First Insights into the Population Genetic Structure and Heterozygosity–Fitness Relationship in Roe Deer Inhabiting the Area between the Alps and Dinaric Mountains



Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethical Statement
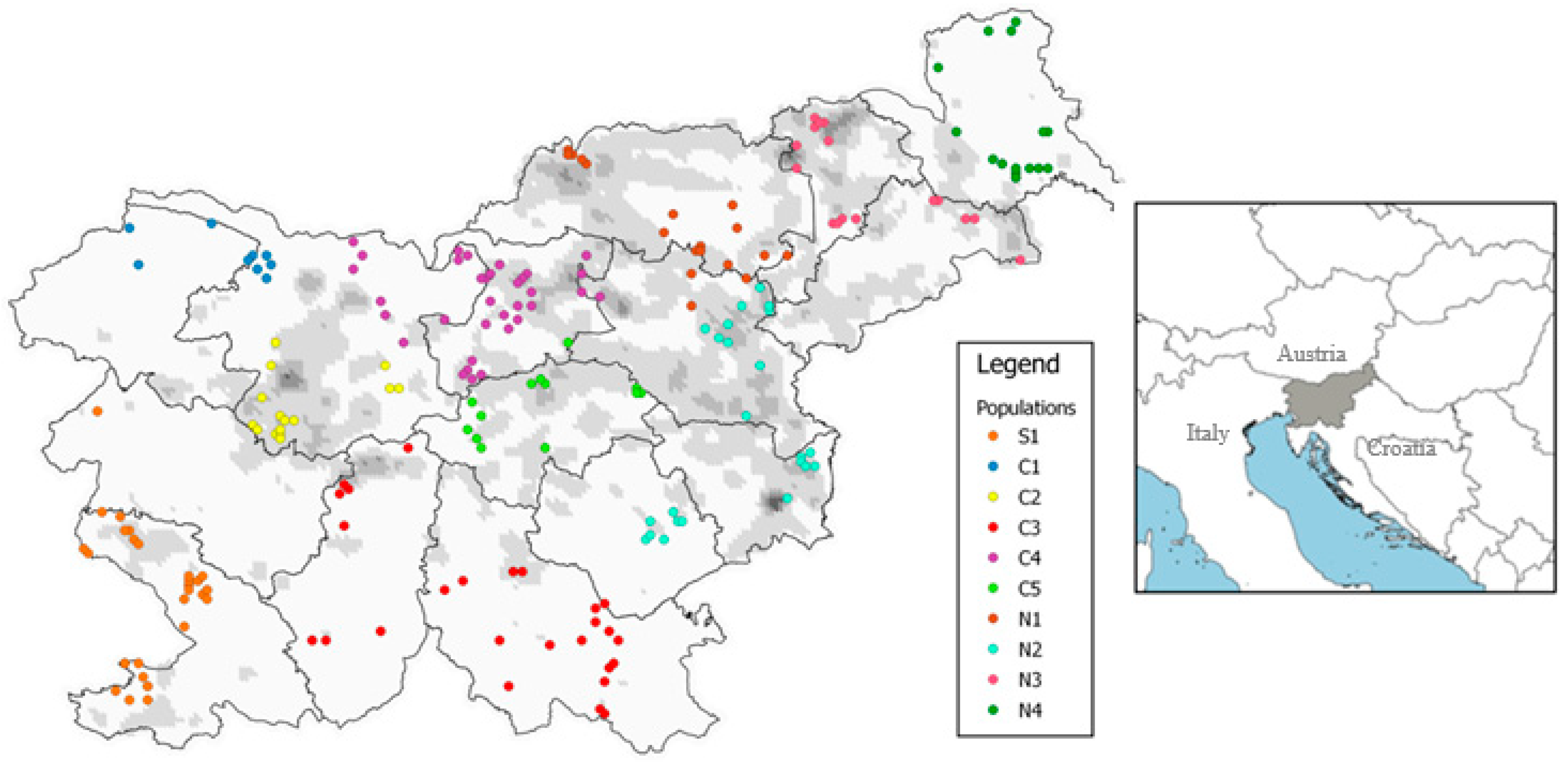
2.2. Study Area and Sampling
2.3. Data Collection and Preparation
2.4. DNA Extraction and Microsatellite Genotyping
2.5. Determination of Genetic Variability among Populations
2.6. Effects of Genetic Features (Heterozygosity) on Body Mass and Reproductive Ability
3. Result
3.1. Intra-Population Genetic Diversity
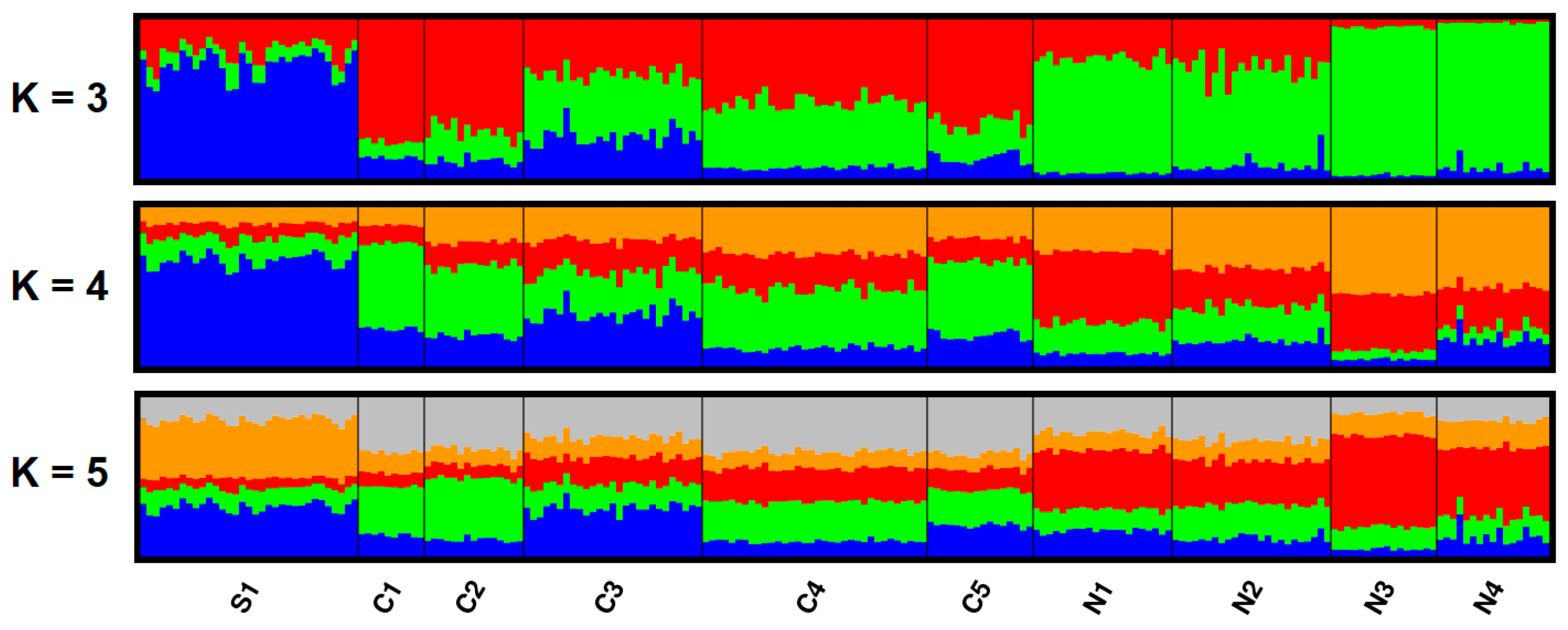
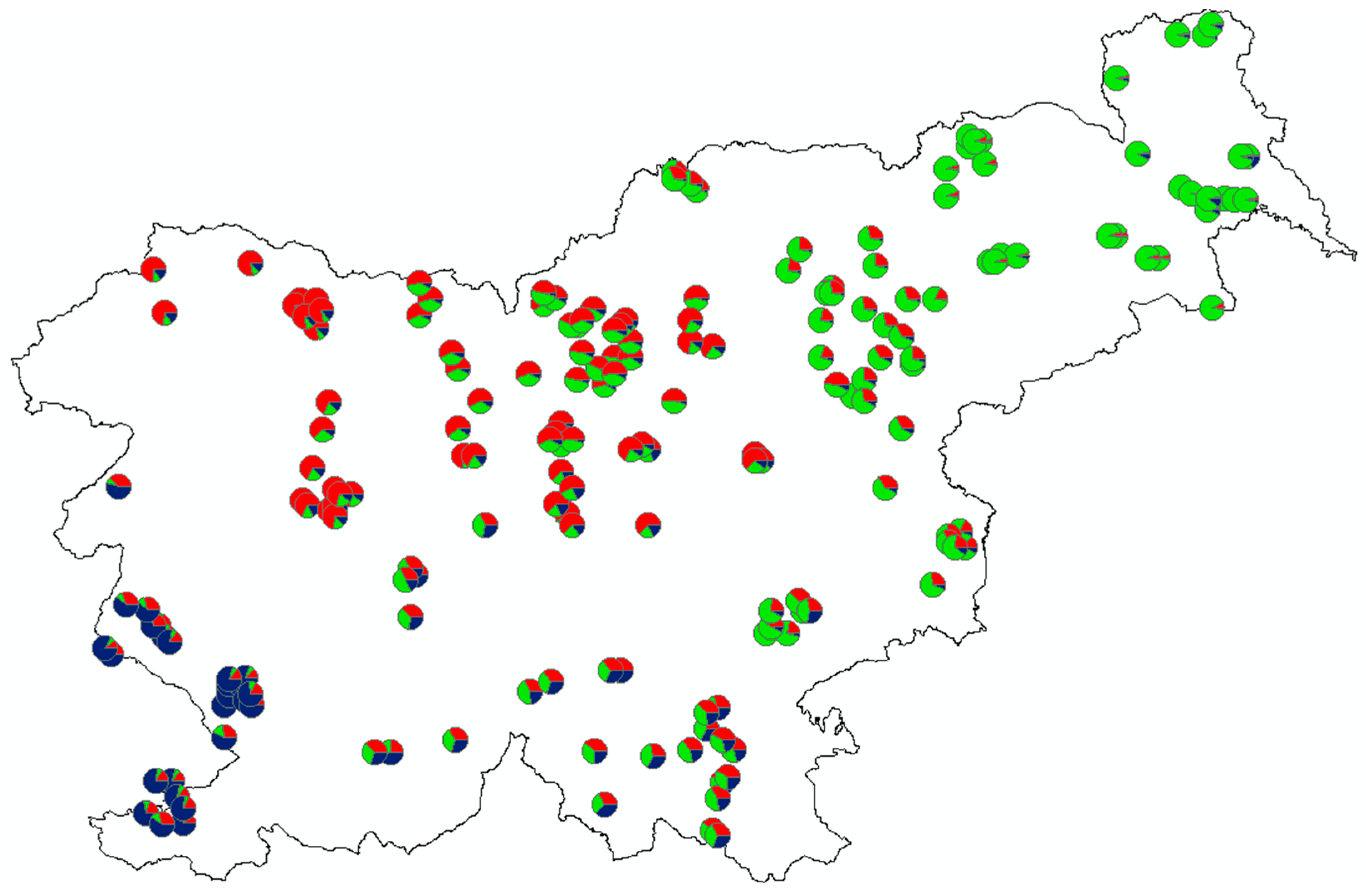
3.2. Spatial Genetic Structure
3.3. Isolation by Distance
3.4. Correlation between Individual Multilocus Heterozygosity and Fitness of Individuals
4. Discussion
4.1. Genetic Diversity of Roe Deer in Slovenia
4.2. Genetic Differentiation among Slovene Roe Deer Populations
4.3. Correlation between Genetic Traits and Fitness of Roe Deer
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Danilkin, A. Behavioural Ecology of Siberian and European Roe Deer; Chapman & Hall Press: London, UK, 1996. [Google Scholar]
- Apollonio, M.; Andersen, R.; Putman, R. European Ungulates and Their Management in the 21st Century; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Hufthammer, A.K.; Aaris–Sørensen, K. Late-and postglacial European roe deer. In The European Roe Deer: The Biology of Success; Andersen, R., Duncan, P., Linnell, J.D.C., Eds.; Scandinavian University Press: Oslo, Norway, 1998; pp. 47–69. [Google Scholar]
- Hartl, G.B.; Hewison, A.J.M.; Apollonio, M.; Kurt, F.; Wiehler, J. Genetics of European roe deer. In The European Roe Deer: The Biology of Success; Andersen, R., Duncan, P., Linnell, J.D.C., Eds.; Scandinavian University Press: Oslo, Norway, 1998; pp. 71–90. [Google Scholar]
- Wiehler, J.; Tiedemann, R. Phylogeography of the European roe deer Capreolus capreolus as revealed by sequence analysis of the mitochondrial control region. Acta Theriol. 1998, 43, 187–197. [Google Scholar] [CrossRef]
- Randi, E.; Pierpaoli, M.; Danilkin, A. Mitochondrial DNA polymorphism in populations of Siberian and European roe deer (Capreolus pygargus and C. capreolus). Heredity 1998, 80, 429–437. [Google Scholar] [CrossRef]
- Vernesi, C.; Pecchioli, E.; Caramelli, D.; Tiedemann, R.; Randi, E.; Bertorelle, G. The genetic structure of natural and reintroduced roe deer (Capreolus capreolus) populations in the Alps and central Italy, with reference to the mitochondrial DNA phylogeography of Europe. Mol. Ecol. 2002, 11, 1285–1297. [Google Scholar] [CrossRef]
- Randi, E.; Alves, P.C.; Carranza, J.; Milošević-Zlatanović, S.; Sfougrais, A.; Mucci, N. Phylogeography of roe deer (Capreolus capreolus) populations: The effects of historical genetic subdivisions and recent nonequilibrium dynamics. Mol. Ecol. 2004, 13, 3071–3083. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, R.; José, C.S.; Braza, F.; Aragón, S. Genetic differentiation and phylogeography of roe deer in Spain, as suggested by mitochondrial DNA and microsatellite analysis. Ital. J. Zool. 2003, 70, 89–99. [Google Scholar] [CrossRef]
