Urban Hedgehog Behavioural Responses to Temporary Habitat Disturbance versus Permanent Fragmentation

 ,
,  , , and
, , and 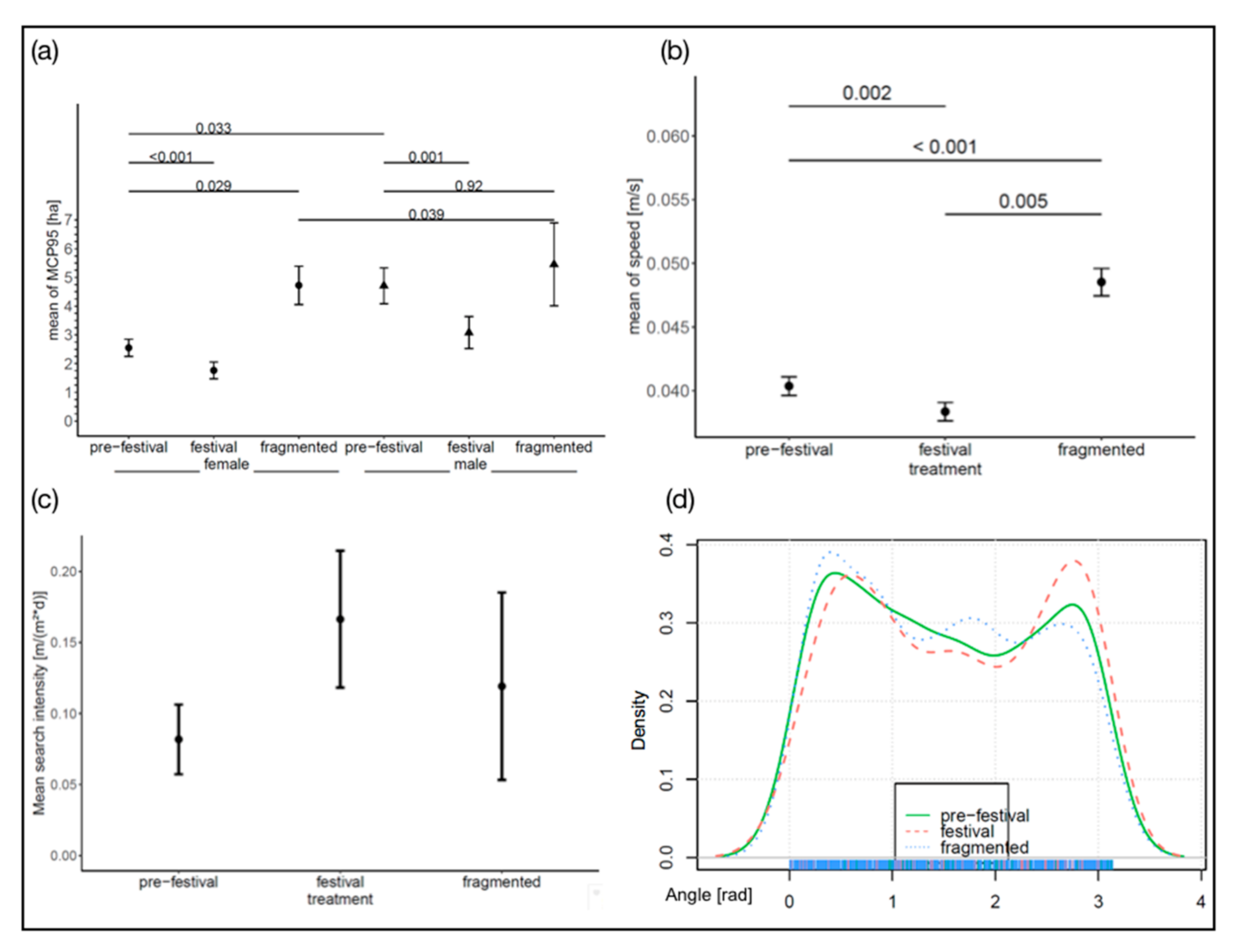{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
- (1)
- Regarding space use, we predict that hedgehogs avoid transient disturbed areas in their habitat, by either leaving the area or adjusting movement patterns: (1a) We predict avoidance of the disturbed (festival) area by shifting the centre of the nightly used area. (1b) Additionally, we predict a decreased size of nightly home ranges in or close to the festival area due to avoidance of the disturbed (festival) area.
- (2)
- We predict that hedgehogs adjust their movement behaviour. The animals now have to look for the same amount of food in a potentially less favourable and/or smaller area and thus foraging effort may have to be increased. We, therefore, predict an increase in search intensity, greater turning angles and slower speed under disturbance.
- (3)
- The general levels of activity will be reduced due to increased vigilance behaviour which in hedgehogs is characterised by immobility (little activity).
