Polymorphism of the Melanocortin 1 Receptor (MC1R) Gene and its Role in Determining the Coat Colour of Central European Cattle Breeds

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area, Data Collection and Characteristics of Populations Selected for Research
2.2. Molecular Identification
2.3. Statistical Analysis
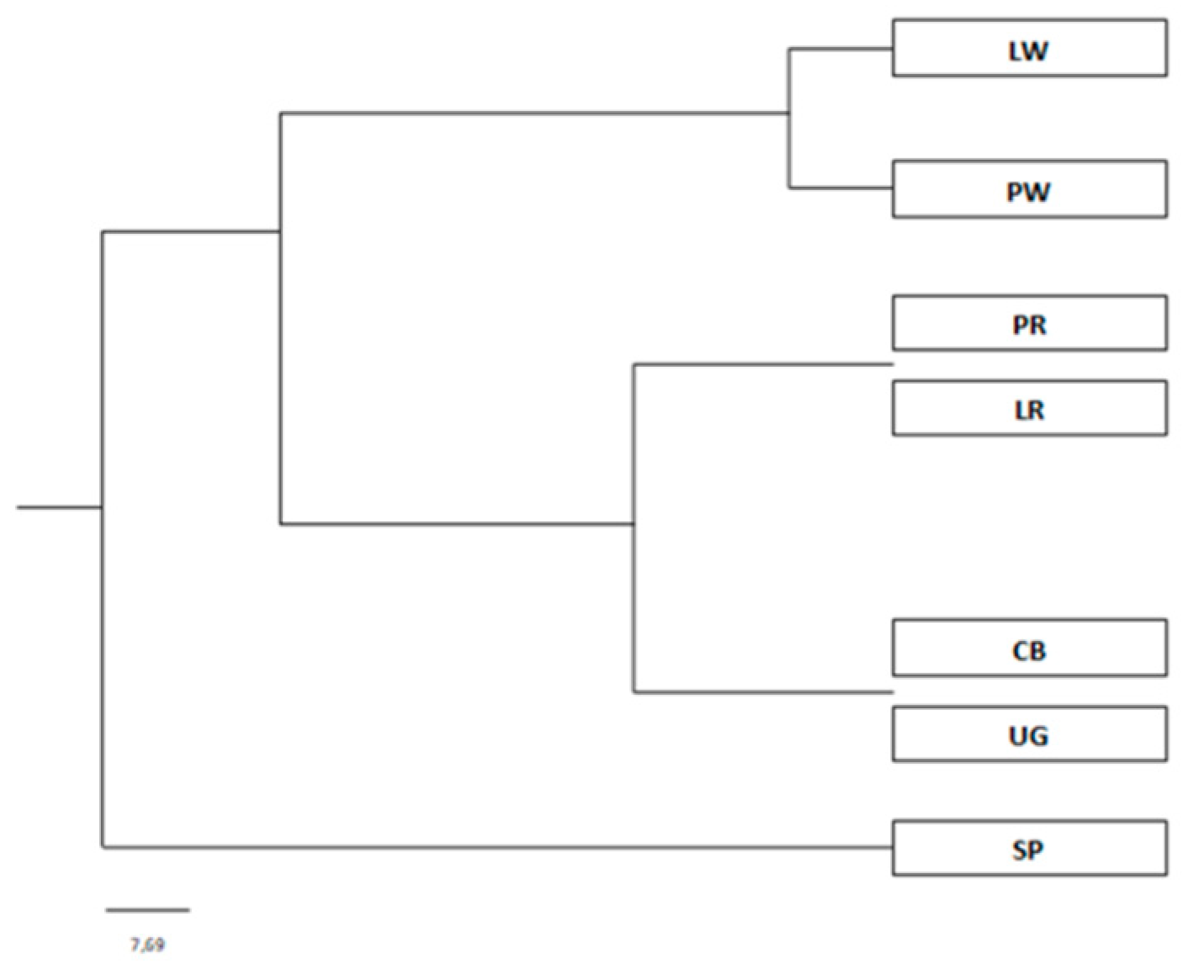
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, I.J. Colour-coded switches. Nature 1993, 362, 587–588. [Google Scholar] [CrossRef] [PubMed]
- Klungland, H.; Vage, D.I.; Gomez-Raya, L.; Adalsteinsson, S.; Lien, S. The role of melanocyte-stimulating hormone (MSH) receptor in bovine coat color determination. Mamm. Genome 1995, 6, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Searle, A.G. Comparative Genetics of Coat Colour in Mammals; Logos Press Ltd.: London, UK, 1968. [Google Scholar]
- Fontanesi, L.; Beretti, F.; Riggio, V.; Dall’Olio, S.; González, E.G.; Finocchiaro, R.; Davoli, R.; Russo, V.; Portolano, B. Missense and nonsense mutations in melanocortin 1 receptor (MC1R) gene of different goat breeds: Association with red and black coat colour phenotypes but with unexpected evidences. BMC Genet. 2009, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Chhajlani, V.; Wikberg, J.E.S. Molecular cloning and expression of the human melanocyte stimulating hormone receptor cDNA. FEBS Lett. 1992, 309, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Mountjoy, K.G.; Robbins, L.S.; Mortrud, M.T.; Cone, R.D. The cloning of a family of genes that encode the melanocortin receptors. Science 1992, 257, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkinson, W.O.; et al. Agouti protein is an antagonist of the melanocyte-stimulating-hormone receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes Dev. 1998, 12, 316–330. [Google Scholar] [CrossRef] [Green Version]
- Valverde, P.; Healy, E.; Jackson, I.; Rees, J.; Thody, A. Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat. Genet. 1995, 11, 328–330. [Google Scholar] [CrossRef]
- Kijas, J.M.H.; Wales, R.; Törnsten, A.; Chardon, P.; Moller, M.; Andersson, L. Melanocortin receptor 1 (MC1R) mutations and coat color in pigs. Genetics 1998, 150, 1177–1185. [Google Scholar]
- Eizirik, E.; Yuhki, N.; Johnson, W.E.; Menotti-Raymond, M.; Hannah, S.S.; O’Brien, S.J. Molecular genetics and evolution of melanism in the cat family. Curr. Biol. 2003, 13, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Robbins, L.; Nadeau, J.; Johnson, K.R.; Kelly, M.; Roselli-Rehfuss, L.; Baack, E. Pigmentation phenotypes of variant extension locus alleles result from point mutations that alter MSH receptor function. Cell 1993, 72, 827–834. [Google Scholar] [CrossRef]
