Semen Modulates Inflammation and Angiogenesis in the Reproductive Tract of Female Rabbits

 , ,
, ,  ,
,  ,
,  and
and
Abstract
Simple Summary
Abstract
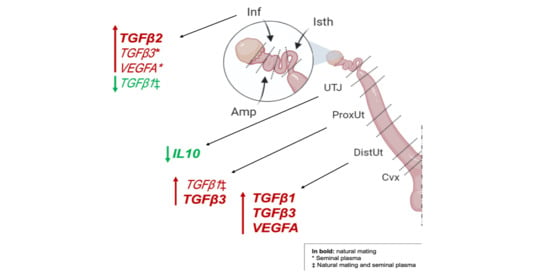
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Experimental Design
2.3. Mating and Semen Collection
2.4. Tissue Sample Collection
2.5. Quantitative PCR Analyses
2.6. Statistical Analyses
3. Results
3.1. Natural Mating and Sperm-Free Seminal Plasma Infusion Differentially Modulated Anti-Inflammatory Cytokines and VEGF-A Genes
3.2. Temporal Gene Expression of Anti-Inflammatory cytokines and VEGF-A at 10 h up to 72 h Post-Mating
3.3. Spatial Gene Expression Triggered by Natural Mating and Sperm-Free Seminal Plasma Infusion
3.4. Spatial Gene Expression at 10 h up to 72 h Post-Mating
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- United States Department of Agriculture. Annual Report Animal Usage by Fiscal Year. Available online: https://www.aphis.usda.gov/animal_welfare/downloads/reports/Annual-Report-Animal-Usage-by-FY2017.pdf (accessed on 29 May 2020).
- Roy-Dufresne, E.; Lurgi, M.; Brown, S.C.; Wells, K.; Cooke, B.; Mutze, G.; Peacock, D.; Cassey, P.; Berman, D.; Brook, B.W.; et al. The Australian National Rabbit Database: 50 yr of population monitoring of an invasive species. Ecology 2019, 100, e2750. [Google Scholar] [CrossRef] [PubMed]
- Petracci, M.; Soglia, F.; Leroy, F. Rabbit meat in need of a hat-trick: From tradition to innovation (and back). Meat Sci. 2018, 146, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, P.L.; García-García, R.M.; Árias-Álvarez, M.; Rebollar, P.G. Reproductive and nutritional management on ovarian response and embryo quality on rabbit does. Reprod. Domest. Anim. 2014, 49, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J.; Baum, M.J. Neuroendocrine regulation of GnRH release in induced ovulators. Front. Neuroendocrinol. 2000, 21, 220–262. [Google Scholar] [CrossRef] [PubMed]
- Ratto, M.H.; Berland, M.; Silva, M.E.; Adams, G.P. New insights of the role of B-NGF in the ovulation mechanism of induced ovulating species. Reproduction 2019, 157, R199–R207. [Google Scholar] [CrossRef] [PubMed]
- Dukelow, W.R.; Williams, W.L. Survival of Capacitated Spermatozoa in the Oviduct of the Rabbit. J. Reprod. Fert. 1967, 14, 477–479. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Morrell, J.M. Artificial insemination in rabbits. Br. Vet. J. 1995, 151, 477–488. [Google Scholar] [CrossRef]
- Mocé, E.; Vicente, J.S. Rabbit sperm cryopreservation: A review. Anim. Reprod. Sci. 2009, 110, 1–24. [Google Scholar] [CrossRef]
- Schjenken, J.E.; Robertson, S.A. Seminal Fluid Signalling in the Female Reproductive Tract: Implications for Reproductive Success and Offspring Health. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2015; Volume 868, pp. 127–158. ISBN 978-3-319-18880-5. [Google Scholar]
- Parada-Bustamante, A.; Oróstica, M.L.; Reuquen, P.; Zuñiga, L.M.; Cardenas, H.; Orihuela, P.A. The role of mating in oviduct biology. Mol. Reprod. Dev. 2016, 83, 875–883. [Google Scholar] [CrossRef]
- Ghersevich, S.; Massa, E.; Zumoffen, C. Oviductal secretion and gamete interaction. Reproduction 2015, 149, R1–R14. [Google Scholar] [CrossRef]
- Suarez, S.S. Interactions of Gametes with the Female Reproductive Tract. Cell Tissue Res 2016, 363, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Rodriguez, M.; Atikuzzaman, M.; Venhoranta, H.; Wright, D.; Rodriguez-Martinez, H. Expression of Immune Regulatory Genes in the Porcine Internal Genital Tract Is Differentially Triggered by Spermatozoa and Seminal Plasma. Int. J. Mol. Sci. 2019, 20, 513. [Google Scholar] [CrossRef] [PubMed]
- Katila, T. Post-mating Inflammatory Responses of the Uterus. Reprod. Domest. Anim. 2012, 47, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. The Immunology of Early Pregnancy in Farm Animals. Reprod. Domest. Anim. 2011, 46, 18–30. [Google Scholar] [CrossRef] [PubMed]