- (4)
- We predict that high levels of disturbance during the festival induces females and males to switch their nests more often and the number of days spent in the same nest decreases.
- (1)
- In urban areas, or areas with predation risk, hedgehog movements are strongly associated with linear structures [35], fragmentation will increase the area of space that is of no interest to hedgehogs and thus increase the distances they have to cover. Thus, in a highly fragmented park, the home range area would be bigger than in the low-fragmented park.
- (2)
- As fragmentation is likely to increase distances for commuting between favourable food patches, movement characteristics should change. We expect a faster speed, a larger number of smaller turning angels and a lower search intensity than in a low-disturbed, low-fragmented habitat.
- (3)
- Fragmentation could influence nesting behaviour in two ways: animals either have to change their nests more frequently to be closer to favourable food patches, or extend their stay in nests if they are close to favourable food patches. During disturbances, we predict a more frequent nest change than in undisturbed areas or at undisturbed time.
2. Materials and Methods
2.1. Study Areas and Study Design
2.2. Hedgehog Capture and Logger Attachment
2.3. Logger Sampling Setup
2.4. Nesting Behaviour Monitoring
2.5. Analyses
2.5.1. GPS Data
2.5.2. ACC/Acceleration Data
Behaviour Prediction and Budget
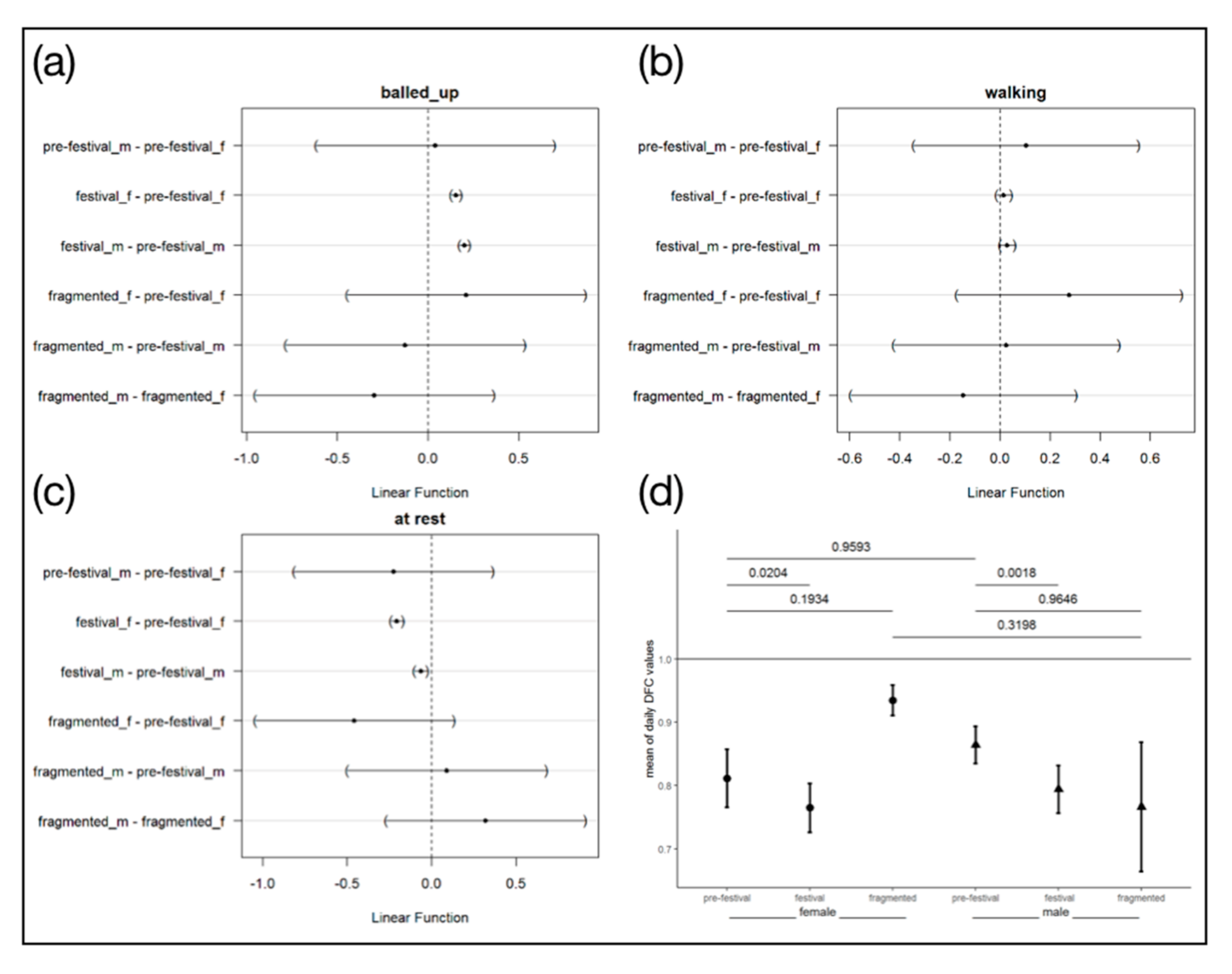
Stress Detection via the Degree of Functional Coupling (DFC)
2.5.3. Nesting Behaviour
3. Results
3.1. Home Range Size
3.2. Movement Speed and Turning Angle
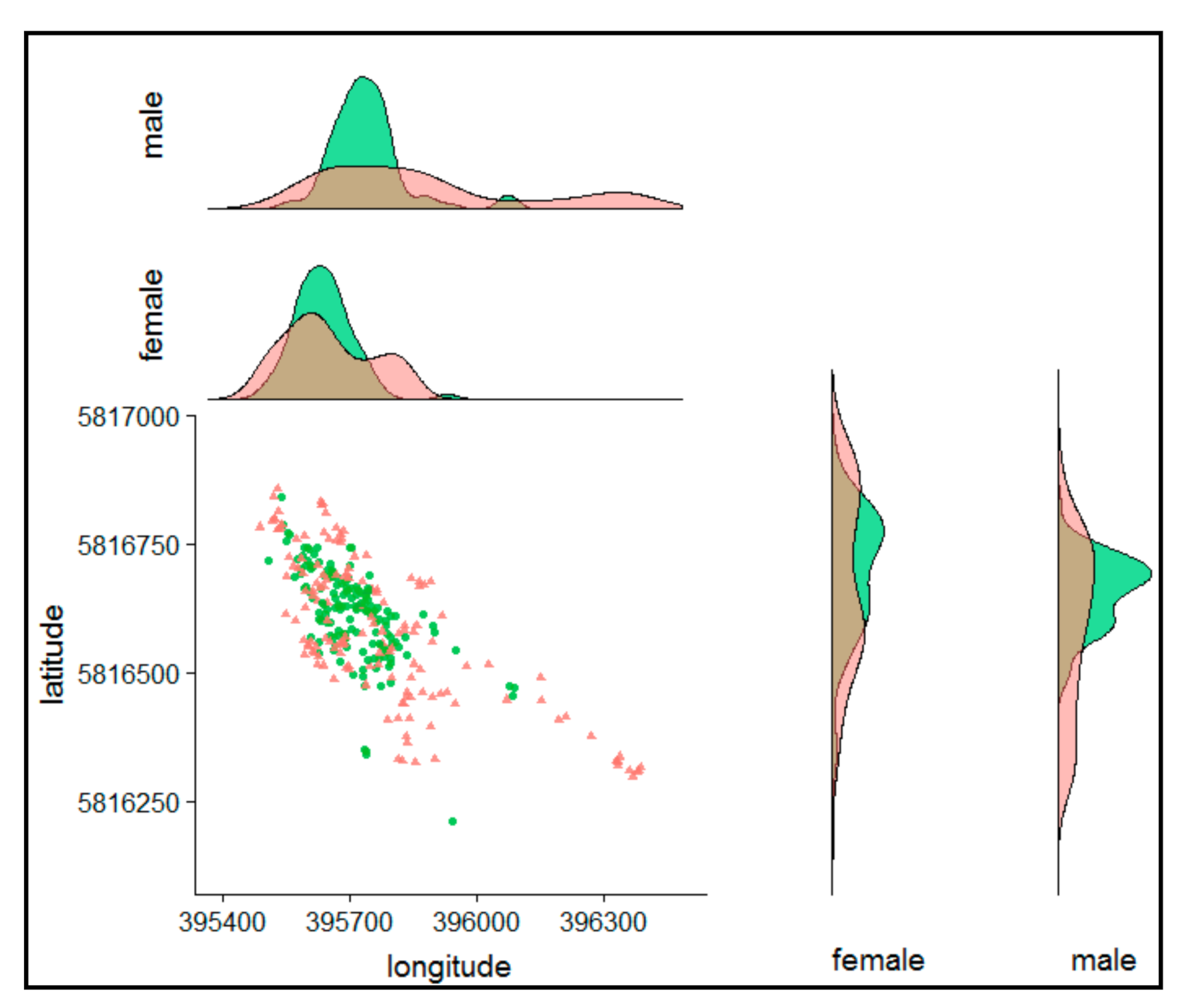
3.3. The Centre of the Nightly Home Range
3.4. Behaviour Parameters
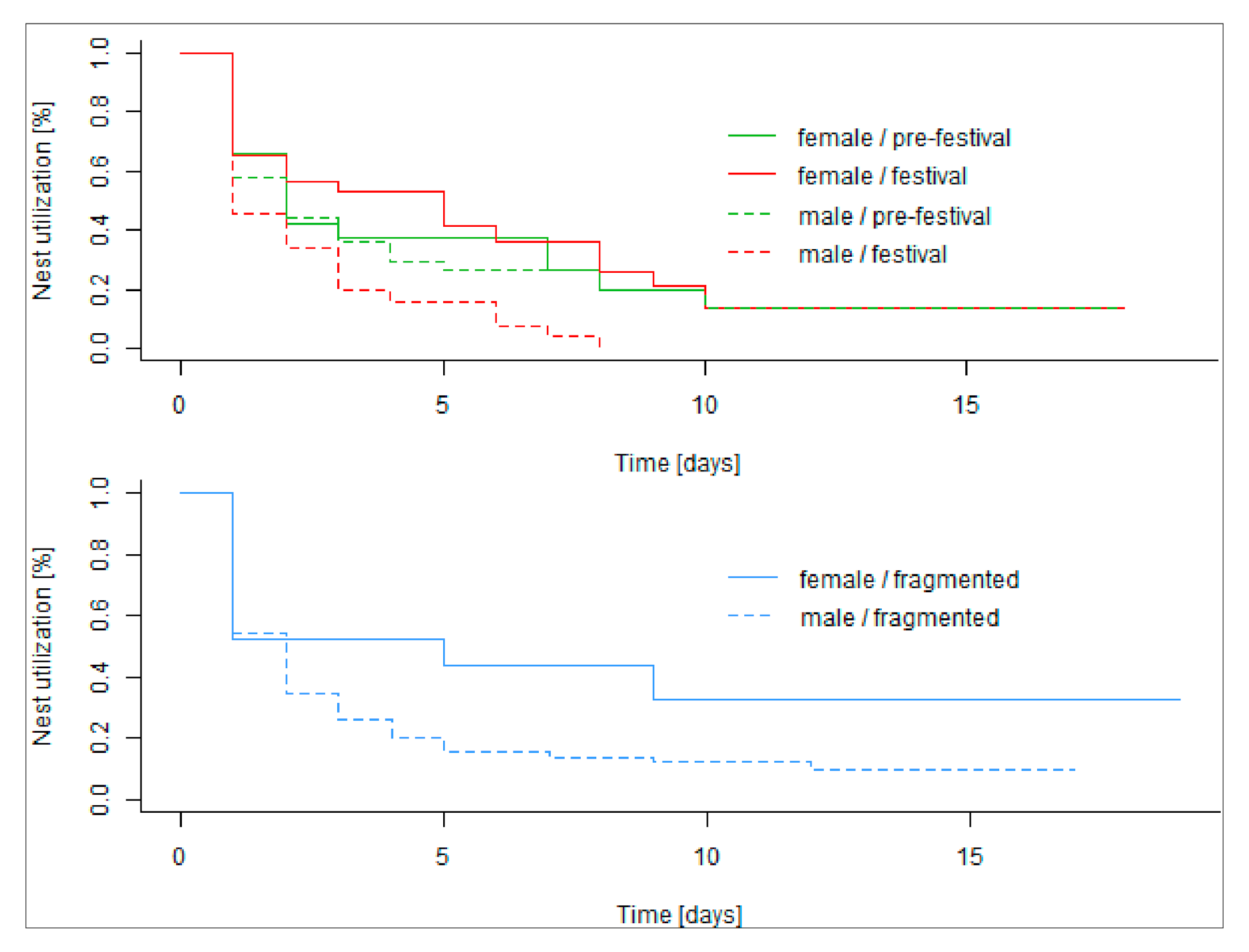
3.5. Nesting Behaviour
4. Discussion
4.1. Home Range Sizes
4.2. Movement Speed and Turning Angle
4.3. Centre of Nightly Home Range
4.4. Behaviour Parameter
4.5. Nesting Behaviour
4.6. Limitations and Outlook
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rykiel, E.J. Towards a definition of disturbance. Aust. J. Ecol. 1985, 10, 361–365. [Google Scholar] [CrossRef]
