Effect of Tourist Activities on Fecal and Salivary Glucocorticoids and Immunoglobulin A in Female Captive Asian Elephants in Thailand

 ,
,  ,
,  ,
,  , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethical Consent
2.2. Study Animals
2.3. Sample Collection
2.4. Physical Scoring Factors
2.5. Fecal Extraction
2.5.1. Fecal Extraction for GC Analysis
2.5.2. Fecal Extraction for IgA Analysis
2.6. Enzymeimmunoassays
2.7. Statistical Analysis
3. Results
3.1. Tourist Camp Activities
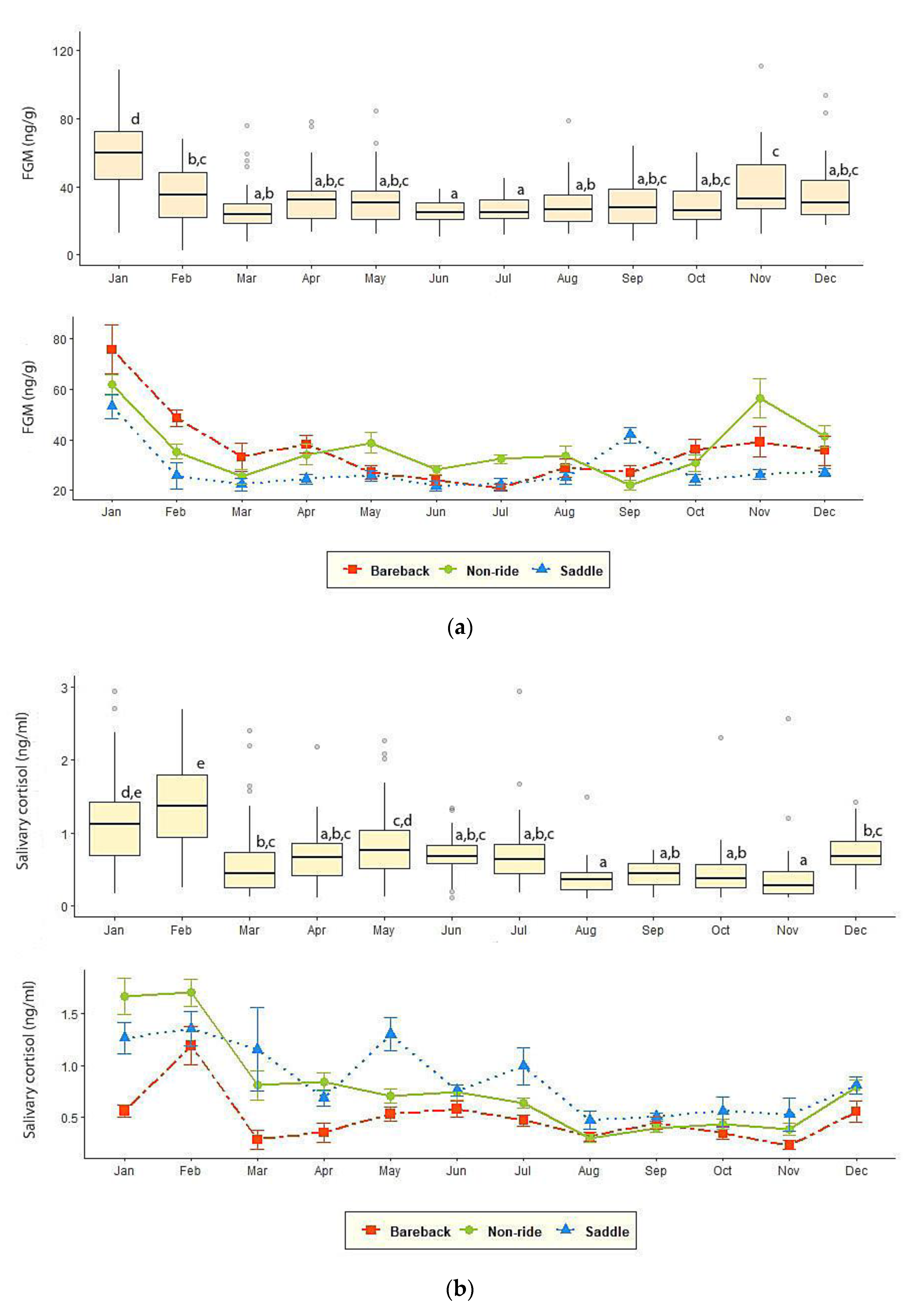
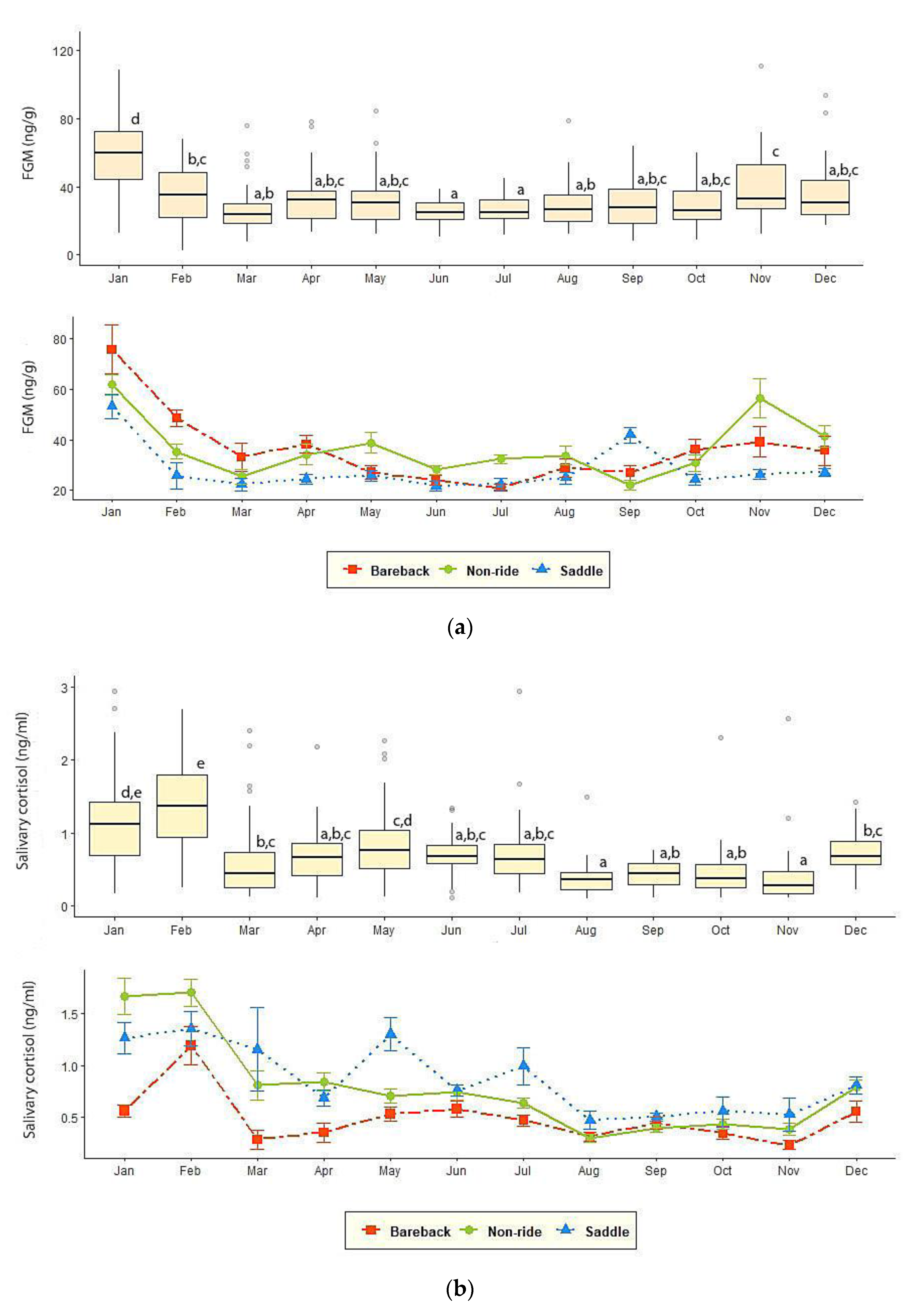
3.2. FGM and Salivary Cortisol Measures
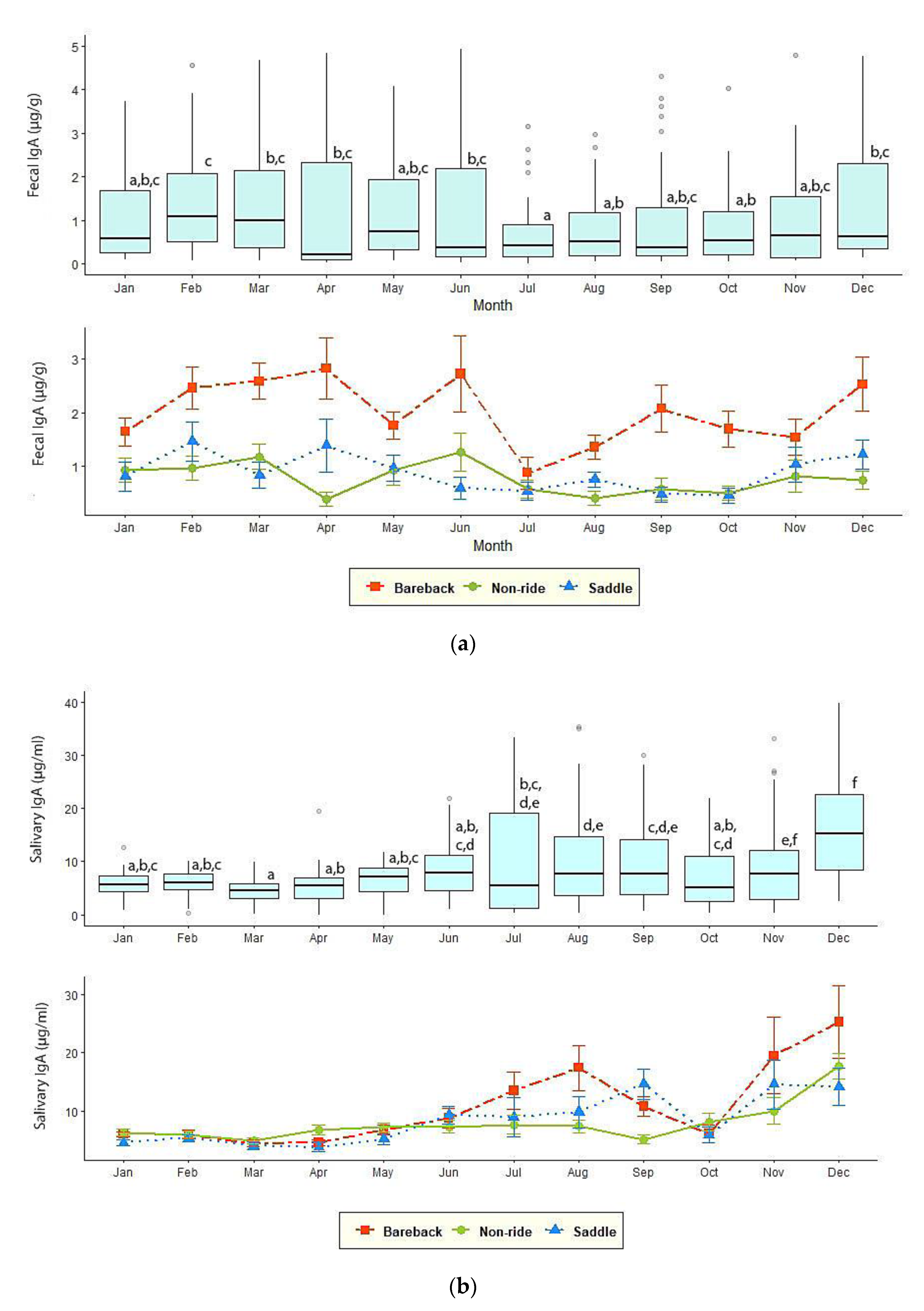
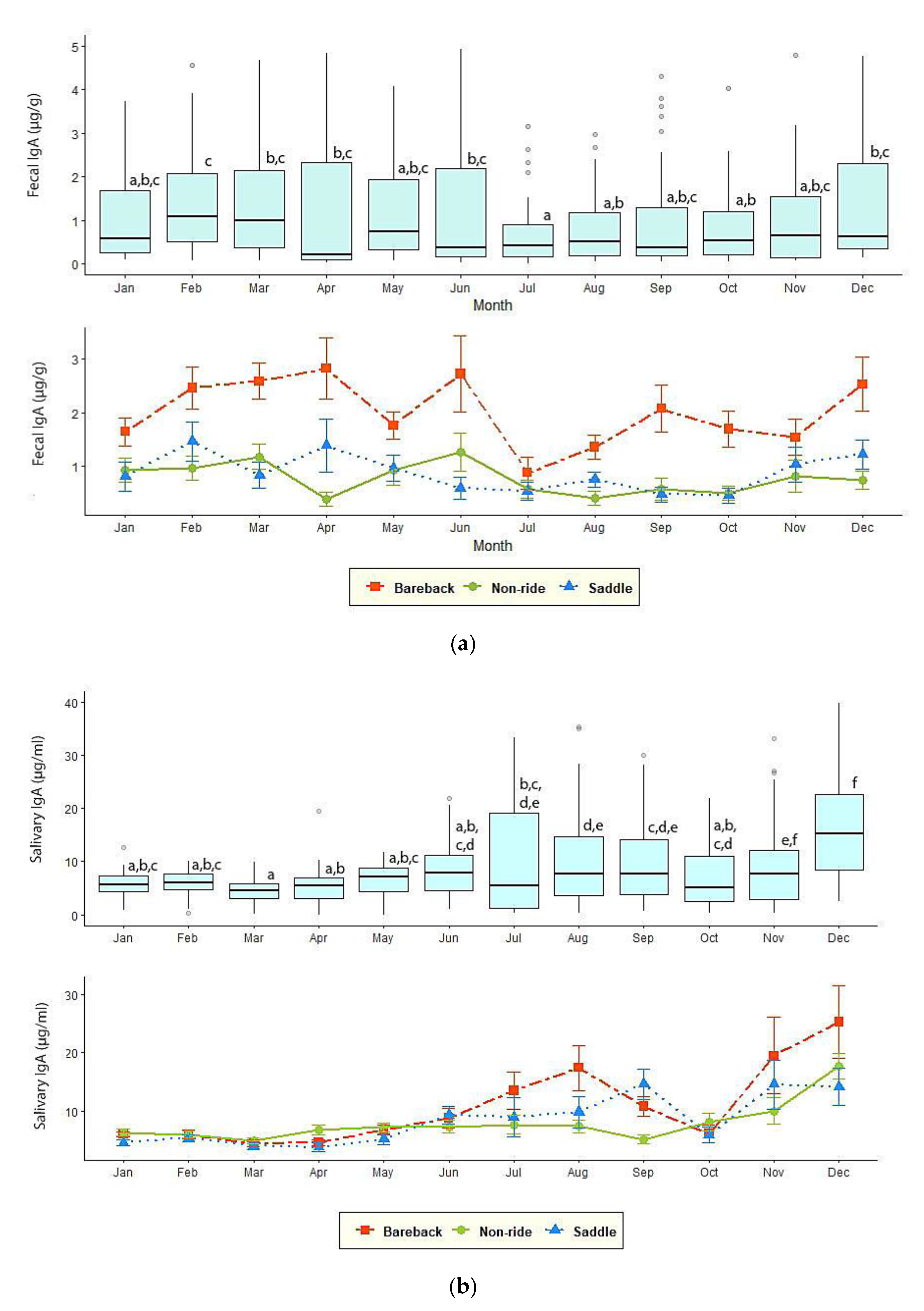
3.3. Fecal and Salivary IgA
3.4. Variable Relationships
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kontogeorgopoulos, N. The role of tourism in elephant welfare in northern Thailand. J. Tour. 2009, 10, 1–19. [Google Scholar]
- Bansiddhi, P.; Brown, J.L.; Thitaram, C. Welfare assessment and activities of captive elephants in Thailand. Animals 2020, 10, 919. [Google Scholar] [CrossRef] [PubMed]
- Bansiddhi, P.; Brown, J.L.; Khonmee, J.; Norkaew, T.; Nganvongpanit, K.; Punyapornwithaya, V.; Angkawanish, T.; Somgird, C.; Thitaram, C. Management factors affecting adrenal glucocorticoid activity of tourist camp elephants in Thailand and implications for elephant welfare. PLoS ONE 2019, 14, e0221537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norkaew, T.; Brown, J.L.; Thitaram, C.; Bansiddhi, P.; Somgird, C.; Punyapornwithaya, V.; Punturee, K.; Vongchan, P.; Somboon, N.; Khonmee, J. Associations among tourist camp management, high and low tourist seasons, and welfare factors in female Asian elephants in Thailand. PLoS ONE 2019, 14, e0218579. [Google Scholar] [CrossRef]
- Godfrey, A.; Kongmuang, C. Distribution, demography and basic husbandry of the Asian elephant in the tourism industry in Northern Thailand. Gajah 2009, 30, 13–18. [Google Scholar]
- Bansiddhi, P.; Brown, J.L.; Thitaram, C.; Punyapornwithaya, V.; Somgird, C.; Edwards, K.L.; Nganvongpanit, K. Changing trends in elephant camp management in northern Thailand and implications for welfare. PeerJ 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Norkaew, T.; Brown, J.L.; Bansiddhi, P.; Somgird, C.; Thitaram, C.; Punyapornwithaya, V.; Punturee, K.; Vongchan, P.; Somboon, N.; Khonmee, J. Body condition and adrenal glucocorticoid activity affects metabolic marker and lipid profiles in captive female elephants in Thailand. PLoS ONE 2018, 13, e0204965. [Google Scholar] [CrossRef] [Green Version]
- Cohen, E.H. Explorations in Thai Tourism: Collected Case Studies; Emerald Group Publishing: Warrington, UK, 2008; pp. 160–165. ISBN 978-0-08-046736-8. [Google Scholar]
- Kumar, V.; Pradheeps, M.; Kokkiligadda, A.; Niyogi, R.; Umapathy, G. Non-invasive assessment of physiological stress in captive Asian elephants. Animals 2019, 9, 553. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Bansiddhi, P.; Khonmee, J.; Thitaram, C. Commonalities in management and husbandry factors important for health and welfare of captive elephants in North America and Thailand. Animals 2020, 10, 737. [Google Scholar] [CrossRef] [Green Version]
- Mellor, D. Operational details of the five domains model and its key applications to the assessment and management of animal welfare. Animals 2017, 7, 60. [Google Scholar] [CrossRef] [Green Version]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W. Assessment of welfare in zoo animals: Towards optimum quality of life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menargues, A.; Limiñana, R.; Urios, V.; Mauri, M. Circadian rhythm of salivary cortisol in Asian elephants (Elephas maximus): A factor to consider during welfare assessment. J. Appl. Anim. Welf. Sci. 2012, 15, 383–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Carlstead, K.; Bray, J.D.; Dickey, D.; Farin, C.; Ange-van Heugten, K. Individual and environmental risk factors associated with fecal glucocorticoid metabolite concentrations in zoo-housed Asian and African elephants. PLoS ONE 2019, 14, e0217326. [Google Scholar] [CrossRef] [Green Version]