- McDonald, L.E.; Black, W.G.; Casida, L.E. The Response of the Rabbit Uterus to Instillation of Semen at Different Phases of the Estrous Cycle. Am. J. Vet. Res. 1952, 13, 419–424. [Google Scholar] [PubMed]
- Phillips, D.M.; Mahler, S. Phagocytosis of spermatozoa by the rabbit vagina. Anat. Rec. 1977, 189, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.M.; Mahler, S. Leukocyte emigration and migration in the vagina following mating in the rabbit. Anat. Rec. 1977, 189, 45–59. [Google Scholar] [CrossRef]
- El-Banna, A.A.; Hafez, E.S.E. Sperm transport and distribution in rabbit and cattle female tract. Fertil. Steril. 1970, 21, 534–540. [Google Scholar] [CrossRef]
- Yousef, M.S.; Marey, M.A.; Hambruch, N.; Hayakawa, H.; Shimizu, T.; Hussien, H.A.; Abdel-Razek, A.R.K.; Pfarrer, C.; Miyamoto, A. Sperm binding to oviduct epithelial cells enhances TGFB1 and IL10 expressions in epithelial cells aswell as neutrophils in vitro: Prostaglandin E2 as a main regulator of anti-inflammatory response in the bovine oviduct. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Ingman, W.V.; Robertson, S.A. Defining the actions of transforming growth factor beta in reproduction. BioEssays 2002, 24, 904–914. [Google Scholar] [CrossRef]
- Massagué, J. TGF-β signal transduction. Annu. Rev. Biochem. 1998, 67, 753–791. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Bond, M.W.; Mosmann, T.R. Two types of mouse T helper cell. IV. Th2 clones secrete a factor that inhibits cytokine production by Th1 clones. J. Exp. Med. 1989, 170, 2081–2095. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Houck, K.; Jakeman, L.; Leung, D.W. Molecular and biological properties of the vascular endothelial growth factor family of proteins. Endocr. Rev. 1992, 13, 18–32. [Google Scholar] [CrossRef]
- Senger, D.R.; Perruzzi, C.A.; Feder, J.; Dvorak, H.F. A Highly Conserved Vascular Permeability Factor Secreted by a Variety of Human and Rodent Tumor Cell Lines. Cancer Res. 1986, 46, 5629–5632. [Google Scholar]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Rabbani, M.; Rogers, P. Role of vascular endothelial growth factor in endometrial vascular events before implantation in rats. Reproduction 2001, 122, 85–90. [Google Scholar] [CrossRef]
- Welter, H.; Wollenhaupt, K.; Tiemann, U.; Einspanier, R. Regulation of the VEGF-System in the Endometrium during Steroid-Replacement and Early Pregnancy of Pigs. Exp. Clin. Endocrinol. Diabetes 2003, 111, 33–40. [Google Scholar] [CrossRef]
- Chennazhi, K.; Nayak, N. Regulation of Angiogenesis in the Primate Endometrium: Vascular Endothelial Growth Factor. Semin. Reprod. Med. 2009, 27, 080–089. [Google Scholar] [CrossRef]
- Das, S.K.; Chakraborty, I.; Wang, J.; Dey, S.K.; Hoffman, L.H. Expression of Vascular Endothelial Growth Factor (VEGF) and VEGF-Receptor Messenger Ribonucleic Acids in the Peri-Implantation Rabbit Uterus. Biol. Reprod. 1997, 56, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Gardela, J.; Jauregi-Miguel, A.; Martinez, C.A.; Rodriguez-Martinez, H.; Lopez-Bejar, M.; Alvarez-Rodriguez, M. Semen Modulates the Expression of NGF, ABHD2, VCAN, and CTEN in the Reproductive Tract of Female Rabbits. Genes 2020, 11, 758. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.r-project.org/ (accessed on 10 August 2020).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarjar, D.; Team, R.C. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-145. Available online: https://cran.r-project.org/package=nlme (accessed on 10 August 2020).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Robertson, S.A.; Sharkey, D.J. The role of semen in induction of maternal immune tolerance to pregnancy. Semin. Immunol. 2001, 13, 243–254. [Google Scholar] [CrossRef]
- Robertson, S.A.; Ingman, W.V.; O’Leary, S.; Sharkey, D.J.; Tremellen, K.P. Transforming growth factor β—A mediator of immune deviation in seminal plasma. J. Reprod. Immunol. 2002, 57, 109–128. [Google Scholar] [CrossRef]