- Acevedo-Whitehouse, K.; Duffus, A.L.J. Effects of environmental change on wildlife health. Phil. Trans. R. Soc. B 2009, 364, 3429–3438. [Google Scholar] [CrossRef]
- Walker, L.R. The Biology of Disturbed Habitats; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- McDonnell, M.J.; Hahs, A.K. Adaptation and adaptedness of organisms to urban environments. Ann. Rev. Ecol. Evol. Syst. 2015, 46, 261–280. [Google Scholar] [CrossRef]
- Bleicher, S.S. The landscape of fear conceptual framework: Definition and review of current applications and misuses. PeerJ 2017, 5, e3772. [Google Scholar] [CrossRef] [PubMed]
- Stillfried, M.; Gras, P.; Börner, K.; Göritz, F.; Painer, J.; Röllig, K.; Wenzler, M.; Hofer, H.; Ortmann, S.; Kramer-Schadt, S. Secrets of success in a landscape of fear: Urban wild boar adjust risk perception and tolerate disturbance. Front. Ecol. Evol. 2017, 5, 157. [Google Scholar] [CrossRef]
- Wong, B.B.M.; Candolin, U. Behavioral responses to changing environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef]
- Hastings, A.; Abbott, K.C.; Cuddington, K.; Francis, T.; Gellner, G.; Lai, Y.C.; Morozov, A.; Petrovskii, S.; Scranton, K.; Zeeman, M.L. Transient phenomena in ecology. Science 2018, 361. [Google Scholar] [CrossRef]
- Doncaster, C.P.; Dickman, C.R. The ecology of small mammals in urban habitats. I. Population in a patchy environment. J. Anim. Ecol. 1987, 56, 629–640. [Google Scholar]
- Wilcove, D.S.; McLellan, C.H.; Dobson, A.P. Habitat fragmentation in the temperate zone. In Conservation Biology—The Science of Scarcity and Diversity; Soulé, M.E., Ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 1986; pp. 237–256. [Google Scholar]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Ann. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Baker, P.J.; Harris, S. Urban mammals: What does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mammal Rev. 2007, 37, 297–315. [Google Scholar] [CrossRef]
- Banks, S.C.; Piggott, M.P.; Stow, A.J.; Taylor, A.C. Sex and sociality in a disconnected world: A review of the impacts of habitat fragmentation on animal social interactions. Can. J. Zool. 2007, 85, 1065–1079. [Google Scholar] [CrossRef]