- Magda, S.; Spohn, O.; Angkawanish, T.; Smith, D.A.; Pearl, D.L. Risk factors for saddle-related skin lesions on elephants used in the tourism industry in Thailand. BMC Vet. Res. 2015, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plangsangmas, T.; Brown, J.L.; Thitaram, C.; Silva-Fletcher, A.; Edwards, K.L.; Punyapornwithaya, V.; Towiboon, P.; Somgird, C. Circadian rhythm of salivary immunoglobulin A and associations with cortisol as a stress biomarker in captive Asian elephants (Elephas maximus). Animals 2020, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Romero, L.M. Seasonal changes in plasma glucocorticoid concentrations in free-living vertebrates. Gen. Comp. Endocrinol. 2002, 128, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Marcilla, A.M.; Urios, V.; Limiñana, R. Seasonal rhythms of salivary cortisol secretion in captive Asian elephants (Elephas maximus). Gen. Comp. Endocrinol. 2012, 176, 259–264. [Google Scholar] [CrossRef]
- Fanson, K.V.; Keeley, T.; Fanson, B.G. Cyclic changes in cortisol across the estrous cycle in parous and nulliparous Asian elephants. Endocr. Connect. 2014, 3, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Somerville, M.; Riddle, H.S.; Keele, M.; Duer, C.K.; Freeman, E.W. Comparative endocrinology of testicular, adrenal and thyroid function in captive Asian and African elephant bulls. Gen. Comp. Endocrinol. 2007, 151, 153–162. [Google Scholar] [CrossRef]
- Huang, S.; Li, L.; Wu, J.; Li, C.; Bai, J.; Sun, Y.; Wang, G. Seasonal variations in immunoreactive cortisol and fecal immunoglobulin levels in Sichuan golden monkey (Rhinopithecus roxellana). Turk. J. Zool. 2014, 38, 642–650. [Google Scholar] [CrossRef]
- Yin, Y.J.; Nie, C.Y.; Liu, W.S.; Zou, Q.; Zhai, J.C.; Han, H.S.; Li, H.P. Non-invasive determination of the immune physiological state of reindeer (Rangifer tarandus) in the Greater Khingan Mountains, China. Genet. Mol. Res. 2015, 14, 6664–6673. [Google Scholar] [CrossRef] [PubMed]
- Lantz, E.L.; Lonsdorf, E.V.; Heintz, M.R.; Murray, C.M.; Lipende, I.; Travis, D.A.; Santymire, R.M. Non-invasive quantification of immunoglobulin A in chimpanzees (Pan troglodytes schweinfurthii) at Gombe National Park, Tanzania: Immunoglobulin A in Chimpanzees at Gombe. Am. J. Primatol. 2018, 80, e22558. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.L.; Bansiddhi, P.; Paris, S.; Galloway, M.; Brown, J.L. The development of an immunoassay to measure immunoglobulin A in Asian elephant feces, saliva, urine and serum as a potential biomarker of well-being. Conserv. Physiol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Staley, M.; Conners, M.G.; Hall, K.; Miller, L.J. Linking stress and immunity: Immunoglobulin A as a non-invasive physiological biomarker in animal welfare studies. Horm. Behav. 2018, 102, 55–68. [Google Scholar] [CrossRef]
- Decaro, N.; Pratelli, A.; Tinelli, A.; Martella, V.; Camero, M.; Buonavoglia, D.; Tempesta, M.; Caroli, A.M.; Buonavoglia, C. Fecal immunoglobulin A antibodies in dogs infected or vaccinated with canine coronavirus. CVI 2004, 11, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Tsujita, S.; Morimoto, K. Secretory IgA in saliva can be a useful stress marker. Environ. Health Prev. Med. 1999, 4, 1–8. [Google Scholar] [CrossRef]
- Campos-Rodríguez, R.P.; Godínez-Victoria, M.P.; Abarca-Rojano, E.P.; Pacheco-Yepez, J.P.; Reyna-Garfias, H.M.; Barbosa-Cabrera, R.E.P.; Drago-Serrano, M.E. Stress modulates intestinal secretory immunoglobulin A. Front. Integr. Neurosci. 2013, 7. [Google Scholar] [CrossRef] [Green Version]
- He, C.-S.; Tsai, M.-L.; Ko, M.-H.; Chang, C.-K.; Fang, S.-H. Relationships among salivary immunoglobulin A, lactoferrin and cortisol in basketball players during a basketball season. Eur. J. Appl. Physiol. 2010, 110, 989–995. [Google Scholar] [CrossRef]
