The Impact of BPI Expression on Escherichia coli F18 Infection in Porcine Kidney Cells



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. BPI Overexpression in PK15 Cells
2.3. LPS and E. coli F18 Stimulation
2.4. Analysis of E. coli F18 Adhesion to PK15 Cells
2.4.1. Colony Counting
2.4.2. Fluorescence Quantitative Polymerase Chain Reaction
2.5. Primer Design
2.6. RNA Extraction and Preparation
2.7. qPCR
2.8. Cytokine ELISAs
2.9. Statistical Analysis
3. Results
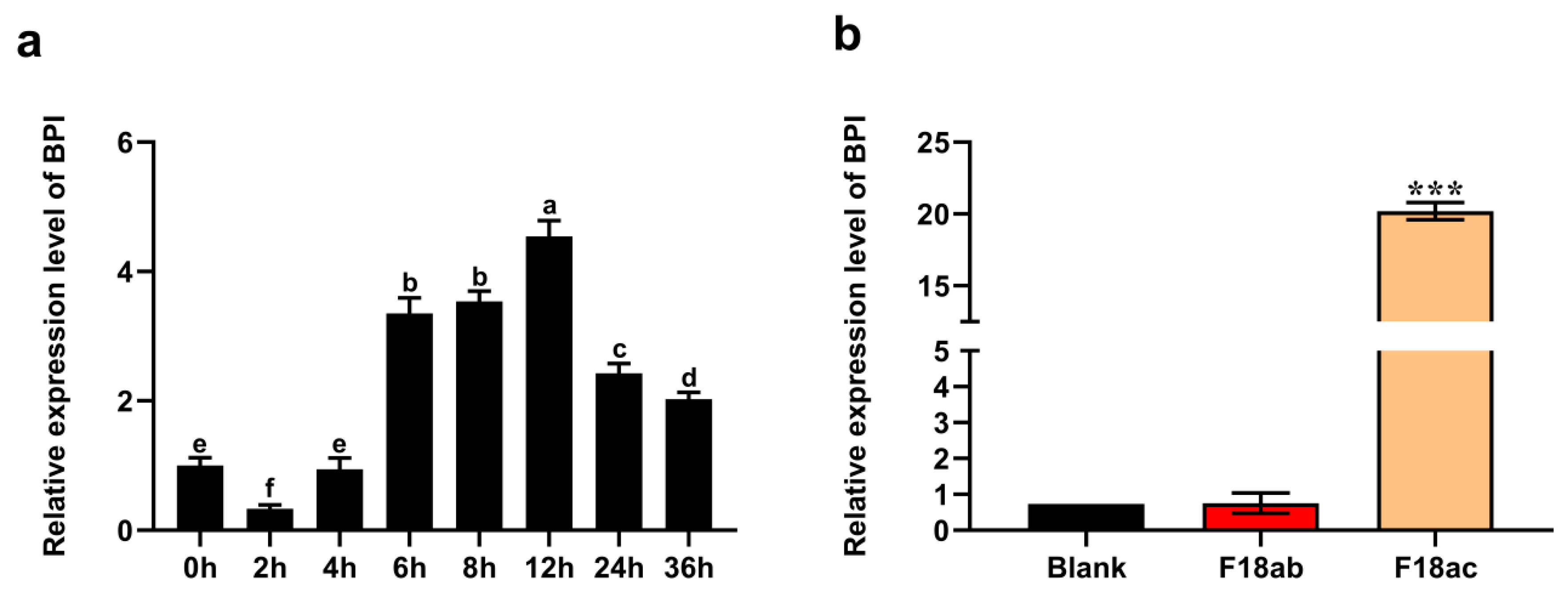
3.1. LPS and E. coli F18 Induce BPI Upregulation in PK15 Cells
3.2. Preparation of BPI-Overexpressing PK15 Cells
3.3. The Impact of BPI Overexpression on E. coli F18 Adhesion to PK15 Cells
3.4. The Impact of BPI Overexpression on TLR4 and Glycosphingolipid Biosynthesis-Globo Series Pathway-Related Gene Expression
3.5. The Impact of BPI Overexpression on the Upregulation of TLR4-Related Proinflammatory Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Weiss, J.; Elsbach, P.; Olsson, I.; Odeberg, H. Purification and characterization of a potent bactericidal and membrane active protein from the granules of human polymorphonuclear leukocytes. J. Biol. Chem. 1978, 253, 2664–2672. [Google Scholar]
- Nishimura, H.; Gogami, A.; Miyagawa, Y.; Nanbo, A.; Murakami, Y.; Baba, T.; Nagasawa, S. Bactericidal/permeability-increasing protein promotes complement activation for neutrophil-mediated phagocytosis on bacterial surface. Immunology 2001, 103, 519–525. [Google Scholar] [CrossRef]
- Wilde, C.G.; Seilhamer, J.J.; McGrogan, M.; Ashton, N.; Snable, J.L.; Lane, J.C.; Leong, S.R.; Thornton, M.B.; Miller, K.L.; Scott, R.W.; et al. Bactericidal/permeability-increasing protein and lipopolysaccharide (LPS)-binding protein. LPS binding properties and effects on LPS-mediated cell activation. J. Biol. Chem. 1994, 269, 17411–17416. [Google Scholar]
- Fan, L.; Fu, G.; Ding, Y.Y.; Lv, P.; Li, H. Bactericidal/permeability increasing protein gene polymorphism and inflammatory bowel diseases: Meta-analysis of five case–control studies. Int. J. Colorectal. Dis. 2017, 32, 433–435. [Google Scholar] [CrossRef]
- Esteve, E.; Castro, A.; Moreno, J.M.; Vendrell, J.; Ricart, W.; Fernández-Real, J.M. Circulating bactericidal/permeability-increasing protein (BPI) is associated with serum lipids and endothelial function. Thromb. Haemost. 2010, 103, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Der Schaft, D.W.; Wagstaff, J.; Mayo, K.H.; Griffioen, A.W. The antiangiogenic properties of bactericidal/permeability-increasing protein (BPI). Ann. Med. 2002, 34, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, M.; Rook, S.L.; Sassa, Y.; Ma, R.C.; Geraldes, P.; Goddard, L.; Clermont, A.; Gao, B.; Salti, H.; Gundel, R.; et al. Bactericidal/permeability-increasing protein’s signaling pathways and its retinal trophic and anti-angiogenic effects. FASEB J. 2006, 20, 2058–2067. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Wu, Y.F.; Tang, L.; He, Y.; Shi, L.; Xiong, F.; Gong, Z.J.; Guo, C.; Li, X.Y.; Liao, Q.J.; et al. BPIFB1 (LPLUNC1) inhibits migration and invasion of nasopharyngeal carcinoma by interacting with VTN and VIM. Br. J. Cancer 2018, 118, 233–247. [Google Scholar] [CrossRef] [PubMed]