- Shepard, D.B.; Kuhns, A.R.; Dreslik, M.J.; Phillips, C.A. Roads as barriers to animal movement in fragmented landscapes. Anim. Conserv. 2008, 11, 288–296. [Google Scholar] [CrossRef]
- Lowry, H.; Lill, A.; Wong, B.B.M.M. Behavioural responses of wildlife to urban environments. Biol. Rev. 2013, 88, 537–549. [Google Scholar] [CrossRef]
- Hertel, A.G.; Steyaert, S.M.J.G.; Zedrosser, A.; Mysterud, A.; Lodberg-Holm, H.K.; Gelink, H.W.; Kindberg, J.; Swenson, J.E. Bears and berries: Species-specific selective foraging on a patchily distributed food resource in a human-altered landscape. Behav. Ecol. Sociobiol. 2016, 70, 831–842. [Google Scholar] [CrossRef] [PubMed]
- Hertel, A.G.; Swenson, J.E.; Bischof, R. A case for considering individual variation in diel activity patterns. Behav. Ecol. 2017, 28, 1524–1531. [Google Scholar] [CrossRef]
- Soanes, K.; Taylor, A.C.; Sunnucks, P.; Vesk, P.A.; Cesarini, S.; van der Ree, R. Evaluating the success of wildlife crossing structures using genetic approaches and an experimental design: Lessons from a gliding mammal. J. Appl. Ecol. 2018, 55, 129–138. [Google Scholar] [CrossRef]
- Reeve, N. Hedgehogs; T. & A.D. Poyser: London, UK, 1994. [Google Scholar]
- Morris, P. Hedgehogs; Whittet Books Ltd.: Stansted, UK, 2014. [Google Scholar]
- Pettett, C.E.; Johnson, P.J.; Moorhouse, T.P.; Macdonald, D.W. National predictors of hedgehog Erinaceus europaeus distribution and decline in Britain. Mammal Rev. 2018, 48, 1–6. [Google Scholar] [CrossRef]
- Krange, M. Change in the Occurrence of the West European Hedgehog (Erinaceus europaeus) in Western Sweden During 1950–2010; Karlstad University: Karlstad, Sweden, 2015. [Google Scholar]
- Holsbeek, L.; Rodts, J.; Muyldermans, S. Hedgehog and other animal traffic victims in Belgium: Results of a countrywide survey. Lutra 1999, 42, 111–119. [Google Scholar]
- Reichholf, J.H. Starker Rückgang der Häufigkeit überfahrener Igel Erinaceus europaeus in Südostbayern und seine Ursachen. Mitt. Zool. Ges. Braunau 2015, 11, 309–314. [Google Scholar]
- Hof, A.R.; Bright, P.W. The value of green-spaces in built-up areas for western hedgehogs. Lutra 2009, 52, 69–82. [Google Scholar]