- Våge, D.I.; Klungland, H.; Dongsi, L.; Cone, R.D. Molecular and pharmacological characterization of dominant black coat color in sheep. Mamm. Genome 1999, 10, 39–43. [Google Scholar] [CrossRef]
- Everts, R.E.; Rothuizen, J.; Van Oost, B.A. Identification of a premature stop codon in the melanocyte-stimulating hormone receptor gene (MC1R) in Labrador and Golden retrievers with yellow coat colour. Anim. Genet. 2000, 31, 194–199. [Google Scholar] [CrossRef]
- Våge, D.I.; Lu, D.; Klungland, H.; Lien, S.; Adalsteinsson, S.; Cone, R.D. A non-epistatic interaction of agouti and extension in the fox, Vulpes vulpes. Nat. Genet. 1997, 15, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Ritland, K.; Newton, C.; Marshall, H.D. Inheritance and population structure of the white-phased “Kermode” black bear. Curr. Biol. 2001, 11, 1468–1472. [Google Scholar] [CrossRef] [Green Version]
- Marklund, L.; Moller, M.; Sandberg, K.; Andersson, L. A missense mutation in the gene for melanocyte-stimulating hormone receptor (MC1R) is associated with the chestnut coat color in horses. Mamm. Genome 1996, 7, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Joerg, H.; Fries, H.; Meijerink, E.; Stranzinger, G. Red coat color in Holstein cattle is associated with a deletion in the MSHR gene. Mamm. Genome 1996, 7, 317–318. [Google Scholar] [CrossRef]
- Rouzaud, F.; Martin, J.; Gallet, P.F.; Delourme, D.; Goulemont-Leger, V.; Amigues, Y.; Ménissier, F.; Levéziel, H.; Julien, R.; Oulmouden, A. A first genotyping assay of French cattle breeds based on a new allele of the extension gene encoding the melanocortin-1 receptor (MC1R). Genet. Sel. Evol. 2000, 32, 511–520. [Google Scholar] [CrossRef]
- Adalsteinsson, S.; Bjarnadottir, S.; Vage, D.; Jonmundsson, J. Brown coat color in Icelandic cattle produced by the loci Extension and Agouti. J. Hered. 1994, 86, 395–398. [Google Scholar] [CrossRef]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Litwińczuk, Z.; Chabuz, W.; Stanek, P.; Sawicka, W. Genetic potential and reproductive performance of Whitebacks—Polish native breed of cows. Arch. Tierz. 2006, 49, 289–296. [Google Scholar]
- Litwińczuk, Z.; Stanek, P.; Chabuz, W.; Jankowski, P. Whitebacks—the native cattle of the Polesie region. Teka Comm. Prot. Form. Nat. Environ. 2004, 1, 130–138. [Google Scholar]
- Szarek, J.; Adamczyk, K.; Felenczak, A. Polish Red Cattle breeding: past and present. Anim. Genet. Resour. Inf. 2004, 35, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, P.D.; Lancioni, H.; Ceccobelli, S.; Colli, L.; Cardinali, I.; Karsli, T.; Capodiferro, M.R.; Sahin, E.; Ferretti, L.; Marsan, P.A.; et al. Mitochondrial DNA variants of Podolian cattle breeds testify for a dual maternal origin. PLoS ONE 2018, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ruban, S.Y.; Prijma, S.V. Animal genetic resources of Ukraine: Current status and perspectives. J. Vet. Med. Biotechnol. Biosaf. 2015, 1, 23–31. [Google Scholar]