- Plis, K.; Niedzialkowska, M.; Jędrzejewska, B. Phylogeography of roe deer (Capreolus capreolus) in central and eastern Europe. In Proceedings of the 4th Hunting and Game Management Symposium: Abstract book; Flajšman, K., Pokorny, B., Eds.; Slovenian Forestry Service, Silva Slovenica: Ljubljana, Slovenia, 2015; p. 45. [Google Scholar]
- Simonič, A. Srnjad: Biologija in Gospodarjenje; Lovska zveza Slovenije: Ljubljana, Slovenia, 1976. [Google Scholar]
- Kryštufek, B. Sesalci Slovenije; Prirodoslovni muzej Slovenije: Ljubljana, Slovenia, 1991. [Google Scholar]
- Pokorny, B. Kako še izboljšati upravljanje s srnjadjo v Sloveniji? Lovec 2009, 92, 130–134. [Google Scholar]
- Stergar, M.; Borkovič, D.; Hiršelj, J.; Kavčič, I.; Krofel, M.; Mrakič, M.; Troha, R.; Videmšek, U.; Vrčon, B.; Jerina, K. Ugotavljanje Gostot Prostoživečih Parkljarjev s Kombinirano Metodo Štetja Kupčkov Iztrebkov in Podatkov O Odvzemu; Biotehniška fakulteta, Oddelek za gozdarstvo in obnovljive gozdne vire: Ljubljana, Slovenia, 2012. [Google Scholar]
- Jerina, K.; Stergar, M.; Videmšek, U.; Kobler, A.; Pokorny, B.; Jelenko, I. Prostorska Razširjenost, Vitalnost in Populacijska Dinamika Prostoživečih Vrst Parkljarjev V Sloveniji: Preučevanje Vplivov Okoljskih in Vrstno-Specifičnih Dejavnikov Ter Napovedovanje Razvojnih Trendov; Biotehniška fakulteta, Oddelek za gozdarstvo in obnovljive gozdne vire: Ljubljana, Slovenia, 2010. [Google Scholar]
- Andersen, R.; Duncan, P.; Linnell, J.D.C. (Eds.) The European roe deer: A portrait of a successful species. In The European Roe Deer: The Biology of Success; Scandinavian University Press: Oslo, Norway, 1998; pp. 11–22. [Google Scholar]
- Maehr, D. Large Mammal. Restoration: Ecological and Sociological Challenges in the 21st Century; Maehr, D.S., Noss, R.F., Larkin, J.L., Eds.; Island Press: Washington, DC, USA, 2001. [Google Scholar]
- Harris, B.R.; Wall, W.A.; Allendorf, F.W. Genetic consequences of hunting: What do we know and what should we do? Wildl. Soc. Bull. 2002, 30, 634–643. [Google Scholar]
- Coulon, A.; Guillot, G.; Cosson, J.F.; Angibault, J.M.A.; Aulagnier, S.; Cargnelutti, B.; Galan, M.; Hewison, A.J.M. Genetic structure is influenced by landscape features: Empirical evidence from a roe deer population. Mol. Ecol. 2006, 15, 1669–1679. [Google Scholar] [CrossRef]
- Pérez-Espona, S.; Pérez-Barbería, F.J.; Mcleod, J.E.; Jiggins, C.D.; Gordon, I.J.; Pemberton, J.M. Landscape features affect gene flow of Scottish Highland red deer (Cervus elaphus). Mol. Ecol. 2008, 17, 981–996. [Google Scholar] [CrossRef]
- Breyne, P.; Mergeay, J.; Casaer, J. Roe deer population structure in a highly fragmented landscape. Eur. J. Wildl. Res. 2014, 60, 909–917. [Google Scholar] [CrossRef]