- Williams, B.M.; Baker, P.J.; Thomas, E.; Wilson, G.; Judge, J.; Yarnell, R.W. Reduced occupancy of hedgehogs (Erinaceus europaeus) in rural England and Wales: The influence of habitat and an asymmetric intra-guild predator. Sci. Rep. 2018, 8, 12156. [Google Scholar] [CrossRef]
- Wright, P.G.; Coomber, F.G.; Bellamy, C.C.; Perkins, S.E.; Mathews, F. Predicting hedgehog mortality risks on British roads using habitat suitability modelling. PeerJ 2020, 7, e8154. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. The value of agri-environment schemes for macro-invertebrate feeders: Hedgehogs on arable farms in Britain. Anim. Conserv. 2010, 13, 467–473. [Google Scholar] [CrossRef]
- Becher, S.A.; Griffiths, R. Genetic differentiation among local populations of the European hedgehog (Erinaceus europaeus) in mosaic habitats. Mol. Ecol. 1998, 7, 1599–1604. [Google Scholar] [CrossRef]
- Rondinini, C.; Doncaster, C. Roads as barriers to movement for hedgehogs. Funct. Ecol. 2002, 16, 504–509. [Google Scholar] [CrossRef]
- Braaker, S.; Kormann, U.; Bontadina, F.; Obrist, M.K. Prediction of genetic connectivity in urban ecosystems by combining detailed movement data, genetic data and multi-path modelling. Landsc. Urban Plan. 2017, 160, 107–114. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Yashiro, E.; Sverrisdóttir, E.; Nielsen, K.L.; Lukassen, M.B.; Nielsen, J.L.; Asp, T.; Pertoldi, C. Applying the GBS technique for the genomic characterization of a Danish population of European hedgehogs (Erinaceus europaeus). Gen. Biodiv. J. 2019, 3, 78–86. [Google Scholar]
- Hof, A.R.; Allen, A.M.; Bright, P.W. Investigating the role of the Eurasian Badger (Meles meles) in the nationwide distribution of the Western European hedgehog (Erinaceus europaeus) in England. Animals 2019, 9, 759. [Google Scholar] [CrossRef]
- Braaker, S.; Moretti, M.; Boesch, R.; Ghazoul, J.; Obrist, M.K.; Bontadina, F. Assessing habitat connectivity for ground-dwelling animals in an urban environment. Ecol. Appl. 2014, 24, 1583–1595. [Google Scholar] [CrossRef]
