Influence of Season and Liquid Storage at 16 °C on Beni Arouss Bucks’ Semen Quality

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management and Semen Collection
2.2. Semen Quality Assessment and Processing
2.3. Statistical Analysis
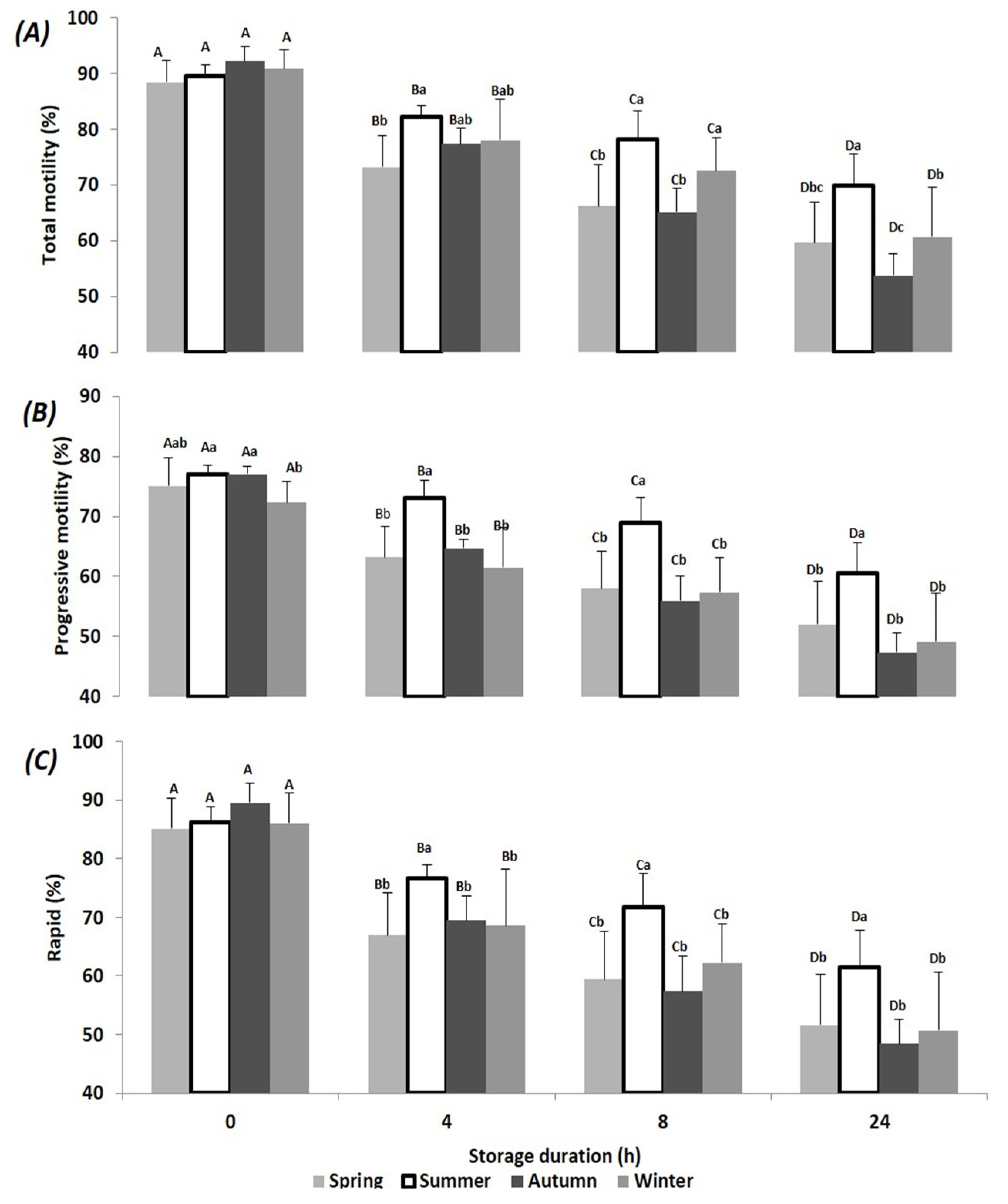
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chentouf, M.; Bister, J.L.; Boulanouar, B. Reproduction characteristics of North Moroccan indigenous goats. Small Rumin. Res. 2011, 98, 185–188. [Google Scholar] [CrossRef]
- Chentouf, M.; Ben Bati, M.; Zantar, S.; Boulanouar, B.; Bister, J.L. Evaluation des performances des élevages caprins extensifs dans le nord du Maroc. Options Méditerr. 2006, 70, 87–93. [Google Scholar]
- El Otmani, S.; Hilal, B.; Chentouf, M. Milk production and composition of ‘Beni Arousse’ North Moroccan local goat. In Proceedings of the 39th ICAR Session, Berlin, Germany, 19–23 May 2014. [Google Scholar]
- Leboeuf, B.; Guillouet, P.; Batellier, F.; Bernelas, D.; Bonne, J.L.; Forgerit, Y.; Renaud, R.; Magistrini, M. Effect of native phosphocaseinate on the in vitro preservation of fresh semen. Theriogenology 2003, 60, 867–877. [Google Scholar] [CrossRef]
- Paulenz, H.; Soderquist, L.; Adnoy, T.; Soltun, K.; Sæther, P.A.; Fjellsøy, K.R.; Andersen Berg, K. Effect of cervical and vaginal insemination with liquid semen stored at room temperature on fertility of goats. Anim. Reprod. Sci. 2005, 86, 109–117. [Google Scholar] [CrossRef]
- Luo, J.; Wang, W.; Sun, S. Research advances in reproduction for dairy goats. Asian-Australas. J. Anim. Sci. 2019, 32, 1284–1295. [Google Scholar] [CrossRef]
- El Kadili, S.; Raes, M.; Bister, J.L.; Archa, B.; Chentouf, M.; Kirschvink, N. Effect of season on sexual behavior, testicular measurements and seminal characteristics in “Beni arouss” North Moroccan bucks. Anim. Reprod. Sci. 2019, 201, 41–54. [Google Scholar] [CrossRef]
- Benmoula, A.; Badi, A.; El fadili, M.; El Khalil, K.; Allai, L.; El Hilali, A.; El Amiri, B. Effect of season on scrotal circumference, semen characteristics, seminal plasma composition and spermatozoa motility during liquid storage in INRA180 rams. Anim. Reprod. Sci. 2017, 180, 17–22. [Google Scholar] [CrossRef]
- D’Alessandro, A.G.; Martemucci, G. Evaluation of seasonal variations of semen freezability in Leccese ram. Anim. Reprod. Sci. 2003, 79, 93–102. [Google Scholar] [CrossRef]
- Wang, W.; Luo, J.; Sun, S.; Xi, L.; Gao, Q.; Haile, A.B.; Shi, H.; Zhang, W.; Shi, H. The Effect of Season on Spermatozoa Motility, Plasma Membrane and Acrosome Integrity in Fresh and Frozen–Thawed Semen from Xinong Saanen Bucks. Reprod. Domest. Anim. 2015, 50, 23–28. [Google Scholar] [CrossRef]