- Kuehn, R.; Hindenlang, K.E.; Holzgang, O.; Senn, J.; Stoeckle, B.; Sperisen, C. Genetic effect of transportation infrastructure on roe deer populations (Capreolus capreolus). J. Hered. 2007, 98, 13–22. [Google Scholar] [CrossRef]
- Hepenstrick, D.; Thiel, D.; Holderegger, R.; Gugerli, F. Genetic discontinuities in roe deer (Capreolus capreolus) coincide with fenced transportation infrastructure. Basic Appl. Ecol. 2012, 13, 631–638. [Google Scholar] [CrossRef]
- Kurt, F. Das Reh in der Kulturlandschaft: Sozialverhalten und Oëkologie eines Anpassers; Parey: Hamburg, Germany, 1991. [Google Scholar]
- Hewison, A.J.M.; Vincent, J.P.; Reby, D. Social organisation of European roe deer. In The European Roe Deer: The Biology of Success; Andersen, R., Duncan, P., Linnell, J.D.C., Eds.; Scandinavian University Press: Oslo, Norway, 1998; pp. 189–219. [Google Scholar]
- Strandgaard, H. An investigation of copora lutea, embryonic development, and time of birth of roe deer (Capreolus capreolus) in Denmark. Danish Rev. Game Biol. 1972, 6, 1–22. [Google Scholar]
- Ellenberg, H. The population ecology of roe deer, Capreolus capreolus (Cervidae) in Central Europe. Spixiana 1978, 2, 1–211. [Google Scholar]
- Stubbe, C. Rehwild, 3rd ed.; Deutscher Landwirtschaftsverlag: Berlin, Germany, 1990. [Google Scholar]
- Andersen, R.; Gaillard, J.M.; Linnell, J.D.C.; Duncan, P. Factors affecting maternal care in an income breeder, the European roe deer. J. Anim. Ecol. 2000, 69, 672–682. [Google Scholar] [CrossRef]
- Apollonio, M.; Merli, E.; Chirichella, R.; Pokorny, B.; Alagić, A.; Flajšman, K.; Stephens, P.A. Capital and income breeding in male ungulates: Causes and consequences of strategy differences among species. Front. Ecol. Evol. 2020, 8, 308. [Google Scholar] [CrossRef]
- Flajšman, K.; Jelenko, I.; Poličnik, H.; Pokorny, B. Reproductive potential of roe deer (Capreolus capreolus L.): Review of the most important influential factors. Acta Silvae et Ligni 2013, 102, 1–20. [Google Scholar]
- Flajšman, K.; Borowik, T.; Pokorny, B.; Jędrzejewska, B. Effects of population density and female body mass on litter size in European roe deer at a continental scale. Mammal. Res. 2018, 63, 91–98. [Google Scholar] [CrossRef]
- Flajšman, K.; Jerina, K.; Pokorny, B. Age-related effects of body mass on fertility and litter size in roe deer. PLoS ONE 2017, 12, e0175579. [Google Scholar] [CrossRef]
- Hewison, A.J.M.; Morellet, N.; Verheyden, H.; Daufresne, T.; Angibault, J.M.; Cargnelutti, B.; Merlet, J.; Picot, D.; Rames, J.L.; Joachim, J.; et al. Landscape fragmentation influences winter body mass of roe deer. Ecography 2009, 32, 1062–1070. [Google Scholar] [CrossRef]
- Flajšman, K. Effects of individual, population and environmental factors on parameters of reproductive success of female roe deer. Ph.D. Disertation, University of Ljubljana, Ljubljana, Slovenia, 2017. [Google Scholar]
- Nilsen, E.B.; Linnell, J.D.C.; Andersen, R. Individual access to preferred habitat affects fitness components in female roe deer (Capreolus capreolus). J. Anim. Ecol. 2004, 73, 44–50. [Google Scholar] [CrossRef]