- Royo, F.; Bjork, N.; Carlsson, H.; Mayo, S.; Hau, J. Impact of chronic catheterization and automated blood sampling (Accusampler) on serum corticosterone and fecal immunoreactive corticosterone metabolites and immunoglobulin A in male rats. J. Endocrinol. 2004, 180, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Pihl, L.; Hau, J. Faecal corticosterone and immunoglobulin A in young adult rats. Lab. Anim. 2003, 37, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Allwin, B.; Jayathangaraj, M.G. Kalaignan Faecal cortisol a non invasive biomarker for stress assessment in wild animals, confounding factors, estimation, quantification, and Interpretation. Int. J. Adv. Res. Biol.Sci. 2015, 2, 239–251. [Google Scholar]
- Morfeld, K.A.; Meehan, C.L.; Hogan, J.N.; Brown, J.L. Assessment of body condition in African (Loxodonta africana) and Asian (Elephas maximus) elephants in North American zoos and management practices associated with high body condition scores. PLoS ONE 2016, 11, e0155146. [Google Scholar] [CrossRef] [PubMed]
- Schein, M.; Rogers, P.N.; Leppäniemi, A.; Rosin, D. Schein’s Common Sense Prevention and Management of Surgical Complications: For Surgeons, Residents, Lawyers, and Even Those Who Never Have Any Complications; Tfm Publishing Limited: Harley Shrewsbury, UK, 2013; ISBN 978-1-903378-98-4. [Google Scholar]
- Almeida, A.; Loy, A.; Hofmann, H. Qqplotr: Quantile-Quantile Plot Extensions for “ggplot2”. 2020. Available online: https://cran.r-project.org/package=qqplotr (accessed on 11 June 2020).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models. 2020. Available online: https://cran.r-project.org/package=nlme (accessed on 11 June 2020).
- Højsgaard, S.; Halekoh, U.; Yan, J.; Ekstrøm, C. Geepack: Generalized Estimating Equation Package. 2019. Available online: https://cran.r-project.org/package=geepack (accessed on 20 September 2020).
- Lenth, R. Lsmeans: Least-Squares Means. 2018. Available online: https//cran.r-project.org/package=lsmeans (accessed on 11 June 2020).
- Bakdash, J.Z.; Marusich, L.R. Repeated measures correlation. Front. Psychol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, B.J.; Meehan, C.L.; Heinsius, J.L.; Mench, J.A. Why pace? The influence of social, housing, management, life history, and demographic characteristics on locomotor stereotypy in zoo elephants. Appl. Anim. Behav. Sci. 2017, 194, 104–111. [Google Scholar] [CrossRef]
- Meehan, C.L.; Mench, J.A.; Carlstead, K.; Hogan, J.N. Determining connections between the daily lives of zoo elephants and their welfare: An epidemiological approach. PLoS ONE 2016, 11, e0158124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romain, S.; Angkawanish, T.; Bampenpol, P.; Pongsopawijit, P.; Sombatphuthorn, P.; Nomsiri, R.; Silva-Fletcher, A. Diet composition, food intake, apparent digestibility, and body condition score of the captive Asian elephant (Elephas maximus): A pilot study in two collections in Thailand. J. Zoo. Wildlife Med. 2014, 45, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ange-van Heugten, K.D.; van Heugten, E.; Timmer, S.; Bosch, G.; Elias, A.; Whisnant, S.; Swarts, H.J.M.; Ferket, P.; Verstegen, M.W.A. Fecal and Salivary Cortisol Concentrations in Woolly (Lagothrix ssp.) and Spider Monkeys (Ateles spp.). Int. J. Zool. 2009, 2009, 1–9. [Google Scholar] [CrossRef]
- Otovic, P. Limits to using HPA axis activity as an indication of animal welfare. ALTEX 2015, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, C.E.; Thierfelder, T.; Svennersten-Sjaunja, K.; Berg, C.; Orihuela, A.; Lidfors, L. Time lag between peak concentrations of plasma and salivary cortisol following a stressful procedure in dairy cattle. Acta. Vet. Scand. 2014, 56, 61. [Google Scholar] [CrossRef] [Green Version]
- Wasser, S.K.; Hunt, K.E.; Brown, J.L.; Cooper, K.; Crockett, C.M.; Bechert, U.; Millspaugh, J.J.; Larson, S.; Monfort, S.L. A generalized fecal glucocorticoid assay for use in a diverse array of nondomestic mammalian and avian species. Gen. Comp. Endocrinol. 2000, 120, 260–275. [Google Scholar] [CrossRef]
- Grandin, T.; Shivley, C. How farm animals react and perceive stressful situations such as handling, restraint, and transport. Animals 2015, 5, 1233–1251. [Google Scholar] [CrossRef]