- Gallego-Calvo, L.; Carolina Gatica, M.; Santiago-Moreno, J.; Guzman, J.L.; Zarazaga, L.A. Seasonal changes in reproductive activity, sperm variables and sperm freezability in Blanca Andaluza bucks. Span. J. Agric. Res. 2015, 13, 1–10. [Google Scholar] [CrossRef]
- INRA. Alimentation Des Bovins, Ovins et Caprins; Jarrige, E.R., Ed.; INRA: Paris, France, 1988; pp. 151–153. [Google Scholar]
- Hayden, S.S.; Blanchard, T.L.; Brinsko, S.P.; Varner, D.D.; Hinrichs, K.; Love, C.C. The “dilution effect” in stallion sperm. Theriogenology 2015, 83, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.; Maxwell, W.M.C. Salamon’s Artificial Insemination of Sheep and Goats; Butterworths: Sydney, Australia, 1987; pp. 107–141. [Google Scholar]
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical Analysis of Repeated Measures Data Using SAS Procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, S.T. CASA-practical aspects. J. Androl. 2000, 21, 515–524. [Google Scholar] [PubMed]
- Verstegen, J.; Iguer-Ouada, M.; Onclin, K. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology 2002, 57, 149–179. [Google Scholar] [CrossRef]
- Gòdia, M.; Castelló, A.; Rocco, M.; Cabrera, B.; Rodríguez-Gil, J.E.; Balasch, S.; Lewis, C.; Sánchez, A.; Clop, A. Identification of circular RNAs in porcine sperm and evaluation of their relation to sperm motility. Sci. Rep. 2020, 10, 7985. [Google Scholar] [CrossRef]
- Kathiravan, P.; Kalatharan, J.; John Edwin, M.; Veerapandian, C. Computer automated motion analysis of crossbred bull spermatozoa and its relationship with in vitro fertility in zona-free hamster oocytes. Anim. Reprod. Sci. 2008, 104, 9–17. [Google Scholar] [CrossRef]
- Del Olmo, E.; Bisbal, A.; Maroto-Morales, A.; Garcia-Alvarez, O.; Ramon, M.; Jimenez-Rabadan, P.; Martinez-Pastor, F.; Soler, A.J.; Garde, J.J.; Fernandez-Santos, M.R. Fertility of cryopreserved ovine semen is determined by sperm velocity. Anim. Reprod. Sci. 2013, 138, 102–109. [Google Scholar] [CrossRef]
- Buchelly Imbachí, F.; Zalazar, L.; Pastore, J.I.; Greco, M.B.; Iniesta-Cuerda, M.; Garde, J.J.; Soler, A.J.; Ballarin, V.; Cesari, A. Objective evaluation of ram and buck sperm motility by using a novel sperm tracker software. Reproduction 2018, 156, 11–21. [Google Scholar] [CrossRef]
- Fernandez-Santos, M.R.; Soler, A.J.; Ramon, M.; Ros-Santaella, J.L.; Maroto-Morales, A.; Garcia-Alvarez, O.; Bisbal, A.; Garde, J.J.; Coloma, M.A.; Santiago-Moreno, J. Effect of post-mortem time on post-thaw characteristics of Spanish ibex (Capra pyrenaica) spermatozoa. Anim. Reprod. Sci. 2011, 129, 56–66. [Google Scholar] [CrossRef]