- De Smet, E.G.; Seys, L.J.; Verhamme, F.M.; Vanaudenaerde, B.M.; Brusselle, G.G.; Bingle, C.D.; Bracke, K.R. Association of innate defense proteins bpifa1 and bpifb1 with disease severity in copd. Int. J. Chronic Obstruct. Pulm. Dis. 2017, 13, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Little, M.S.; Redinbo, M.R. Crystal structure of the mouse innate immunity factor bacterial permeability-increasing family member a1. Acta Crystallogr. F Struct. Biol. Commun. 2018, 74, 268–276. [Google Scholar] [CrossRef]
- Leclair, E.E. Four BPI (bactericidal/permeability-increasing protein)-like genes expressed in the mouse nasal, oral, airway and digestive epithelia. Biochem. Soc. Trans. 2003, 31, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Seo, H.; Moxley, R.A.; Zhang, W. Mapping the neutralizing epitopes of F18 fimbrial adhesin subunit FedF of enterotoxigenic Escherichia coli (ETEC). Vet. Microbiol. 2019, 230, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.; Surendran, N.; Pichichero, M. Immune responses in neonates. Expert Rev. Clin. Immunol. 2014, 10, 1171–1184. [Google Scholar] [CrossRef] [PubMed]
- Fairbrother, J.M.; Nadeau, É.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef]
- Zhu, J.; Zi, C.; Wu, Z.C.; Liu, L.; Zheng, X.R.; Su, X.M.; Zhu, G.Q.; Huang, X.G.; Wu, S.L.; Bao, W.B. Age-dependent expression of the BPI gene in Sutai piglets. Genet. Mol. Res. 2013, 12, 2120–2126. [Google Scholar] [CrossRef]
- Sun, L.; Wang, J.; Yin, X.M.; Sun, S.Y.; Zi, C.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Identification of a 5-Methylcytosine Site that may Regulate C/EBPβ Binding and Determine Tissue-Specific Expression of the BPI Gene in Piglets. Sci. Rep. 2016, 6, 28506. [Google Scholar] [CrossRef]
- Liu, L.; Wang, J.; Zhao, Q.H.; Zi, C.; Wu, Z.C.; Su, X.M.; Huo, Y.J.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Genetic variation in exon 10 of the BPI gene is associated with Escherichia coli F18 susceptibility in Sutai piglets. Gene 2013, 523, 70–75. [Google Scholar] [CrossRef]
- Liu, Y.; Gan, L.N.; Qin, W.Y.; Sun, S.Y.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Differential expression of Toll-like receptor 4 signaling pathway genes in Escherichia coli F18-resistant and-sensitive Meishan piglets. Pol. J. Vet. Sci. 2016, 19, 303–308. [Google Scholar] [CrossRef]
- Dong, W.H.; Dai, C.H.; Sun, L.; Wang, J.; Sun, S.Y.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Expression of key glycosphingolipid biosynthesis-globo series pathway genes in Escherichia coli F18-resistant and Escherichia coli F18-sensitive piglets. Anim. Genet. 2016, 47, 428–435. [Google Scholar] [CrossRef]
- Dai, C.H.; Gan, L.N.; Qin, W.U.; Zi, C.; Zhu, G.Q.; Wu, S.L.; Bao, W.B. Use of Fluorescence Quantitative Polymerase Chain Reaction (PCR) for the Detection of Escherichia coli Adhesion to Pig Intestinal Epithelial Cells. Pol. J. Vet. Sci. 2016, 19, 619–625. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Preston, A.; Mandrell, R.E.; Gibson, B.W.; Apicella, M.A. The lipooligosaccharides of pathogenic gram-negative bacteria. Crit. Rev. Microbiol. 1996, 22, 139–180. [Google Scholar] [CrossRef] [PubMed]