- Pajor, F.; Kovács, A.; Tőzsér, J.; Póti, P. The influence of temperament on cortisol concentration and metabolic profile in Tsigai lambs. Arch. Anim. Breed. 2013, 56, 573–580. [Google Scholar] [CrossRef]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the stress response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Burdick, N.C.; Carroll, J.A.; Hulbert, L.E.; Dailey, J.W.; Willard, S.T.; Vann, R.C.; Welsh, T.H.; Randel, R.D. Relationships between temperament and transportation with rectal temperature and serum concentrations of cortisol and epinephrine in bulls. Livest. Sci. 2010, 129, 166–172. [Google Scholar] [CrossRef]
- Grandin, T. Assessment of stress during handling and transport. J. Anim. Sci. 1997, 75, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Hall, S.J.G.; Broom, D.M.; Kiddy, G.N.S. Effect of transportation on plasma cortisol and packed cell volume in different genotypes of sheep. Small Rumin. Res. 1998, 29, 233–237. [Google Scholar] [CrossRef]
- Zavy, M.; Juniewicz, P.; Phillips, W.; VonTungeln, D. Effect of initial restraint, weaning, and transport stress on baseline and ACTH-stimulated cortisol responses in beef calves of different genotypes. Am. J. Vet. Res. 1992, 53, 551–557. [Google Scholar] [PubMed]
- Scott, K.; Heistermann, M.; Cant, M.A.; Vitikainen, E.I.K. Group size and visitor numbers predict faecal glucocorticoid concentrations in zoo meerkats. R. Soc. Open Sci. 2017, 4, 161017. [Google Scholar] [CrossRef] [Green Version]
- Rajagopal, T.; Archunan, G.; Sekar, M. Impact of Zoo Visitors on the Fecal Cortisol Levels and Behavior of an Endangered Species: Indian Blackbuck (Antelope cervicapra L.). J. Appl. Anim. Welf. Sci. 2011, 14, 18–32. [Google Scholar] [CrossRef]
- Webster, K.; Narayan, E.; de Vos, N. Fecal glucocorticoid metabolite response of captive koalas (Phascolarctos cinereus) to visitor encounters. Gen. Comp. Endocrinol. 2017, 244, 157–163. [Google Scholar] [CrossRef]
- Muehlenbein, M.P.; Ancrenaz, M.; Sakong, R.; Ambu, L.; Prall, S.; Fuller, G.; Raghanti, M.A. Ape conservation physiology: Fecal glucocorticoid responses in wild Pongo pygmaeus morio following human visitation. PLoS ONE 2012, 7, e33357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherwen, S.L.; Hemsworth, P.H.; Butler, K.L.; Fanson, K.V.; Magrath, M.J.L. Impacts of visitor number on kangaroos housed in free-range exhibits: Kangaroo welfare in free-range exhibits. Zoo Biol. 2015, 34, 287–295. [Google Scholar] [CrossRef]
- Fanson, K.V.; Lynch, M.; Vogelnest, L.; Miller, G.; Keeley, T. Response to long-distance relocation in Asian elephants (Elephas maximus): Monitoring adrenocortical activity via serum, urine, and feces. Eur. J. Wildl. Res. 2013, 59, 655–664. [Google Scholar] [CrossRef]
- Cook, N.J. Review: Minimally invasive sampling media and the measurement of corticosteroids as biomarkers of stress in animals. Can. J. Anim. Sci. 2012, 92, 227–259. [Google Scholar] [CrossRef]
- Paramastri, Y.; Royo, F.; Eberova, J.; Carlsson, H.-E.; Sajuthi, D.; Fernstrom, A.-L.; Pamungkas, J.; Hau, J. Urinary and fecal immunoglobulin A, cortisol and 11-17 dioxoandrostanes, and serum cortisol in metabolic cage housed female cynomolgus monkeys (Macaca fascicularis). J. Med. Primatol. 2007, 36, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Leicht, C.A.; Goosey-Tolfrey, V.L.; Bishop, N.C. Exercise intensity and its impact on relationships between salivary immunoglobulin A, saliva flow rate and plasma cortisol concentration. Eur. J. Appl. Physiol. 2018, 118, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- Muneta, Y.; Yoshikawa, T.; Minagawa, Y.; Shibahara, T.; Maeda, R.; Omata, Y. Salivary IgA as a useful non-invasive marker for restraint stress in pigs. J. Vet. Med. Sci. 2010, 72, 1295–1300. [Google Scholar] [CrossRef] [Green Version]
- Kawano, Y.; Demura, S.; Tanaka, Y.; Matsuura, Y. Physiological stress response based on salivary α-amylase activity and secretory immunoglobulin A levels in female care workers and general workers. Nur. Primary Care 2019, 3. [Google Scholar] [CrossRef]
- Birkett, M.; Johnson, L.; Gelety, C. Investigation of sex differences in sIgA response to the trier social stress test. Stress Health 2017, 33, 158–163. [Google Scholar] [CrossRef]