- Chentouf, M.; Boulanouar, B.; Bister, J.L. Performances de production et physiologie de la reproduction de la chèvre du Nord du Maroc. In L’élevage Caprin au Nord du Maroc; Chentouf, M., Boulanouar, B., Bister, J.L., Eds.; INRA-Editions Press: Rabat, Morocco, 2014; pp. 47–48. [Google Scholar]
- O’Hara, L.; Hanrahan, J.P.; Richardson, L.; Donovan, A.; Fair, S.; Evans, A.C.; Lonergan, P. Effect of storage duration, storage temperature, and diluent on the viability and fertility of fresh ram sperm. Theriogenology 2010, 73, 541–549. [Google Scholar] [CrossRef]
- Sadeghi, S.; Del Gallego, R.; García-Colomer, B.; Gómez, E.A.; Yániz, J.L.; Gosálvez, J.; López-Fernández, C.; Silvestre, M.A. Effect of Sperm Concentration and Storage Temperature on Goat Spermatozoa during Liquid Storage. Biology 2020, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Tuli, R.K.; Holtz, W. Effect of season on the freezability of Boer goat semen in the northern temperate zone. Theriogenology 1995, 43, 1359–1363. [Google Scholar] [CrossRef]
- D’Alessandro, A.G.; Martemucci, G. Post-thaw survival and acrosome integrity of spermatozoa of Leccese rams frozen in different seasons with a milk-egg yolk extender. Ital. J. Anim. Sci. 2005, 4, 139–148. [Google Scholar] [CrossRef]
- Barranco, I.; Ortega, M.D.; Martinez-Alborcia, M.J.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Season of ejaculate collection influences the freezability of boar spermatozoa. Cryobiology 2013, 67, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Bromfield, J. A role for seminal plasma in modulating pregnancy outcomes in domestic species. Reproduction 2016, 152, 223–232. [Google Scholar] [CrossRef]
- Nasrin, S.J.; Calogero, S. Seminal Plasma: An Essential Attribute to Spermatozoa. J. Androl. 2012, 33, 536–551. [Google Scholar]
- Höfner, L.; Luther, A.M.; Waberski, D. The role of seminal plasma in the liquid storage of spermatozoa. Anim. Reprod. Sci. 2020, 24, 106290. [Google Scholar] [CrossRef]
- Druart, X.; Rickard, J.P.; Mactier, S.; Kohnke, P.L.; Kershaw-Young, C.M.; Bathgate, R.; Gibb, Z.; Crossett, B.; Tsikis, G.; Labas, V.; et al. Proteomic characterization and cross species comparison of mammalian seminal plasma. J. Proteom. 2013, 91, 13–22. [Google Scholar] [CrossRef]
- Soleilhavoup, C.; Tsikis, G.; Labas, V.; Harichaux, G.; Kohnke, P.L.; Dacheux, J.L.; Guérin, Y.; Gatti, J.L.; de Graaf, S.P.; Druart, X. Ram seminal plasma proteome and its impact on liquid preservation of spermatozoa. J. Proteom. 2014, 109, 245–260. [Google Scholar] [CrossRef]