- Alderson, L. The categorisation of types and breeds of cattle in Europe. Arch. Zootec. 1992, 41, 4. [Google Scholar]
- Russo, V.; Fontanesi, L.; Scotti, E.; Tazzoli, M.; Dall’Olio, S.; Davoli, R. Analysis of melanocortin 1 receptor (MC1R) gene polymorphisms in some cattle breeds: Their usefulness and application for breed traceability and authentication of Parmigiano Reggiano cheese. Ital. J. Anim. Sci. 2007, 6, 257–272. [Google Scholar] [CrossRef]
- Maudet, C.; Taberlet, P. Holstein’s milk detection in cheeses inferred from melanocortin receptor 1 (MC1R) gene polymorphism. J. Dairy Sci. 2002, 85, 707–715. [Google Scholar] [CrossRef]
- Crepaldi, P.; Marilli, M.; Gorni, C.; Meggiolaro, D.; Cicogna, M.; Renieri, C. Preliminary study on MC1R polymorphism in some cattle breeds raised in Italy. Ital. J. Anim. Sci. 2003, 2, 13–15. [Google Scholar] [CrossRef]
- Rolando, A.; Di Stasio, L. MC1R gene analysis applied to breed traceability of beef. Ital. J. Anim. Sci. 2006, 5, 87–91. [Google Scholar] [CrossRef]
- Lee, S.S.; Yang, Y.H.; Kang, S.Y.; Oh, W.Y.; Yang, B.S.; Ko, S.B.; Kim, K.I. Comparison of the genotypes and frequencies of MSH receptor (MC1R) gene in Korean Cattle, Cheju Native Black Cattle, Japanese Black and Japanese Brown Cattle. Korean J. Anim. Sci. 2000, 42, 253–260. [Google Scholar]
- Brenig, B.; Beck, J.; Floren, C.; Bornemann-Kolatzki, K.; Wiedemann, I.; Hennecke, S.; Swalve, H.; Schütz, E. Molecular genetics of coat colour variations in White Galloway and White Park cattle. Anim. Genet. 2013, 44, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Putra, D.E.; Paul, R.C.; Thu, L.N.A.; Okuda, Y.; Ibi, T.; Kunieda, T. Genetic characterization of Indonesian Pesisir cattle using mitochondrial DNA and Y-chromosomal haplotypes and loci associated with economical traits and coat color. J. Anim. Genet. 2018, 46, 17–23. [Google Scholar] [CrossRef]
- Niemi, M.; Sajantila, A.; Vilkki, J. Temporal variation in coat colour (genotypes) supports major changes in the Nordic cattle population after Iron Age. Anim. Genet. 2016, 47, 495–498. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak-Filipek, K.; Sawicka-Zugaj, W.; Litwińczuk, Z.; Chabuz, W.; Šveistienė, R.; Bulla, J. Assessment of the genetic structure of Central European cattle breeds based on functional gene polymorphism. Glob. Ecol. Conserv. 2019, 17. [Google Scholar] [CrossRef]