- Dowding, C.V.; Harris, S.; Poulton, S.; Baker, P.J. Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward. Anim. Behav. 2010, 80, 13–21. [Google Scholar] [CrossRef]
- Reeve, N.J.; Morris, P.A. Construction and use of summer nests by the hedgehog Erinaceus europaeus. Mammalia 1985, 49, 187–194. [Google Scholar] [CrossRef]
- Haigh, A.; O’Riordan, R.M.; Butler, F. Nesting behaviour and seasonal body mass changes in a rural Irish population of the Western hedgehog (Erinaceus europaeus). Acta Theriol. 2012, 57, 321–331. [Google Scholar] [CrossRef]
- Mori, E.; Menchetti, M.; Bertolino, S.; Mazza, G.; Ancillotto, L. Reappraisal of an old cheap method for marking the European hedgehog. Mammal Res. 2015, 60, 189–193. [Google Scholar] [CrossRef]
- Reeve, N.J.; Bowen, C.; Gurnell, J. An Improved Identification Marking Method for Hedgehogs; The Mammal Society: Southampton, UK, 2019. [Google Scholar]
- Barthel, L.M.F.; Hofer, H.; Berger, A. An easy, flexible solution to attach devices to hedgehogs (Erinaceus europaeus) enables long-term high-resolution studies. Ecol. Evol. 2019, 9, 672–679. [Google Scholar] [CrossRef]
- Hofer, H.; East, M.L. Biological conservation and stress. Adv. Study Behav. 1998, 21, 405–525. [Google Scholar] [CrossRef]
- Sikes, R.S.; Gannon, W.L.; The Animal Care and Use Committee of the American Society of Mammalogists. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Calenge, C. The package adehabitat for the R software: Tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 1035. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. 2018. Available online: https://cran.r-project.org/package=nlme (accessed on 25 March 2019).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed.; Sage: Thousand Oaks, CA, USA, 2011; Available online: http://socserv.socsci.mcmaster.ca/jfox/Books/Companion (accessed on 25 March 2019).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Rousset, F.; Ferdy, J.-B. Testing environmental and genetic effects in the presence of spatial autocorrelation. Ecography 2014, 37, 781–790. [Google Scholar] [CrossRef]
- Hijmans, R.J. Geosphere: Spherical Trigonometry. 2017. Available online: https://cran.r-project.org/package=geosphere (accessed on 25 March 2019).