- Leahy, T.; Rickard, J.P.; Bernecic, N.C.; Druart, X.; de Graaf, S.P. Ram seminal plasma and its functional proteomic assessment. Reproduction 2019, 157, 243–256. [Google Scholar] [CrossRef]
- Perez-Pe, R.; Cebrian-Perez, J.A.; Muino-Blanco, T. Semen plasma proteins prevent cold-shock membrane damage to ram spermatozoa. Theriogenology 2001, 56, 425–434. [Google Scholar] [CrossRef]
- Barrios, B.; Fernández-Juan, M.; Muiño-Blanco, T.; Cebrián-Pérez, J. Immunocytochemical localization and biochemical characterization of two seminal plasma proteins that protect ram spermatozoa against cold shock. J. Androl. 2005, 26, 40–47. [Google Scholar] [CrossRef]
- Bernardini, A.; Hozbor, F.; Sanchez, E.; Fornés, M.W.; Alberio, R.H.; Cesari, A. Conserved ram seminal plasma proteins bind to the sperm membrane and repair cryopreservation damage. Theriogenology 2011, 76, 436–447. [Google Scholar] [CrossRef]
- Villemure, M.; Lazure, C.; Manjunath, P. Isolation and characterization of gelatin-binding proteins from goat seminal plasma. Reprod. Biol. Endocrinol. 2003, 1, 39. [Google Scholar] [CrossRef] [PubMed]
- Ménard, M.; Nauc, V.; Lazure, C.; Vaillancourt, D.; Manjunath, P. Novel purification method for mammalian seminal plasma phospholipid-binding proteins reveals the presence of a novel member of this family of proteins in stallion seminal fluid. Mol. Reprod. Dev. 2003, 66, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Plante, G.; Prud’homme, B.; Fan, J.; Lafleur, M.; Manjunath, P. Evolution and function of mammalian binder of sperm proteins. Cell Tissue Res. 2016, 363, 105–127. [Google Scholar] [CrossRef]
- Pini, T.; Farmer, K.; Druart, X.; Teixeira Gomes, A.P.; Tsikis, G.; Labas, V.; Leahy, T.; de Graaf, S.P. Binder of sperm proteins protect ram spermatozoa from freeze-thaw damage. Cryobiology 2018, 82, 78–87. [Google Scholar] [CrossRef]
- Manjunath, P.; Bergeron, A.; Lefebvre, J.; Fan, J. Seminal plasma proteins: Functions and interaction with protective agents during semen preservation. Soc. Reprod. Fertil. Suppl. 2007, 65, 217–228. [Google Scholar]
- Mendoza, N.; Casao, A.; Pérez-Pé, R.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. New insights into the mechanisms of ram sperm protection by seminal plasma proteins. Biol. Reprod. 2013, 88, 1–15. [Google Scholar] [CrossRef]
- Fraser, L.; Gorszczaruk, M.; Lecewicz, M.; Strzezek, J. Age-related changes and seasonal variations in boar sperm metabolism during liquid storage at 5° and 16 °C. J. Anim. Feed Sci. 2003, 12, 803–811. [Google Scholar] [CrossRef]
- Fraser, L.; Lecewicz, M.; Strzezek, J. Fluorometric assessments of viability and mitochondrial status of boar spermatozoa following liquid storage. Pol. J. Vet. Sci. 2002, 5, 85–92. [Google Scholar] [PubMed]
