A.; Hofer, H.; Fickel, J. Unexpected Gene-Flow in Urban Environments: The Example of the European Hedgehog. Animals 2020, 10, 2315. https://doi.org/10.3390/ani10122315
Barthel LMF, Wehner D, Schmidt A, Berger A, Hofer H, Fickel J. Unexpected Gene-Flow in Urban Environments: The Example of the European Hedgehog. Animals. 2020; 10(12):2315. https://doi.org/10.3390/ani10122315
Chicago/Turabian StyleBarthel, Leon M. F., Dana Wehner, Anke Schmidt, Anne Berger, Heribert Hofer, and Jörns Fickel. 2020. "Unexpected Gene-Flow in Urban Environments: The Example of the European Hedgehog" Animals 10, no. 12: 2315. https://doi.org/10.3390/ani10122315