- Michelot, T.; Langrock, R.; Patterson, T.A. moveHMM: An R package for the statistical modelling of animal movement data using hidden Markov models. Methods Ecol. Evol. 2016, 7, 1308–1315. [Google Scholar] [CrossRef]
- Rast, W.; Barthel, L.M.F.; Berger, A. Music festival makes Hedgehogs move: How individuals cope behaviorally in response to human-induced stressors. Animals 2019, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Scheibe, K.M.; Berger, A.; Langbein, J.; Streich, W.J.; Eichhorn, K. Comparative analysis of ultradian and circadian behavioural rhythms for diagnosis of biorhythmic state of animals. Biol. Rhyth. Res. 1999, 30, 216–233. [Google Scholar] [CrossRef]
- Berger, A.; Scheibe, K.-M.; Michaelis, S.; Streich, W.J. Evaluation of living conditions of free-ranging animals by automated chronobiological analysis of behavior. BRMIC 2003, 35, 458–466. [Google Scholar] [CrossRef]
- Berger, A. Activity patterns, chronobiology and the assessment of stress and welfare in zoo and wild animals. Int. Zoo Yearb. 2011, 45, 80–90. [Google Scholar] [CrossRef]
- Therneau, T.M. A Package for Survival Analysis in R. 2015. Available online: https://cran.r-project.org/package=survival (accessed on 1 March 2019).
- Von der Lippe, M.; Buchholz, S.; Hiller, A.; Seitz, B.; Kowarik, I. CityScapeLab Berlin: A research platform for untangling urbanization effects on biodiversity. Sustainability 2020, 12, 2565. [Google Scholar] [CrossRef]
- Buchholz, S.; Hannig, K.; Möller, M.; Schirmel, J. Reducing management intensity and isolation as promising tools to enhance ground-dwelling arthropod diversity in urban grasslands. Urban Ecosyst. 2018, 21, 1139–1149. [Google Scholar] [CrossRef]
- Seress, G.; Lipovits, Á.; Bókony, V.; Czúni, L. Quantifying the urban gradient: A practical method for broad measurements. Landsc. Urban Plan. 2014, 131, 42–50. [Google Scholar] [CrossRef]
- Harris, S. The food of suburban foxes (Vulpes vulpes), with special reference to London. Mammal Rev. 2008, 11, 151–168. [Google Scholar] [CrossRef]
- Luniak, M. Synurbization—Adaptation of animal wildlife to urban development. In Proceedings of the 4th International Symposium on Urban Wildlife Conservation, Tucson, AZ, USA, 1–5 May 1999. [Google Scholar]
- Schoenfeld, M.; Yom-Tov, Y. The biology of two species of hedgehogs, Erinaceus europaeus concolor and Hemiechinus auritus aegyptius, in Israel. Mammalia 1985, 49, 339–354. [Google Scholar] [CrossRef]
- Boitani, L.; Reggiani, G. Movements and activity patterns of Hedgehogs (Erinaceus europaeus) in Mediterranean coastal habitats. Z. Säugetierkunde 1984, 49, 193–206. [Google Scholar]
- Kristiansson, H. Ecology of a Hedgehog Erinaceus europaeus Population in Southern Sweden; University of Lund: Lund, Sweden, 1984. [Google Scholar]
- Morris, P.A. A study of home range and movements in the hedgehog (Erinaceus europaeus). J. Zool. 1988, 214, 433–449. [Google Scholar] [CrossRef]
- Reeve, N.J. The survival and welfare of hedgehogs (Erinaceus europaeus) after release back into the wild. Anim. Welf. 1997, 7, 189–202. [Google Scholar]
- Rautio, A.; Valtonen, A.; Kunnasranta, M. The effects of sex and season on home range in European hedgehogs at the northern edge of the species range. Ann. Zool. Fenn. 2013, 50, 107–123. [Google Scholar] [CrossRef]
- Baker, M.A.A.; Reeve, N.; Conkey, A.A.T.; Macdonald, D.W.; Yamaguchi, N. Hedgehogs on the move: Testing the effects of land use change on home range size and movement patterns of free-ranging Ethiopian hedgehogs. PLoS ONE 2017, 12, e0180826. [Google Scholar] [CrossRef]
- Pettett, C.E.; Johnson, P.J.; Moorhouse, T.P.; Hambly, C.; Speakman, J.R.; Macdonald, D.W. Daily energy expenditure in the face of predation: Hedgehog energetics in rural landscapes. J. Exp. Biol. 2017, 220, 460–468. [Google Scholar] [CrossRef]
- Tucker, M.A.; Böhning-Gaese, K.; Fagan, W.F.; Fryxell, J.M.; van Moorter, B.; Alberts, S.C.; Ali, A.H.; Allen, A.M.; Attias, N.; Avgar, T.; et al. Moving in the Anthropocene: Global reductions in terrestrial mammalian movements. Science 2018, 359, 466–469. [Google Scholar] [CrossRef]