- Overstreet, J.W.; Cooper, G.W. Sperm Transport in the Reproductive Tract of the Female Rabbit: I. The Rapid Transit Phase of Transport. Biol. Reprod. 1978, 19, 101–114. [Google Scholar] [CrossRef]
- Overstreet, J.W.; Cooper, G.W. Sperm Transport in the Reproductive Tract of the Female Rabbit: II. The Sustained Phase of Transport. Biol. Reprod. 1978, 19, 115–132. [Google Scholar] [CrossRef]
- Greenwald, G.S. A study of the transport of ova through the rabbit oviduct. Fertil. Steril. 1961, 12, 80–95. [Google Scholar] [CrossRef]
- Roca, J.; Martínez, S.; Orengo, J.; Parrilla, I.; Vázquez, J.M.; Martínez, E.A. Influence of constant long days on ejaculate parameters of rabbits reared under natural environment conditions of Mediterranean area. Livest. Prod. Sci. 2005, 94, 169–177. [Google Scholar] [CrossRef]
- McNitt, J.I. Rabbit Management. In Rabbit Production; McNitt, J.I., Lukefahr, S.D., Cheeke, P.R., Patton, H.M., Eds.; CABI: Boston, MA, USA, 2013; pp. 42–61. [Google Scholar]
- Pincus, G.; Enzemann, E. V Fertilisation in the Rabbit. J. Exp. Biol. 1932, 9, 403–408. [Google Scholar]
- Duffy, D.M.; Ko, C.; Jo, M.; Brannstrom, M.; Curry, T.E. Ovulation: Parallels With Inflammatory Processes. Endocr. Rev. 2019, 40, 369–416. [Google Scholar] [CrossRef] [PubMed]
- Braden, A.W.H. Distribution of sperms in the genital tract of the female rabbit after coitus. Aust. J. Biol. Sci. 1953, 6, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Morton, D.B.; Glover, T.D. Sperm transport in the female rabbit: The role of the cervix. J. Reprod. Fertil. 1974, 38, 131–138. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ashary, N.; Tiwari, A.; Modi, D. Embryo Implantation: War in Times of Love. Endocrinology 2018, 159, 1188–1198. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Man, G.C.W.; Liu, Y.; Wu, F.; Huang, J.; Li, T.C.; Wang, C.C. Physiological and pathological angiogenesis in endometrium at the time of embryo implantation. Am. J. Reprod. Immunol. 2017, 78, e12693. [Google Scholar] [CrossRef]
- Waldhalm, S.J.; Dickson, W.M. Blood Flow to the Uterine Tube (Oviduct) of the Early Pregnant and Pseudopregnant Rabbit. Am. J. Vet. Res. 1976, 37, 817–821. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | Product Size (bp) | Efficiency (%) |
|---|---|---|---|
| β-ACTIN | F: commercial, not available | 120 | 88.6 |
| R: commercial, not available | |||
| IL10 | F: TTGTTAACCGAGTCCCTGCT | 209 | 107.5 |
| R: TTTTCACAGGGGAGAAATCG | |||
| TGFβ1 | F: CACCATTCACAGCATGAACC | 129 | 99.3 |
| R: TTCTCTGTGGAGCTGAAGCA | |||
| TGFβ2 | F: CCGGAGGTGATCTCCATCTA | 236 | 94.7 |
| R: AGGTACCCACAGAGCACCTG | |||
| TGFβ3 | F: CACGAACCCAAGGGCTACTA | 186 | 97.5 |
| R: GGTCCTCCCGACGTAGTACA | |||
| VEGF-A | F: GGAGACAATAAACCCCACGA | 219 | 87.0 |
| R: CTGCATGGTGACGTTGAACT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gardela, J.; Jauregi-Miguel, A.; Martinez, C.A.; Rodríguez-Martinez, H.; López-Béjar, M.; Álvarez-Rodríguez, M. Semen Modulates Inflammation and Angiogenesis in the Reproductive Tract of Female Rabbits. Animals 2020, 10, 2207. https://doi.org/10.3390/ani10122207
Gardela J, Jauregi-Miguel A, Martinez CA, Rodríguez-Martinez H, López-Béjar M, Álvarez-Rodríguez M. Semen Modulates Inflammation and Angiogenesis in the Reproductive Tract of Female Rabbits. Animals. 2020; 10(12):2207. https://doi.org/10.3390/ani10122207
Chicago/Turabian StyleGardela, Jaume, Amaia Jauregi-Miguel, Cristina A. Martinez, Heriberto Rodríguez-Martinez, Manel López-Béjar, and Manuel Álvarez-Rodríguez. 2020. "Semen Modulates Inflammation and Angiogenesis in the Reproductive Tract of Female Rabbits" Animals 10, no. 12: 2207. https://doi.org/10.3390/ani10122207
APA StyleGardela, J., Jauregi-Miguel, A., Martinez, C. A., Rodríguez-Martinez, H., López-Béjar, M., & Álvarez-Rodríguez, M. (2020). Semen Modulates Inflammation and Angiogenesis in the Reproductive Tract of Female Rabbits. Animals, 10(12), 2207. https://doi.org/10.3390/ani10122207