- Engels, H.-J.; Kendall, B.J.; Fahlman, M.M.; Gothe, N.P.; Bourbeau, K.C. Salivary immunoglobulin A in healthy adolescent females: Effects of maximal exercise, physical activity, body composition and diet. J. Sport Med. Phys. Fit. 2018, 58, 6. [Google Scholar]
- Usui, T.; Yoshikawa, T.; Orita, K.; Ueda, S.; Katsura, Y.; Fujimoto, S.; Yoshimura, M. Changes in salivary antimicrobial peptides, immunoglobulin A and cortisol after prolonged strenuous exercise. Eur. J. Appl. Physiol. 2011, 111, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Hanstock, H.G.; Walsh, N.P.; Edwards, J.P.; Fortes, M.B.; Cosby, S.L.; Nugent, A.; Curran, T.; Coyle, P.V.; Ward, M.D.; Yong, X.H.A. Tear fluid SIgA as a noninvasive biomarker of mucosal immunity and common cold risk. Med. Sci. Sports Exerc. 2016, 48, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Bansiddhi, P.; Nganvongpanit, K.; Brown, J.L.; Punyapornwithaya, V.; Pongsopawijit, P.; Thitaram, C. Management factors affecting physical health and welfare of tourist camp elephants in Thailand. PeerJ 2019, 7, e6756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.A.; Mudappa, D.; Raman, T.R.S. Asian elephant (Elephas Maximus) habitat use and ranging in fragmented rainforest and plantations in the Anamalai Hills, India. Trop. Conserv. Sci. 2010, 3, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Karaffová, V.; Bobíková, K.; Husáková, E.; Levkut, M.; Herich, R.; Revajová, V.; Levkutová, M.; Levkut, M. Interaction of TGF-β4 and IL-17 with IgA secretion in the intestine of chickens fed with E. faecium AL41 and challenged with S. Enteritidis. Res. Vet. Sci. 2015, 100, 75–79. [Google Scholar] [CrossRef]
- Rodríguez, O.L.; Hagel, I.; González, Y.; Roque, M.E.; Vásquez, N.; López, E.; Di Prisco, M.C. Secretory IgA antibody responses in Venezuelan children infected with Giardia duodenalis. J. Trop. Pediatr. 2004, 50, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Jarillo-Luna, R.A.; Rivera-Aguilar, V.; Pacheco-Yépez, J.; Godínez-Victoria, M.; Oros-Pantoja, R.; Miliar-García, A.; Campos-Rodríguez, R. Nasal IgA secretion in a murine model of acute stress. The possible role of catecholamines. J. Neuroimmunol. 2015, 278, 223–231. [Google Scholar] [CrossRef]
- Norkaew, T.; Brown, J.L.; Bansiddhi, P.; Somgird, C.; Thitaram, C.; Punyapornwithaya, V.; Punturee, K.; Vongchan, P.; Somboon, N.; Khonmee, J. Influence of season, tourist activities and camp management on body condition, testicular and adrenal steroids, lipid profiles, and metabolic status in captive Asian elephant bulls in Thailand. PLoS ONE 2019, 14, e0210537. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Paris, S.; Prado-Oviedo, N.A.; Meehan, C.L.; Hogan, J.N.; Morfeld, K.A.; Carlstead, K. Reproductive health assessment of female elephants in North American zoos and association of husbandry practices with reproductive dysfunction in african elephants (Loxodonta africana). PLoS ONE 2016, 11, e0145673. [Google Scholar] [CrossRef]
- Mumby, H.S.; Mar, K.U.; Thitaram, C.; Courtiol, A.; Towiboon, P.; Min-Oo, Z.; Htut-Aung, Y.; Brown, J.L.; Lummaa, V. Stress and body condition are associated with climate and demography in Asian elephants. Conserv. Physiol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Young, B.A. Temperature-induced changes in metabolism and body weight of cattle (Bos taurus). Can. J. Physiol. Pharmacol. 1975, 53, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Vaanholt, L.M.; Daan, S.; Schubert, K.A.; Visser, G.H. Metabolism and aging: Effects of cold exposure on metabolic rate, body composition, and longevity in mice. Physiol. Biochem. Zool. 2009, 82, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Corlatti, L.; Palme, R.; Frey-Roos, F.; Hackländer, K. Climatic cues and glucocorticoids in a free-ranging riparian population of red deer (Cervus elaphus). Folia Zool. 2011, 60, 176–180. [Google Scholar] [CrossRef]
- Huber, S.; Palme, R.; Arnold, W. Effects of season, sex, and sample collection on concentrations of fecal cortisol metabolites in red deer (Cervus elaphus). Gen. Comp. Endocrinol. 2003, 130, 48–54. [Google Scholar] [CrossRef]