- Benin, A.M.; Ducher-Suchaux, M.F. Relationship between virulence and adherence of various enterotoxigenic Escherichia coli: Strains to isolated intestinal epithelial cells from Chinese Meishan and European large white pigs. Am. J. Vet. Res. 1991, 52, 45–49. [Google Scholar]
- Boldin, B. Persistence and Spread of Gastro-Intestinal Infections: The case of enterotoxigenic Escherichia coli in piglets. Bull. Math Biol. 2008, 70, 2077–2101. [Google Scholar] [CrossRef] [PubMed]
- Roy, K.; Hilliard, G.M.; Hamilton, D.J.; Luo, J.; Ostmann, M.M.; Fleckenstein, J.M. Enterotoxigenic Escherichia coli EtpA mediates adhesion between flagella and host cells. Nature 2009, 457, 594–598. [Google Scholar] [CrossRef]
- Jiang, S.; Li, F.Q.; Li, X.L.; Wang, L.I.; Zhang, L.; Lu, C.; Zheng, L.; Yan, M.H. Transcriptome analysis of PK-15 cells in innate immune response to porcine deltacoronavirus infection. PLoS ONE 2019, 14, e0223177. [Google Scholar] [CrossRef]
- Frommel, U.; Bohm, A.; Nitschke, J.; Weinreich, J.; Groß, J.; Rödiger, S.; Wex, T.; Ansorge, H.; Zinke, O.; Schröder, C.; et al. Adhesion patterns of commensal and pathogenic Escherichia coli from humans and wild animals on human and porcine epithelial cell lines. Gut. Pathog. 2013, 5, 31. [Google Scholar] [CrossRef][Green Version]
- Meijerink, E.; Neuenschwander, S.; Fries, R.; Dinter, A.; Bertschinger, H.U.; Stranzinger, G.; Vögeli, P. A DNA polymorphism influencing α(1,2) fucosyltransferase activity of the pig FUT1 enzyme determines susceptibility of small intestinal epithelium to Escherichia coli F18 adhesion. Immunogenetics 2000, 52, 129–136. [Google Scholar] [CrossRef]
- Wu, Z.; Feng, H.; Cao, Y.; Huang, Y.; Dai, C.; Wu, S.; Bao, W. New Insight into the Molecular Mechanism of the FUT2 Regulating Escherichia coli F18 Resistance in Weaned Piglets. Int. J. Mol. Sci. 2018, 19, 3301. [Google Scholar] [CrossRef]
- De Nardo, D. Toll-like receptors: Activation, signalling and transcriptional modulation. Cytokine 2015, 74, 181–189. [Google Scholar] [CrossRef]
- Barton, G.M.; Medzhitov, R. Toll-like receptor signaling pathways. Science 2003, 300, 1524–1525. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, A.; Schnare, M.; Chakravortty, D. Of men not mice: Bactericidal/permeability-increasing protein expressed in human macrophages acts as a phagocytic receptor and modulates entry and replication of Gram-negative bacteria. Front. Immunol. 2016, 7, 455. [Google Scholar] [CrossRef] [PubMed]
- Canny, G.; Cario, E.; Lennartsson, A.; Gullberg, U.; Brennan, C.; Levy, O.; Colgan, S.P. Functional and biochemical characterization of epithelial bactericidal/permeability-increasing protein. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G557–G567. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.; Weiss, J.; Carroll, S.F.; Gross, W.L. The endotoxin-binding bactericidal/permeability-increasing protein (BPI): A target antigen of autoantibodies. J. Leukoc. Biol. 2001, 69, 505–512. [Google Scholar] [PubMed]