- Quéméré, E.; Gaillard, J.M.; Galan, M.; Vanpé, C.; David, I.; Pellerin, M.; Kjellander, P.; Hewison, A.J.M.; Pemberton, J.M. Between-population differences in the genetic and maternal components of body mass in roe deer. BMC Evol. Biol. 2018, 18, 1–10. [Google Scholar] [CrossRef]
- Hewison, A.J.M. Evidence for a genetic component of female fecundity in British roe deer from studies of cranial morphometrics. Funct. Ecol. 1997, 11, 508–517. [Google Scholar] [CrossRef]
- Kurt, F.; Hartl, G.B.; Völk, F.P. Breeding strategies and genetic variation in European roe deer (Capreolus capreolus) populations. Acta Theriol. 1993, 38, 187. [Google Scholar] [CrossRef]
- Lamberti, P.; Mauri, L.; Apollonio, M. Two distinct patterns of spatial behaviour of female roe deer (Capreolus capreolus) in a mountainous habitat. Ethol. Ecol. Evol. 2004, 16, 41–53. [Google Scholar] [CrossRef]
- Bončina, A.; Pokorny, B.; Sternad, M.; Bužan, E. Genetic connectivity and relativeness of European roe deer in the model hunting ground of central Slovenia. Zlatorogov Zb. 2019, 6, 10–28. [Google Scholar]
- Short, R.V.; Hay, M.F. Delayed implantation in the roe deer (Capreolus capreolus). In Comparative Biology of Reproduction in Mammals; Rowlands, I.W., Ed.; Symposium of the Zoological Society of London: London, UK, 1966; pp. 173–194. [Google Scholar]
- Chirichella, R.; Pokorny, B.; Bottero, E.; Flajšman, K.; Mattioli, L.; Apollonio, M. Factors affecting implantation failure in roe deer. J. Wildl. Manag. 2019, 83, 599–609. [Google Scholar] [CrossRef]
- Ratcliffe, P.R.; Mayle, B.A. Roe deer biology and management; Forestry Commission Bulletin 105: London, UK, 1992. [Google Scholar]
- Pokorny, B.; Jelenko Turinek, I. Čeljustnice Prostoživečih Parkljarjev; Lovska zveza Slovenije: Ljubljana, Slovenia, 2018. [Google Scholar]
- Vial, L.; Maudet, C.; Luikart, G. Thirty-four polymorphic microsatellites for European roe deer. Mol. Ecol. Notes 2003, 3, 523–527. [Google Scholar] [CrossRef]
- Røed, K.H. Microsatellite variation in Scandinavian Cervidae using primers derived from Bovidae. Hereditas 1998, 129, 19–25. [Google Scholar] [CrossRef]
- Røed, K.H.; Midthjell, L. Microsatellites in reindeer, Rangifer tarandus, and their use in other cervids. Mol. Ecol. 1998, 7, 1773–1776. [Google Scholar] [CrossRef]
- Poetsch, M.; Seefeldt, S.; Maschke, M.; Lignitz, E. Analysis of microsatellite polymorphism in red deer, roe deer, and fallow deer—Possible employment in forensic applications. Forensic Sci. Int. 2001, 116, 1–8. [Google Scholar] [CrossRef]
- Buzan, E.; Potušek, S.; Urzi, F.; Pokorny, B.; Šprem, N. Genetic characterisation of wild ungulates: Successful isolation and analysis of DNA from widely available bones can be cheap, fast and easy. Zookeys 2020, 965, 141–156. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Chapuis, M.P.; Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05. Available online: http://www.genetix.univ-montp2.fr/genetix/genetix.htm (accessed on 20 June 2019).