- Bontadina, F. Strassenüberquerungen von Igeln Zürich, Institute of Zoology; University of Zurich: Zurich, Switzerland, 1991. [Google Scholar]
- Zingg, R. Aktivitaet Habitat- und Raumnutzung von Igeln (Erinaceus Europaeus) in Einem Ländlichen Siedlungsgebiet. Ph.D. Thesis, University of Zurich, Zurich, Switzerland, 1994. [Google Scholar]
- Doncaster, C.P.; Krebs, J.R. The wider countryside-principles underlying the responses of mammals to heterogeneous environments. Mammal Rev. 1993, 23, 113–120. [Google Scholar] [CrossRef]
- Phillips, S. Aversive behaviour by koalas (Phascolarctos cinereus) during the course of a music festival in northern New South Wales, Australia. Aust. Mammal. 2016, 38, 158. [Google Scholar] [CrossRef]
- Fitzgibbon, S.I.; Gillett, A.K.; Barth, B.J.; Taylor, B.; Ellis, W.A. Do koalas really get the blues? Critique of aversive behaviour by koalas (Phascolarctos cinereus) during the course of a music festival in northern New South Wales, Australia. Aust. Mammal 2017, 39, 108. [Google Scholar] [CrossRef]
- Doncaster, C.P.; Rondinini, C.; Johnson, P.C.D. Field test for environmental correlates of dispersal in hedgehogs Erinaceus europaeus. J. Anim. Ecol. 2001, 70, 33–46. [Google Scholar] [CrossRef]
- Lott, D.F. Intraspecific Variation in the Social Systems of Wild Vertebrates; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Krone, O.; Berger, A.; Schulte, R. Recording movement and activity pattern of a White-tailed Sea Eagle (Haliaeetus albicilla) by a GPS datalogger. J. Ornithol. 2009, 150, 273–280. [Google Scholar] [CrossRef]
- Langbein, J.; Scheibe, K.-M.; Eichhorn, K. Investigations on periparturient behaviour in free-ranging mouflon sheep (Ovis orientalis musimon). J. Zool. 1998, 244, 553–561. [Google Scholar] [CrossRef]
- Santini, L.; González-Suárez, M.; Russo, D.; Gonzalez-Voyer, A.; von Hardenberg, A.; Ancillotto, L. One strategy does not fit all: Determinants of urban adaptation in mammals. Ecol. Lett. 2018, 22, 365–376. [Google Scholar] [CrossRef]
- Kerley, G.I.H.; Kowalczyk, R.; Cromsigt, J.P.G.M. Conservation implications of the refugee species concept and the European bison: King of the forest or refugee in a marginal habitat? Ecography 2012, 35, 519–529. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Hickler, T.; Olofsson, J.; Schurgers, G.; Radeloff, V.C. Refugee species: Which historic baseline should inform conservation planning? Divers. Distrib. 2012, 18, 1258–1261. [Google Scholar] [CrossRef]
- Kays, R.; Crofoot, M.C.; Jetz, W.; Wikelski, M. Terrestrial animal tracking as an eye on life and planet. Science 2015, 348, aaa2478. [Google Scholar] [CrossRef]
- Hebblewhite, M.; Haydon, D.T. Distinguishing technology from biology: A critical review of the use of GPS telemetry data in ecology. Philos. Trans. R. Soc. B 2010, 365, 2303–2312. [Google Scholar] [CrossRef]
- Shamoun-Baranes, J.; Bom, R.; van Loon, E.E.; Ens, B.J.; Oosterbeek, K.; Bouten, W. From sensor data to animal behaviour: An oystercatcher example. PLoS ONE 2012, 7, e37997. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berger, A.; Barthel, L.M.F.; Rast, W.; Hofer, H.; Gras, P. Urban Hedgehog Behavioural Responses to Temporary Habitat Disturbance versus Permanent Fragmentation. Animals 2020, 10, 2109. https://doi.org/10.3390/ani10112109
Berger A, Barthel LMF, Rast W, Hofer H, Gras P. Urban Hedgehog Behavioural Responses to Temporary Habitat Disturbance versus Permanent Fragmentation. Animals. 2020; 10(11):2109. https://doi.org/10.3390/ani10112109
Chicago/Turabian StyleBerger, Anne, Leon M. F. Barthel, Wanja Rast, Heribert Hofer, and Pierre Gras. 2020. "Urban Hedgehog Behavioural Responses to Temporary Habitat Disturbance versus Permanent Fragmentation" Animals 10, no. 11: 2109. https://doi.org/10.3390/ani10112109
APA StyleBerger, A., Barthel, L. M. F., Rast, W., Hofer, H., & Gras, P. (2020). Urban Hedgehog Behavioural Responses to Temporary Habitat Disturbance versus Permanent Fragmentation. Animals, 10(11), 2109. https://doi.org/10.3390/ani10112109