- Amaral, A.; Lourenço, B.; Marques, M.; Ramalho-Santos, J. Mitochondria functionality and sperm quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef]
- Marti, E.; Mara, L.; Marti, J.I.; Muino-Blanco, T.; Cebrian-Perez, J.A. Seasonal variations in antioxidant enzyme activity in ram seminal plasma. Theriogenology 2007, 67, 1446–1454. [Google Scholar] [CrossRef] [PubMed]
- Sikka, S.C.; Hellstrom, W.J. Role of oxidative stress and antioxidants in Peyronie’s disease. Int. J. Impot. Res. 2002, 14, 353–360. [Google Scholar] [CrossRef]
- Brooks, D.E. Biochemistry of the male accessory glands. In Marshall’s Physiology of Reproduction, 4th ed.; Lamming, G.E., Ed.; Churchill Livingstone: Edinburgh, UK, 1990; pp. 569–690. [Google Scholar]
- Koziorowska, M.; Koziorowski, M.; Strzezek, J.; Fraser, L. Seasonal changes in antioxidant defence systems in seminal plasma and fluids of the boar reproductive tract. Reprod. Biol. 2011, 11, 37–47. [Google Scholar] [CrossRef]
- Mislei, B.; Bucci, D.; Malama, E.; Bollwein, H.; Mari, G. Seasonal changes in ROS concentrations and sperm quality in unfrozen and frozen-thawed stallion semen. Theriogenology 2020, 144, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Carvajal-Serna, M.; Torres-Ruda, F.; Cardozo, J.A.; Grajales-Lombana, H.; Cebrián-Pérez, J.Á.; Muiño-Blanco, T.; Pérez-Pé, R.; Casao, A. Changes in melatonin concentrations in seminal plasma are not correlated with testosterone or antioxidant enzyme activity when rams are located in areas with an equatorial photoperiod. Anim. Reprod. Sci. 2019, 200, 22–30. [Google Scholar] [CrossRef]
- Corteel, J.M. Collection, Processing and Artificial Insemination of Goat Semen; Gall, C., Ed.; Goat Production, Academic Press: London, UK, 1981; pp. 171–191. [Google Scholar]
- Pellicer-Rubio, M.T.; Combarnous, Y. Deterioration of goat spermatozoa in skimmed milk-based extenders as a result of oleic acid released by the bulbourethral lipase BUSgp60. J. Reprod. Fertil. 1998, 112, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Iritani, A.; Nishikawa, Y. Studies on the egg yolk coagulating factor in goat semen. III. Release of some acids accompanied by the coagulating phenomena. Jpn. J. Anim. Reprod. 1963, 8, 109–112. [Google Scholar] [CrossRef]
- Pellicer-Rubio, M.T.; Magallon, T.; Combarnous, Y. Deterioration of goat sperm viability in milj extenders is due to a bulbourethral 60-kilodalton glycoprotein with triglyceride lipase activity. Biol. Reprod. 1997, 57, 1023–1031. [Google Scholar] [CrossRef]
- Sawyer, D.E.; Brown, D.B. The use on an in vitro sperm activation assay to detect chemically induced damage of human sperm nuclei. Reprod. Toxicol. 1995, 9, 351–357. [Google Scholar] [CrossRef]