- Goudet, J. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices, Version 2.9.3. 2001. Available online: http://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 21 June 2019).
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Jombart, T.; Collins, C. A tutorial for Discriminant Analysis of Principal Components (DAPC) Using Adegenet 2.0.0; Imperial College London, MRC Centre for Outbreak Analysis and Modelling: London, UK, 2015. [Google Scholar]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar]
- Guillot, G.; Mortier, F.; Estoup, A. GENELAND: A computer package for landscape genetics. Mol. Ecol. Notes 2005, 5, 712–715. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Latch, E.K.; Dharmarajan, G.; Glaubitz, J.C.; Rhodes, O.E. Relative performance of Bayesian clustering software for inferring population substructure and individual assignment at low levels of population differentiation. Conserv. Genet. 2006, 7, 295–302. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. RStudio, PBC, Boston. 2020. Available online: http://www.rstudio.com/ (accessed on 15 June 2019).
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 1–15. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 1–14. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef]
- Slatkin, M. Isolation by distance in equilibrium and non-equilibrium populations. Evolution 1993, 47, 264–279. [Google Scholar] [CrossRef]
- Rousset, F. Genetic differentiation and estimation of gene flow from F-statistics under isolation by distance. Genetics 1997, 145, 1219–1228. [Google Scholar]
- Mantel, N. Ranking procedures for arbitrarily restricted observation. Biometrics 1967, 23, 65–78. [Google Scholar] [CrossRef]
- Cavalli-Sforza, L.L.; Edwards, A.W. Phylogenetic analysis. Models and estimation procedures. Am. J. Hum. Genet. 1967, 19, 233–257. [Google Scholar] [CrossRef]
- Edwards, A.W.F. Distances between populations on the basis of gene frequencies. Biometrics 1971, 27, 873–881. [Google Scholar] [CrossRef]
- Aparicio, J.M.; Ortego, J.; Cordero, P.J. What should we weigh to estimate heterozygosity, alleles or loci? Mol. Ecol. 2006, 15, 4659–4665. [Google Scholar] [CrossRef]
- Coulon, A. Genhet: An easy-to-use R function to estimate individual heterozygosity. Mol. Ecol. Resour. 2010, 10, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Hewison, A.J.M.; Gaillard, J.M. Phenotypic quality and senescence affect different components of reproductive output in roe deer. J. Anim. Ecol. 2001, 70, 600–608. [Google Scholar] [CrossRef]
- Bates, D.; Kliegl, R.; Vasishth, S.; Baayen, H. Parsimonious Mixed Models. arXiv 2015, arXiv:1506.04967. [Google Scholar]
- Lorenzini, R.; Lovari, S.; Masseti, M. The rediscovery of the Italian roe deer: Genetic differentiation and management implications. Ital. J. Zool. 2002, 69, 367–379. [Google Scholar] [CrossRef]
- Kuehn, R.; Haller, H.; Schroeder, W.; Rottmann, O. Genetic roots of the red deer (Cervus elaphus) population in Eastern Switzerland. J. Hered. 2004, 95, 136–143. [Google Scholar] [CrossRef][Green Version]
- Zachos, F.E.; Hmwe, S.S.; Hartl, G.B. Biochemical and DNA markers yield strikingly different results regarding variability and differentiation of roe deer (Capreolus capreolus, Artiodactyla: Cervidae) populations from northern Germany. J. Zool. Syst. Evol. Res. 2006, 44, 167–174. [Google Scholar] [CrossRef]
- Baker, K.H.; Rus Hoelzel, A. Evolution of population genetic structure of the British roe deer by natural and anthropogenic processes (Capreolus capreolus). Ecol. Evol. 2013, 3, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Soulé, M.; Gilpin, M.; Conway, W.; Foose, T. The millenium ark: How long a voyage, how many staterooms, how many passengers? Zoo Biol. 1986, 5, 101–113. [Google Scholar] [CrossRef]
- Steinbach, P.; Heddergott, M.; Weigand, H.; Weigand, A.M.; Wilwert, E.; Stubbe, M.; Helm, B.; Campbell, R.E.; Stubbe, A.; Frantz, A.C. Rare migrants suffice to maintain high genetic diversity in an introduced island population of roe deer (Capreolus capreolus): Evidence from molecular data and simulations. Mamm. Biol. 2018, 88, 64–71. [Google Scholar] [CrossRef]
- Putman, R.; Langbein, J. The Deer Manager’s Companion: A Guide to the Management of Deer in the Wild and in Parks.; Swan-Hill Press: Shrewsbury, UK, 2003. [Google Scholar]
- Geist, V. Deer of the World; Stackpole Books: Mechanicsburg, PA, USA, 1998. [Google Scholar]
- Flajšman, K.; Pokorny, B.; Chirichella, R.; Bottero, E.; Mattioli, L.; Apollonio, M. I can produce more offspring as you can imagine: First records on exceptionally large litters in roe deer in central/southern Europe. Eur. J. Wildl. Res. 2017, 63. [Google Scholar] [CrossRef]