- Garde, A.H.; Hansen, Å.M.; Skovgaard, L.T.; Christensen, J.M. Seasonal and biological variation of blood concentrations of total cholesterol, dehydroepiandrosterone sulfate, hemoglobin A1c, IgA, prolactin, and free testosterone in healthy women. Clin. Chem. 2000, 46, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber-Mzell, D.; Kotanko, P.; Hauer, A.C.; Goriup, U.; Haas, J.; Lanner, N.; Erwa, W.; Ahmaida, I.A.; Haitchi-Petnehazy, S.; Stenzel, M.; et al. Gender, age and seasonal effects on IgA deficiency: A study of 7293 Caucasians. Eur. J. Clin. Investig. 2004, 34, 224–228. [Google Scholar] [CrossRef]
- Agarwal, S.; Mayer, L. Diagnosis and treatment of gastrointestinal disorders in patients with primary immunodeficiency. Clin. Gastroenterol. Hepatol. 2013, 11, 1050–1063. [Google Scholar] [CrossRef] [Green Version]
- Chuaplaivech, P.; Pintawong, W.; Prawii, W. Retrospective studies of gastrointestinal tract disorder by National Institute of Elephant Research and Health Service from 1999–2008. JKV 2009, 19, 31–41. [Google Scholar]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. In Comprehensive Physiology; Terjung, R., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 603–621. ISBN 978-0-470-65071-4. [Google Scholar]
- Hansen, I.S.; Baeten, D.L.P.; den Dunnen, J. The inflammatory function of human IgA. Cell. Mol. Life Sci. 2019, 76, 1041–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Variables | Camp ID | |||||
|---|---|---|---|---|---|---|
| A | B | C | D | E | F | |
| Activity types | Saddle | Saddle | Bareback | Bareback | No-ride | No-ride |
| Total number of elephants (M/F) | 79 (34/45) | 52 (16/36) | 19 (1/18) | 5 (1/4) | 17 (4/13) | 47 (13/34) |
| Number of participating elephants | 9 | 5 | 9 | 2 | 6 | 13 |
| Numbers of tourists/d (range) | 200–300 | 100–200 | 20–40 | 7–15 | 30–50 | 50–80 |
| Tourist-to-elephant ratio (range) | 2.5–3.8 | 1.9–3.9 | 1.1–2.1 | 1.4–3 | 1.8–2.9 | 1.4–2.2 |
| Daily work hours | 5–6 | 5–6 | 4.5–6 | 4.5–5 | 4–6 | 4–6 |
| Chain hours total (day and night) | 17 | 17 | 16–18 | 17–18 | 17–18 | 13–18 |
| Chain hours (day only) | 3 | 3 | 3 | 3–4 | 3–4 | 0–2 |
| Daytime chain length (m) | 1.5–2 | 1.5 | 1.5 | 1.5–2 | 1.5 | 1.5 |
| Nighttime chain length (m) | 2–3 | 1.5 | 1.5 | 3–4 | 1.5 | 1.5 |
| Socialization time (h) 1 | 0 | 0 | 1 | 1 | 1 | 2–3 |
| Supplemental feeding by tourists | No | No | Yes | Yes | Yes | Yes |
| FGM (ng/g) | 25.8 ± 1.27 c | 33.0 ± 1.78 a,b,c | 36.30 ± 2.10 a,b | 35.65 ± 2.68 a,b,c | 31.11 ± 1.72 b,c | 39.25 ± 1.72 a |
| Salivary cortisol (ng/mL) | 0.89 ± 0.07 a | 0.82 ± 0.06 a,b | 0.50 ± 0.03 b,c | 0.44 ± 0.07 c | 0.71 ± 0.05 a,b,c | 0.83 ± 0.05 a |
| Fecal IgA (µg/g) | 0.68 ± 0.09 b | 1.26 ± 0.14 b | 2.30 ± 0.13 a | 0.69 ± 0.14 b | 0.67 ± 0.09 b | 0.82 ± 0.08 b |
| Salivary IgA (µg/mL) | 5.65 ± 0.52 b | 13.49 ± 1.52 a | 12.20 ± 1.19 a | 3.36 ± 0.41 b | 7.57 ± 0.73 a,b | 8.00 ± 0.51 a,b |
| Fecal GC/IgA 2 | 52.34 ± 12.45 a,b | 33.69 ± 4.05 b | 17.09 ± 2.67 b | 101.26 ± 32.06 a | 61.94 ± 10.65 a,b | 55.01 ± 7.99 a,b |
| Salivary GC/IgA 2 | 0.26 ± 0.07 a | 0.10 ± 0.03 a,b | 0.06 ± 0.01 b | 0.17 ± 0.05 a,b | 0.11 ± 0.02 a,b | 0.12 ± 0.03 a,b |
| Body condition score (1–5) | 3.84 ± 0.04 a,b | 3.73 ± 0.04 b,c | 4.01 ± 0.04 a | 3.31 ± 0.08 c | 4.09 ± 0.05 a | 4.10 ± 0.04 a |
| Foot score (0–3) | 0.89 ± 0.05 a,b | 0.98 ± 0.04 a | 0.53 ± 0.05 b | 0.60 ± 0.10 b | 0.85 ± 0.07 a,b | 0.90 ± 0.03 a |
| Wound score (0–2) | 0.25 ± 0.04 b,c | 0.52 ± 0.07 a | 0.07 ± 0.03 b,c | 0.31 ± 0.10 a,b | 0.06 ± 0.03 b,c | 0.03 ± 0.01 c |
| Variables | FGM | Salivary Cortisol | Fecal IgA | Salivary IgA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | SEM | p-Value | Estimate | SEM | p-Value | Estimate | SEM | p-Value | Estimate | SEM | p-Value | ||
| Age | (DF = 1523; F = 0.15; AIC = 4570.72) | (DF = 1519, F = 0.90; AIC = 957.51) | (DF = 1523; F = 0.24; AIC = 1577.13) | (DF = 1515; F = 4.11; AIC = 3693.48) | |||||||||
| −0.029 | 0.07 | 0.692 | 0.002 | 0.002 | 0.342 | 0.004 | 0.009 | 0.623 | 0.093 | 0.046 | 0.043 | ||
| Work types | (DF = 2522; F = 9.72; AIC = 4546.44) | (DF = 2518; F = 19.60; AIC = 950.51) | (DF = 2522; F = 15.84; AIC = 1549.68) | (DF = 2514; F = 1.59; AIC = 3686.76) | |||||||||