- Sawicka-Zugaj, W.; Chabuz, W.; Litwińczuk, Z.; Kasprzak-Filipek, K. Evaluation of reproductive performance and genetic variation in bulls of the Polish White-Backed breed. Reprod. Domest. Anim. 2018, 53, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Mei, C.; Wang, H.; Liao, Q.; Wang, L.; Cheng, G.; Wang, H.; Zhao, C.; Zhao, S.; Song, J.; Guang, X.; et al. Genetic architecture and selection of Chinese cattle revealed by whole genome resequencing. Mol. Biol. Evol. 2018, 35, 688–699. [Google Scholar] [CrossRef]
- Ludwig, A.; Pruvost, M.; Reissmann, M.; Benecke, N.; Brockmann, G.A.; Castaños, P.; Cieslak, M.; Lippold, S.; Llorente, L.; Malaspinas, A.S.; et al. Coat color variation at the beginning of Horse domestication. Science 2009, 324, 485. [Google Scholar] [CrossRef] [Green Version]
- Felius, M.; Beerling, M.L.; Buchanan, D.S.; Theunissen, B.; Koolmees, P.A.; Lenstra, J.A. On the history of cattle genetic resources. Diversity 2014, 6, 705–750. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Heterozygosity Observed (Ho) | Genotype in MC1R Locus | |||||
|---|---|---|---|---|---|---|---|
| ED/e | E+/e | ED/E+ | ED/ED | E+/E+ | e/e | ||
| LW | 1.00 | 84% | 16% | ||||
| PW | 0.98 | 28% | 6% | 64% | 2% | ||
| PR | 0.20 | 20% | 80% | ||||
| LR | 0.12 | 12% | 88% | ||||
| CB | 1.00 | 100% | |||||
| UG | 1.00 | 100% | |||||
| SP | 0.00 | 100% | |||||
| Alleles | Allele Frequency in Breeds | Overall Frequency | ||||||
|---|---|---|---|---|---|---|---|---|
| LW | PW | PR | LR | CB | UG | SP | ||
| ED | 0.50 | 0.46 | - | - | - | - | - | 0.17 |
| E+ | 0.08 | 0.37 | 0.90 | 0.94 | 0.50 | 0.50 | - | 0.46 |
| e | 0.42 | 0.17 | 0.10 | 0.06 | 0.50 | 0.50 | 1.00 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasprzak-Filipek, K.; Sawicka-Zugaj, W.; Litwińczuk, Z.; Chabuz, W.; Šveistienė, R.; Bulla, J. Polymorphism of the Melanocortin 1 Receptor (MC1R) Gene and its Role in Determining the Coat Colour of Central European Cattle Breeds. Animals 2020, 10, 1878. https://doi.org/10.3390/ani10101878
Kasprzak-Filipek K, Sawicka-Zugaj W, Litwińczuk Z, Chabuz W, Šveistienė R, Bulla J. Polymorphism of the Melanocortin 1 Receptor (MC1R) Gene and its Role in Determining the Coat Colour of Central European Cattle Breeds. Animals. 2020; 10(10):1878. https://doi.org/10.3390/ani10101878
Chicago/Turabian StyleKasprzak-Filipek, Karolina, Wioletta Sawicka-Zugaj, Zygmunt Litwińczuk, Witold Chabuz, Rūta Šveistienė, and Josef Bulla. 2020. "Polymorphism of the Melanocortin 1 Receptor (MC1R) Gene and its Role in Determining the Coat Colour of Central European Cattle Breeds" Animals 10, no. 10: 1878. https://doi.org/10.3390/ani10101878
APA StyleKasprzak-Filipek, K., Sawicka-Zugaj, W., Litwińczuk, Z., Chabuz, W., Šveistienė, R., & Bulla, J. (2020). Polymorphism of the Melanocortin 1 Receptor (MC1R) Gene and its Role in Determining the Coat Colour of Central European Cattle Breeds. Animals, 10(10), 1878. https://doi.org/10.3390/ani10101878