- Pielowski, Z. Das Feldreh–Wild der Zukunft in der Agrarlandschaft. Beitr. Jagd. Wildforsch. 1977, 10, 193–200. [Google Scholar]
- Krže, B. Srnjad: Biologija, gojitev, ekologija. In Zlatorogova Knjižnica 27; Lovska zveza Slovenije: Ljubljana, Slovenia, 2000; 271p. [Google Scholar]
- Olano-Marin, J.; Plis, K.; Sönnichsen, L.; Borowik, T.; Niedziałkowska, M.; Jędrzejewska, B. Weak population structure in European roe deer (Capreolus capreolus) and evidence of introgressive hybridization with Siberian roe deer (C. pygargus) in Northeastern Poland. PLoS ONE 2014, 9, e109147. [Google Scholar] [CrossRef]
- Narauskaite, G.; Danusevicius, D.; Petelis, K.; Brazaitis, G.; Spinkyte–Backaitiene, R.; Stankeviciute, J.; Simkevicius, K. The roe deer classification to ecotypes in mosaic landscape of Lithuania. J. Agric. For. 2015, 61, 189–196. [Google Scholar] [CrossRef]
- Debeffe, L.; Focardi, S.; Bonenfant, C.; Hewison, A.J.M.; Morellet, N.; Vanpé, C.; Heurich, M.; Kjellander, P.; Linnell, J.D.C.; Mysterud, A.; et al. A one night stand? Reproductive excursions of female roe deer as a breeding dispersal tactic. Oecologia 2014, 176, 431–443. [Google Scholar] [CrossRef]
- Wahlund, S. Zusammensetzung von Populationen und Korrelationserscheinungen vom Standpunkt der Vererbungslehre ausbetrachtet. Hereditas 1928, 11, 65–106. [Google Scholar] [CrossRef]
- Haanes, H.; Røed, A.K.H.; Flagstad, A.Ø.; Rosef, A.O. Genetic structure in an expanding cervid population after population reduction. Conserv. Genet. 2010, 11, 11–20. [Google Scholar] [CrossRef]
- Wang, M.; Schreiber, A. The impact of habitat fragmentation and social structure on the population genetics of roe deer (Capreolus capreolus L.) in Central Europe. Heredity 2001, 86, 703–715. [Google Scholar] [CrossRef]
- Coulon, A.; Cosson, J.F.; Morellet, N.; Angibault, J.M.; Cargnelutti, B.; Galan, M.; Aulagnier, S.; Hewison, A.J.M. Dispersal is not female biased in a resource-defence mating ungulate, the European roe deer. Proc. R. Soc. B Biol. Sci. 2006, 273, 341–348. [Google Scholar] [CrossRef]
- Nies, G.; Zachos, F.E.; Hartl, G.B. The impact of female philopatry on population differentiation in the European roe deer (Capreolus capreolus) as revealed by mitochondrial DNA and allozymes. Mamm. Biol. 2005, 70, 130–134. [Google Scholar] [CrossRef]
- Coltman, D.W. Molecular ecological approaches to studying the evolutionary impact of selective harvesting in wildlife. Mol. Ecol. 2008, 17, 221–235. [Google Scholar] [CrossRef]
- Cargnelutti, B.; Reby, D.; Desneux, L.; Angibault, J.M.A.; Joachim, J.; Hewison, A.J.M. Space use by roe deer in a fragmented landscape: Some preliminary results. Rev. Écol. (Terre Vie) 2002, 57, 29–36. [Google Scholar]
- José, C.S.; Lovari, S. Ranging movements of female roe deer: Do home-loving does roam to mate? Ethology 2010, 104, 721–728. [Google Scholar] [CrossRef]
- Gaillard, J.M.; Hewison, A.J.M.; Kjellander, P.; Pettorelli, N.; Bonenfant, C.; Van Moorter, B.; Liberg, O.; Andren, H.; Van Laere, G.; Klein, F.; et al. Population density and sex do not influence fine-scale natal dispersal in roe deer. Proc. R. Soc. B Biol. Sci. 2008, 275, 2025–2030. [Google Scholar] [CrossRef]
- Bonnot, N.; Gaillard, J.M.; Coulon, A.; Galan, M.; Cosson, J.F.; Delorme, D.; Klein, F.; Hewison, A.J.M. No difference between the sexes in fine-scale spatial genetic structure of roe deer. PLoS ONE 2010, 5, e14436. [Google Scholar] [CrossRef]
- Coulon, A.; Cosson, J.F.; Angibault, J.M.; Cargnelutti, B.; Galan, M.; Morellet, N.; Petit, E.; Aulagnier, S.; Hewison, A.J.M. Landscape connectivity influences gene flow in a roe deer population inhabiting a fragmented landscape: An individual-based approach. Mol. Ecol. 2004, 13, 2841–2850. [Google Scholar] [CrossRef]
- Pokorny, B.; Flajšman, K.; Centore, L.; Krope, F.S.; Šprem, N. Border fence: A new ecological obstacle for wildlife in Southeast Europe. Eur. J. Wildl. Res. 2017, 63, 1–6. [Google Scholar] [CrossRef]
- Debeffe, L.; Morellet, N.; Cargnelutti, B.; Lourtet, B.; Coulon, A.; Gaillard, J.M.; Bon, R.; Hewison, A.J.M. Exploration as a key component of natal dispersal: Dispersers explore more than philopatric individuals in roe deer. Anim. Behav. 2013, 86, 143–151. [Google Scholar] [CrossRef]
- Toïgo, C.; Gaillard, J.M.; Van Laere, G.; Hewison, A.J.M.; Morellet, N. How does environmental variation influence body mass, body size, and body condition? Roe deer as a case study. Ecography 2006, 29, 301–308. [Google Scholar] [CrossRef]