| None | Reference | ||||||||||||
| Bareback | −0.498 | 2.144 | 0.816 | −0.299 | 0.059 | <0.01 | 1.243 | 0.232 | <0.01 | 2.792 | 1.597 | 0.081 | |
| Saddle | −8.267 | 1.995 | <0.01 | 0.073 | 0.054 | 0.177 | 0.111 | 0.215 | 0.604 | 0.521 | 1.485 | 0.725 | |
| Camp size | (DF = 2522; F = 0.09; AIC = 4560.92) | (DF = 2518; F = 15.93; AIC = 932.01) | (DF = 2522; F = 6.53; AIC = 1560.53) | (DF = 2514; F = 3.21; AIC = 3682.57) | |||||||||
| Large | Reference | ||||||||||||
| Medium | 0.560 | 2.207 | 0.799 | −0.260 | 0.052 | <0.01 | 0.803 | 0.228 | <0.01 | 2.203 | 1.312 | 0.093 | |
| Small | 2.008 | 5.014 | 0.689 | −0.409 | 0.118 | <0.01 | −0.158 | 0.522 | 0.761 | −4.831 | 2.992 | 0.107 | |
| Tourist-to-elephant ratio | (DF = 2522; F = 21.28; AIC = 4533.05) | (DF = 2518; F = 3.13; AIC = 950.88) | (DF = 2522; F = 4.01; AIC = 1565.70) | (DF = 2514; F = 3.36; AIC = 3683.47) | |||||||||
| High | Reference | ||||||||||||
| Moderate | 6.745 | 2.093 | <0.01 | −0.176 | 0.087 | 0.043 | 0.228 | 0.323 | 0.481 | 3.506 | 1.761 | 0.047 | |
| Low | 12.254 | 1.914 | <0.01 | −0.194 | 0.079 | 0.015 | 0.752 | 0.295 | 0.011 | 4.093 | 1.601 | 0.011 | |
| Daily work hours | (DF = 1523; F = 13.36; AIC = 4551.26) | (DF = 1519; F = 2.36; AIC = 948.01) | (DF = 1523; F = 0.69; AIC = 1568.63) | (DF = 1515; F = 1.51; AIC = 3687.83) | |||||||||
| −13.804 | 3.775 | <0.01 | 0.205 | 0.133 | 0.124 | 0.416 | 0.498 | 0.403 | 3.331 | 2.706 | 0.218 | ||
| Total chain hours | (DF = 1523; F = 15.59; AIC = 4550.82) | (DF = 1519; F = 3.26; AIC = 948.48) | (DF = 1523; F = 2.74; AIC = 1568.75) | (DF = 1515; F = 0.52; AIC = 3690.92) | |||||||||
| <17 h | Reference | ||||||||||||
| ≥17 h | −7.556 | 1.913 | <0.01 | −0.123 | 0.07 | 0.071 | 0.427 | 0.258 | 0.098 | 1.031 | 1.418 | 0.467 | |
| Socialization time | (DF = 2522; F = 14.30; AIC = 4540.70) | (DF = 2518; F = 14.97; AIC = 934.44) | (DF = 2522; F = 4.59; AIC = 1564.92) | (DF = 2514; F = 0.55; AIC = 3688.80) | |||||||||
| 0-h | Reference | ||||||||||||
| 1-h | 5.967 | 1.907 | <0.01 | −0.296 | 0.059 | <0.01 | 0.658 | 0.266 | 0.013 | 1.190 | 1.559 | 0.445 | |
| 2-h | 10.834 | 2.032 | <0.01 | −0.037 | 0.063 | 0.553 | −0.066 | 0.283 | 0.814 | −0.381 | −0.229 | 0.818 | |
| Tourist feeding | (DF = 1523; F = 19.84; AIC = 4547.85) | (DF = 1519; F = 8.51; AIC = 943.93) | (DF = 1523; F = 1.81; AIC = 1568.86) | (DF = 1515; F = 0.13; AIC = 3690.54) | |||||||||
| No | Reference | ||||||||||||
| Yes | 8.083 | 1.814 | <0.01 | −0.184 | 0.063 | <0.01 | 0.343 | 0.255 | 0.178 | 0.505 | 1.397 | 0.717 | |
| Months | (DF = 11,513; F = 19.31; AIC = 4367.19) | (DF = 11,509; F = 21.56; AIC = 807.05) | (DF = 11,513; F = 3.82; AIC = 1568.63) | (DF = 11,505; F = 13.41; AIC = 3557.42) | |||||||||
| January | Reference | ||||||||||||
| February | −26.844 | 3.273 | <0.01 | 0.202 | 0.102 | 0.047 | 0.429 | 0.202 | 0.034 | 0.076 | 1.530 | 0.960 | |
| March | −35.979 | 3.273 | <0.01 | −0.473 | 0.102 | <0.01 | 0.348 | 0.202 | 0.086 | −1.149 | 1.530 | 0.453 | |
| April | −30.292 | 3.273 | <0.01 | −0.589 | 0.102 | <0.01 | 0.241 | 0.202 | 0.233 | −0.419 | 1.530 | 0.783 | |
| May | −30.608 | 3.273 | <0.01 | −0.406 | 0.102 | <0.01 | 0.080 | 0.203 | 0.693 | 0.713 | 1.539 | 0.643 | |
| June | −37.322 | 3.273 | <0.01 | −0.555 | 0.102 | <0.01 | 0.347 | 0.202 | 0.086 | 2.672 | 1.539 | 0.083 | |
| July | −35.958 | 3.293 | <0.01 | −0.547 | 0.103 | <0.01 | −0.431 | 0.202 | 0.033 | 3.796 | 1.549 | 0.014 | |
| August | −32.672 | 3.293 | <0.01 | −0.904 | 0.105 | <0.01 | −0.262 | 0.202 | 0.195 | 4.999 | 1.549 | <0.01 | |
| September | −32.813 | 3.273 | <0.01 | −0.818 | 0.102 | <0.01 | −0.157 | 0.202 | 0.438 | 3.980 | 1.539 | 0.010 | |
| October | −32.484 | 3.293 | <0.01 | −0.810 | 0.102 | <0.01 | −0.250 | 0.202 | 0.216 | 1.201 | 1.549 | 0.438 | |
| November | −19.914 | 3.273 | <0.01 | −0.868 | 0.102 | <0.01 | −0.001 | 0.202 | 0.995 | 8.131 | 1.530 | <0.01 | |
| December | −27.003 | 3.273 | <0.01 | −0.523 | 0.102 | <0.01 | 0.246 | 0.205 | 0.229 | 12.519 | 1.549 | <0.01 | |
| Environmental season | (DF = 2522; F = 47.39; AIC = 4477.03) | (DF = 2518; F = 32.12; AIC = 897.62) | (DF = 2522; F = 15.69; AIC = 1547.01) | (DF = 2514; F = 15.58; AIC = 3662.10) | |||||||||