- Xie, S.; Chen, X.; Qiao, S.L.; Li, R.; Sun, Y.G.; Xia, S.F.; Wang, L.J.; Luo, X.G.; Deng, R.G.; Zhou, E.M.; et al. Identification of the RNA pseudoknot within the 3′ end of the porcine reproductive and respiratory syndrome virus genome as a pathogen-associated molecular pattern to activate antiviral signaling via RIG-I and toll-like receptor 3. J. Virol. 2018, 92, e00097-18. [Google Scholar] [CrossRef]
- Sato, M.; Suemori, H.; Hata, N.; Asagiri, M.; Ogasawara, K.; Nakao, K.; Nakaya, T.; Katsuki, M.; Noguchi, S.; Tanaka, N.; et al. Distinct and Essential Roles of Transcription Factors IRF-3 and IRF-7 in Response to Viruses for IFN-α/β Gene Induction. Immunity 2000, 13, 539–548. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef]
- Lenschow, D.J.; Lai, C.; Frias-Staheli, N.; Giannakopoulos, N.V.; Lutz, A.; Wolff, T.; Osiak, A.; Levine, B.; Schmidt, R.E.; García-Sastre, A.; et al. IFN-stimulated gene 15 functions as a critical antiviral molecule against influenza, herpes, and Sindbis viruses. Proc. Natl. Acad. Sci. USA 2007, 104, 1371–1376. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Product Length (bp) |
|---|---|---|
| BPI TLR4 | F: 5′-ATATCGAATCTGCGCTCCGA-3′ R: 5′-TTGATGCCAACCATTCTGTCC-3′ F: 5′-CAGATAAGCGAGGCCGTCATT-3′ R: 5′-TTGCAGCCCACAAAAAGCA-3′ | 136 113 |
| MyD88 | F: 5′-GTGCCGTCGGATGGTAGT-3′ R: 5′-CAGTGATGAACCGCAGGAT-3′ | 173 |
| CD14 | F: 5′-CCTCAGACTCCGTAATGTG-3′ R: 5′-CCGGGATTGTCAGATAGG-3′ | 180 |
| TNF-α | F: 5′-CGACTCAGTGCCGAGATCAA-3′ R: 5′-CCTGCCCAGATTCAGCAAAG-3′ | 58 |
| IL-1β FUT1 FUT2 GAPDH PILIN | F: 5′-TGATTGTGGCAAAGGAGGA-3′ R: 5′-TTGGGTCATCATCACAGACG-3′ F: 5′-TTTTAAGCCCCCAAACTGCC-3′ R: 5′-TAAATCGACCCCATCAGCCTC-3′ F: 5′-AATCCCTGACCTCACTCCGTG-3′ R: 5′-CGGAACTACAACTGCTGGCC-3′ F: 5′-ACATCATCCCTGCTTCTACTGG-3′ R: 5′-CTCGGACGCCTGCTTCAC-3′ F: 5′-AGGCCGAACCAAAGAAGCAT-3′ R: 5′-TCACCATCAGGGTTTCTGAGT-3′ | 63 126 123 188 117 |
| β-ACTIN | F: 5′-GTCGTACTCCTGCTTGCTGAT-3′ R: 5′-CCTTCTCCTTCCAGATCATCGC-3′ | 119 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, J.; Huang, Y.; Sun, S.; Wu, Z.; Wu, S.; Yin, Z.; Bao, W. The Impact of BPI Expression on Escherichia coli F18 Infection in Porcine Kidney Cells. Animals 2020, 10, 2118. https://doi.org/10.3390/ani10112118
Jin J, Huang Y, Sun S, Wu Z, Wu S, Yin Z, Bao W. The Impact of BPI Expression on Escherichia coli F18 Infection in Porcine Kidney Cells. Animals. 2020; 10(11):2118. https://doi.org/10.3390/ani10112118
Chicago/Turabian StyleJin, Jian, Yanjie Huang, Shouyong Sun, Zhengchang Wu, Shenglong Wu, Zongjun Yin, and Wenbin Bao. 2020. "The Impact of BPI Expression on Escherichia coli F18 Infection in Porcine Kidney Cells" Animals 10, no. 11: 2118. https://doi.org/10.3390/ani10112118
APA StyleJin, J., Huang, Y., Sun, S., Wu, Z., Wu, S., Yin, Z., & Bao, W. (2020). The Impact of BPI Expression on Escherichia coli F18 Infection in Porcine Kidney Cells. Animals, 10(11), 2118. https://doi.org/10.3390/ani10112118