- Andersen, R.; Gaillard, J.M.; San José, C. Variation in life-history parameters. In The European Roe Deer: The Biology of Success; Andersen, R., Duncan, P., Linnell, J.D.C., Eds.; Scandinavian University Press: Oslo, Norway, 1998; pp. 285–307. [Google Scholar]
- Douhard, M.; Gaillard, J.M.; Delorme, D.; Capron, G.; Duncan, P.; Klein, F.; Bonenfant, C. Variation in adult body mass of roe deer: Early environmental conditions influence early and late body growth of females. Ecology 2013, 94, 1805–1814. [Google Scholar] [CrossRef]
- Chapman, G.; Bork, E.; Donkor, N.; Hudson, R. Yields, quality and suitability of four annual forages for deer pasture in north Central Alberta. Open Agric. J. 2009, 3, 26–31. [Google Scholar] [CrossRef]
- Da Silva, A.; Gaillard, J.M.; Yoccoz, N.G.; Hewison, A.J.M.; Galan, M.; Coulson, T.; Allainé, D.; Vial, L.; Delorme, D.; Van Laere, G.; et al. Heterozygosity-fitness correlations revealed by neutral and candidate gene markers in roe deer from a long-term study. Evolution 2009, 63, 403–417. [Google Scholar] [CrossRef]
- Vanpé, C.; Gaillard, J.M.; Morellet, N.; Kjellander, P.; Liberg, O.; Delorme, D.; Hewison, A.J.M. Age-specific variation in male breeding success of a territorial ungulate species, the European roe deer. J. Mammal. 2009, 90, 661–665. [Google Scholar] [CrossRef]
- Brommer, J.E.; Kekkonen, J.; Wikström, M. Using heterozygosity-fitness correlations to study inbreeding depression in an isolated population of white-tailed deer founded by few individuals. Ecol. Evol. 2015, 5, 357–367. [Google Scholar] [CrossRef]
- Brambilla, A.; Biebach, I.; Bassano, B.; Bogliani, G.; von Hardenberg, A. Direct and indirect causal effects of heterozygosity on fitness-related traits in Alpine ibex. Proc. R. Soc. B Biol. Sci. 2014, 282, 20141873. [Google Scholar] [CrossRef]
- Vanpé, C.; Debeffe, L.; Hewison, A.J.M.; Quéméré, E.; Lemaître, J.F.; Galan, M.; Amblard, B.; Klein, F.; Cargnelutti, B.; Capron, G.; et al. Reduced microsatellite heterozygosity does not affect natal dispersal in three contrasting roe deer populations. Oecologia 2015, 177, 631–643. [Google Scholar] [CrossRef]
- Gillingham, M.A.; Cézilly, F.; Wattier, R.; Béchet, A. Evidence for an association between post-fledging dispersal and microsatellite multilocus heterozygosity in a large population of greater flamingos. PLoS ONE 2013, 8, e81118. [Google Scholar] [CrossRef]
- Gaillard, J.M.; Boutin, J.M.; Delorme, D.; Van Laere, G.; Duncan, P.; Lebreton, J.D. Early survival in roe deer: Causes and consequences of cohort variation in two contrasted populations. Oecologia 1997, 112, 502–513. [Google Scholar] [CrossRef]
- Plard, F.; Gaillard, J.M.; Coulson, T.; Hewison, A.J.M.; Delorme, D.; Warnant, C.; Bonenfant, C. Mismatch between birth date and vegetation phenology slows the demography of roe deer. PLoS Biol. 2014, 12, e1001828. [Google Scholar] [CrossRef]
- Jarnemo, A. Roe deer Capreolus capreolus fawns and mowing – mortality rates and countermeasures. Wildl. Biol. 2002, 8, 211–218. [Google Scholar] [CrossRef]
- Jarnemo, A. Predation processes: Behavioural interactions between red fox and roe deer during the fawning season. J. Ethol. 2004, 22, 167–173. [Google Scholar] [CrossRef]
- Panzacchi, M.; Linnell, J.D.C.; Odden, J.; Odden, M.; Andersen, R. When a generalist becomes a specialist: Patterns of red fox predation on roe deer fawns under contrasting conditions. Can. J. Zool. 2008, 86, 116–126. [Google Scholar] [CrossRef]
- Cukor, J.; Havránek, F.; Vacek, Z.; Bukovjan, K.; Podrázský, V.; Sharma, R.P. Roe deer (Capreolus capreolus) mortality in relation to fodder harvest in agricultural landscape. Mammalia 2019, 83, 461–469. [Google Scholar] [CrossRef]
- Oslis–Central Slovene Hunting Information System. Available online: www.osilis.gozdis.si (accessed on 20 October 2020).
- Pokorny, B.; Jelenko, I.; Poličnik, H. Spatial and temporal variability of roe deer body mass in Slovenia. In Proceedings of the 9th European Roe Deer Congress, Edinburgh, Scotland, 5–10 July 2019; p. 28. [Google Scholar]
- Flajšman, K.; Pokorny, B. Kaj vemo o razmnoževalnem potencialu srnjadi v Sloveniji? Lovec 2017, 100, 10–14. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Geographical Area | Abbr. | n | He ± SD | Ho ± SD | FIS | HWE (p-Values) | A ± SD | AR ± SD |
|---|---|---|---|---|---|---|---|---|
| Coastal Slovenia (Kras and Istra) | S1 | 33 | 0.669 ± 0.139 | 0.655 ± 0.145 | 0.021 | 0.157 | 6.273 ± 2.494 | 4.817 ± 1.804 |
| Julian Alps | C1 | 10 | 0.610 ± 0.154 | 0.646 ± 0.216 | −0.061 | 0.286 | 4.455 ± 1.968 | 4.455 ± 1.968 |