- Upreti, G.C.; Hall, E.L.; Koppens, D.; Oliver, J.E.; Vishwanath, R. Studies on the measurement of phospholipase A2 (PLA2) and PLA2 inhibitor activities in ram semen. Anim. Reprod. Sci. 1999, 56, 107–121. [Google Scholar] [CrossRef]
- Ritar, A.J.; Salamon, S. Effects of month of collection, method of processing, concentration of egg yolk and duration of frozen storage on viability of Angora goat spermatozoa. Small Rumin. Res. 1991, 4, 29–37. [Google Scholar] [CrossRef]
- Jimenez-Rabadan, P.; Ramon, M.; Garcia-Alvarez, O.; Maroto-Morales, A.; del Olmo, E.; Perez-Guzman, M.D.; Bisbal, A.; Fernandez-Santos, M.R.; Garde, J.J.; Soler, A.J. Effect of semen collection method (artificial vagina vs. electroejaculation), extender and centrifugation on post-thaw sperm quality of Blanca-Celtiberica buck ejaculates. Anim. Reprod. Sci. 2012, 132, 88–95. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Season | |||
|---|---|---|---|---|
| Spring | Summer | Autumn | Winter | |
| Total motility | 32.6 ± 7 a | 21.1 ± 5 b | 41.7 ± 4 a | 33.3 ± 8 a |
| Progressive motility | 30.7 ± 9 a,b | 21.4 ± 5 b | 38.6 ± 4 a | 32.3 ± 9 a,b |
| Rapid | 39.4 ± 9 a,b | 28.8 ± 6 b | 45.8 ± 6 a | 41.2 ± 9 a |
| VCL | 21.6 ± 9 b | 22.4 ± 6 a,b | 33.4 ± 7 a | 21.7 ± 5 b |
| VSL * | 3.6 ± 11 | 10.5 ± 6 | 12.5 ± 10 | 5.5 ± 9 |
| VAP | 13.5 ± 11 | 18.5 ± 7 | 27.5 ± 8 | 17.3 ± 7 |
| Viability | 33 ± 3 a | 19.2 ± 4 b | 28.4 ± 3 a | 27.9 ± 4 a |
| Normal morphology | 32.8 ± 9 | 28.8 ± 2 | 31.8 ± 3 | 37.4 ± 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Kadili, S.; Kirschvink, N.; Raes, M.; Bister, J.L.; Archa, B.; Douaik, A.; Chentouf, M. Influence of Season and Liquid Storage at 16 °C on Beni Arouss Bucks’ Semen Quality. Animals 2020, 10, 1986. https://doi.org/10.3390/ani10111986
El Kadili S, Kirschvink N, Raes M, Bister JL, Archa B, Douaik A, Chentouf M. Influence of Season and Liquid Storage at 16 °C on Beni Arouss Bucks’ Semen Quality. Animals. 2020; 10(11):1986. https://doi.org/10.3390/ani10111986
Chicago/Turabian StyleEl Kadili, Sara, Nathalie Kirschvink, Marianne Raes, Jean Loup Bister, Bouchaib Archa, Ahmed Douaik, and Mouad Chentouf. 2020. "Influence of Season and Liquid Storage at 16 °C on Beni Arouss Bucks’ Semen Quality" Animals 10, no. 11: 1986. https://doi.org/10.3390/ani10111986
APA StyleEl Kadili, S., Kirschvink, N., Raes, M., Bister, J. L., Archa, B., Douaik, A., & Chentouf, M. (2020). Influence of Season and Liquid Storage at 16 °C on Beni Arouss Bucks’ Semen Quality. Animals, 10(11), 1986. https://doi.org/10.3390/ani10111986