| Rainy | Reference | ||||||||||||
| Summer | −0.081 | 1.792 | 0.963 | 0.262 | 0.059 | <0.01 | 0.531 | 0.101 | <0.01 | −3.060 | 0.854 | <0.01 | |
| Winter | 15.028 | 1.792 | <0.01 | 0.472 | 0.058 | <0.01 | 0.443 | 0.101 | <0.01 | 1.597 | 0.854 | 0.062 | |
| Tourist season | (DF = 1523; F = 94.97; AIC = 4478.04) | (DF = 1519; F = 42.73; AIC = 911.24) | (DF = 1523; F = 3.87; AIC = 1568.87) | (DF = 1515; F = 17.89; AIC = 3674.32) | |||||||||
| High | Reference | ||||||||||||
| Low | −15.069 | 1.546 | <0.01 | −0.338 | 0.051 | <0.01 | −0.178 | 0.091 | 0.049 | −3.151 | 0.744 | <0.01 | |
| Variables | Sample | Type | GC/IgA Ratio | p-Value |
|---|---|---|---|---|
| Work activity | Feces | Saddle | 37.0 ± 6.0 a,b | <0.01 |
| Bareback | 19.36 ± 2.79 b | |||
| No-ride | 52.68 ± 5.95 a | |||
| Saliva | Saddle | 0.14 ± 0.09 | 0.066 | |
| Bareback | 0.06 ± 0.05 | |||
| No-ride | 0.11 ± 0.08 | |||
| Camp size | Feces | Small | 101.23 ± 32.06 a | 0.014 |
| Medium | 21.95 ± 2.37 b | |||
| Large | 42.14 ± 4.64 a,b | |||
| Saliva | Small | 0.17 ± 0.05 | 0.136 | |
| Medium | 0.07 ± 0.01 | |||
| Large | 0.13 ± 0.03 | |||
| Tourist-to-elephant ratio | Feces | High | 52.15 ± 12.45 | 0.147 |
| Moderate | 39.86 ± 4.99 | |||
| Low | 29.13 ± 4.0 | |||
| Saliva | High | 0.26 ± 0.07 a | 0.015 | |
| Moderate | 0.10 ± 0.02 b | |||
| Low | 0.09 ± 0.02 b | |||
| Environmental season | Feces | Summer | 22.01 ± 2.34 | 0.086 |
| Rainy | 36.59 ± 1.84 | |||
| Winter | 37.29 ± 7.73 | |||
| Saliva | Summer | 0.13 ± 0.02 | 0.259 | |
| Rainy | 0.05 ± 0.01 | |||
| Winter | 0.13 ± 0.06 | |||
| Tourist season | Feces | High | 37.29 ± 7.73 | 0.269 |
| Low | 29.3 ± 3.08 | |||
| Saliva | High | 0.13 ± 0.06 | 0.370 | |
| Low | 0.09 ± 0.02 | |||
| Body condition score | Feces | 3 | 53.5 ± 12.66 a | 0.041 |
| 4 | 25.79 ± 3.19 b | |||
| 5 | 36.85 ± 11.51 a,b | |||
| Saliva | 3 | 0.27 ± 0.05 a | <0.01 | |
| 4 | 0.11 ± 0.03 b | |||
| 5 | 0.03 ± 0.01 b | |||
| Foot score | Feces | 0 | 21.92 ± 4.37 b | <0.01 |
| 1 | 28.25 ± 3.83 b | |||
| 2 | 66.08 ± 13.74 a | |||
| Saliva | 0 | 0.09 ± 0.02 | 0.931 | |
| 1 | 0.10 ± 0.02 | |||
| 2 | 0.1 ± 0.01 | |||
| Wound score | Feces | 0 | 23.45 ± 2.78 | 0.638 |
| 1 | 26.34 ± 5.57 | |||
| Saliva | 0 | 0.08 ± 0.02 | 0.838 | |
| 1 | 0.09 ± 0.03 |
| Parameters | FGM n = 525 | Salivary Cortisol n = 521 | Fecal IgA n = 525 | Salivary IgA n = 521 |
|---|---|---|---|---|
| FGM | 1.00 | |||
| Salivary cortisol | 0.14 * | 1.00 | ||
| Fecal IgA | 0.005 | 0.05 | 1.00 | |
| Salivary IgA | −0.03 | −0.08 | −0.04 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosaruk, W.; Brown, J.L.; Plangsangmas, T.; Towiboon, P.; Punyapornwithaya, V.; Silva-Fletcher, A.; Thitaram, C.; Khonmee, J.; Edwards, K.L.; Somgird, C. Effect of Tourist Activities on Fecal and Salivary Glucocorticoids and Immunoglobulin A in Female Captive Asian Elephants in Thailand. Animals 2020, 10, 1928. https://doi.org/10.3390/ani10101928
Kosaruk W, Brown JL, Plangsangmas T, Towiboon P, Punyapornwithaya V, Silva-Fletcher A, Thitaram C, Khonmee J, Edwards KL, Somgird C. Effect of Tourist Activities on Fecal and Salivary Glucocorticoids and Immunoglobulin A in Female Captive Asian Elephants in Thailand. Animals. 2020; 10(10):1928. https://doi.org/10.3390/ani10101928
Chicago/Turabian StyleKosaruk, Worapong, Janine L. Brown, Tithipong Plangsangmas, Patcharapa Towiboon, Veerasak Punyapornwithaya, Ayona Silva-Fletcher, Chatchote Thitaram, Jaruwan Khonmee, Katie L. Edwards, and Chaleamchat Somgird. 2020. "Effect of Tourist Activities on Fecal and Salivary Glucocorticoids and Immunoglobulin A in Female Captive Asian Elephants in Thailand" Animals 10, no. 10: 1928. https://doi.org/10.3390/ani10101928
APA StyleKosaruk, W., Brown, J. L., Plangsangmas, T., Towiboon, P., Punyapornwithaya, V., Silva-Fletcher, A., Thitaram, C., Khonmee, J., Edwards, K. L., & Somgird, C. (2020). Effect of Tourist Activities on Fecal and Salivary Glucocorticoids and Immunoglobulin A in Female Captive Asian Elephants in Thailand. Animals, 10(10), 1928. https://doi.org/10.3390/ani10101928