| Polhograjsko and Škofjeloško Hills | C2 | 15 | 0.656 ± 0.155 | 0.610 ± 0.157 | 0.073 | 0.500 | 5.455 ± 2.115 | 4.973 ± 1.836 |
| Dinaric Mountains | C3 | 27 | 0.623 ± 0.171 | 0.577 ± 0.184 | 0.075 | 0.569 | 5.545 ± 2.207 | 4.506 ± 1.747 |
| Kamniško-Savinjske Alps | C4 | 34 | 0.652 ± 0.160 | 0.642 ± 0.204 | 0.015 | 0.326 | 6.545 ± 2.659 | 5.013 ± 1.786 |
| Posavsko Hills | C5 | 16 | 0.630 ± 0.139 | 0.640 ± 0.131 | −0.017 | 0.629 | 5.364 ± 2.292 | 4.738 ± 1.901 |
| Pohorje | N1 | 21 | 0.622 ± 0.185 | 0.641 ± 0.207 | −0.031 | 0.802 | 5.636 ± 2.501 | 4.683 ± 1.967 |
| Sub-Pannonian Region | N2 | 25 | 0.629 ± 0.174 | 0.618 ± 0.210 | 0.017 | 0.555 | 6.000 ± 2.490 | 4.865 ± 1.951 |
| Podravje and Slovenske gorice | N3 | 16 | 0.632 ± 0.250 | 0.611 ± 0.239 | 0.035 | 0.013 | 5.364 ± 2.767 | 4.782 ± 2.423 |
| Prekmurje | N4 | 17 | 0.618 ± 0.201 | 0.571 ± 0.222 | 0.078 | 0.566 | 5.273 ± 2.37 | 4.658 ± 1.949 |
| Population | S1 | C1 | C2 | C3 | C4 | C5 | N1 | N2 | N3 |
|---|---|---|---|---|---|---|---|---|---|
| C1 | 0.007 | ||||||||
| C2 | 0.033 | 0.012 | |||||||
| C3 | 0.011 | 0.006 | 0.022 | ||||||
| C4 | 0.020 | −0.002 | 0.012 | 0.006 | |||||
| C5 | 0.024 | 0.000 | 0.024 | 0.012 | 0.012 | ||||
| N1 | 0.030 | 0.019 | 0.028 | 0.001 | −0.002 | 0.021 | |||
| N2 | 0.020 | 0.008 | 0.013 | 0.004 | −0.001 | 0.012 | 0.002 | ||
| N3 | 0.040 | 0.040 | 0.031 | 0.019 | 0.017 | 0.035 | 0.008 | 0.006 | |
| N4 | 0.028 | 0.026 | 0.033 | 0.022 | 0.032 | 0.038 | 0.024 | 0.018 | 0.009 |
| Source of Variation | Variance |
|---|---|
| Within individuals | 94.92 (<0.0004) |
| Among individuals within populations | 3.39 (<0.008) |
| Among populations within groups | 1.20 (<0.0001) |
| Among groups | 0.48 (<0.103) |
| Model 1 (The Best Model): p < 0.001, R2 = 0.188 | ||||
|---|---|---|---|---|
| Variable | Estimate | Standard Error | t-Value | p-Value |
| Intercept | 14.6247 | 0.5768 | 25.355 | <0.001 |
| HL | 2.6683 | 1.2856 | 2.076 | 0.039 |
| q1 | 3.8706 | 0.7453 | 5.193 | <0.001 |
| q3 | 3.0237 | 1.1870 | −2.547 | 0.012 |
| Model 2: p < 0.001, R2 = 0.116 | ||||
| Intercept | 14.1964 | 0.5608 | 25.314 | <0.001 |
| HL | 2.9129 | 1.3030 | 2.235 | 0.027 |
| q1 | 3.9410 | 0.7570 | −5.206 | <0.001 |
| Model 3: p = 0.002, R2 = 0.061 | ||||
| Intercept | 13.0893 | 0.5326 | 24.576 | <0.001 |
| HL | 3.2131 | 1.3779 | 2.332 | 0.021 |
| q3 | 3.2522 | 1.2756 | −2.549 | 0.012 |
| Model 4: p = 0.014, R2 = 0.030 | ||||
| Intercept | 12.5979 | 0.5046 | 24.965 | <0.001 |
| HL | 3.4872 | 31.3961 | 2.498 | 0.013 |
| Fertility | ||||
|---|---|---|---|---|
| Best Model: AIC = 74.65; R2 = 0.192 | ||||
| Variable | Estimate | Standard Error | t-Value | p-Value |
| Intercept | −2769.2212 | 950.8732 | −2.912 | 0.003 |
| HL | 1.4502 | 2.5843 | 0.561 | 0.575 |
| q1 | 5.0109 | 2.1728 | −2.306 | 0.021 |
| q3 | 16.7065 | 12.3516 | 1.353 | 0.176 |
| year | 1.3764 | 0.4723 | 2.914 | 0.004 |
| Potential Reproductive Output | ||||
| Best Model: AIC = 212.3; R2 = 0.037 | ||||
| Intercept | 0.6676 | 0.5435 | 1.228 | 0.219 |
| HL | 0.9163 | 1.2725 | 0.720 | 0.471 |
| q1 | −1.6956 | 0.7565 | −2.241 | 0.025 |
| Model 2: AIC = 214.2; R2 = 0.038 | ||||
| Intercept | 0.7179 | 0.5721 | 1.255 | 0.209 |
| HL | 0.8885 | 1.2765 | 0.696 | 0.486 |
| q1 | −1.6916 | 0.7579 | −2.232 | 0.026 |
| q3 | −0.3268 | 1.1401 | −0.287 | 0.774 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buzan, E.; Gerič, U.; Potušek, S.; Flajšman, K.; Pokorny, B. First Insights into the Population Genetic Structure and Heterozygosity–Fitness Relationship in Roe Deer Inhabiting the Area between the Alps and Dinaric Mountains. Animals 2020, 10, 2276. https://doi.org/10.3390/ani10122276
Buzan E, Gerič U, Potušek S, Flajšman K, Pokorny B. First Insights into the Population Genetic Structure and Heterozygosity–Fitness Relationship in Roe Deer Inhabiting the Area between the Alps and Dinaric Mountains. Animals. 2020; 10(12):2276. https://doi.org/10.3390/ani10122276
Chicago/Turabian StyleBuzan, Elena, Urška Gerič, Sandra Potušek, Katarina Flajšman, and Boštjan Pokorny. 2020. "First Insights into the Population Genetic Structure and Heterozygosity–Fitness Relationship in Roe Deer Inhabiting the Area between the Alps and Dinaric Mountains" Animals 10, no. 12: 2276. https://doi.org/10.3390/ani10122276
APA StyleBuzan, E., Gerič, U., Potušek, S., Flajšman, K., & Pokorny, B. (2020). First Insights into the Population Genetic Structure and Heterozygosity–Fitness Relationship in Roe Deer Inhabiting the Area between the Alps and Dinaric Mountains. Animals, 10(12), 2276. https://doi.org/10.3390/ani10122276