Targeting the Hindgut to Improve Health and Performance in Cattle

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
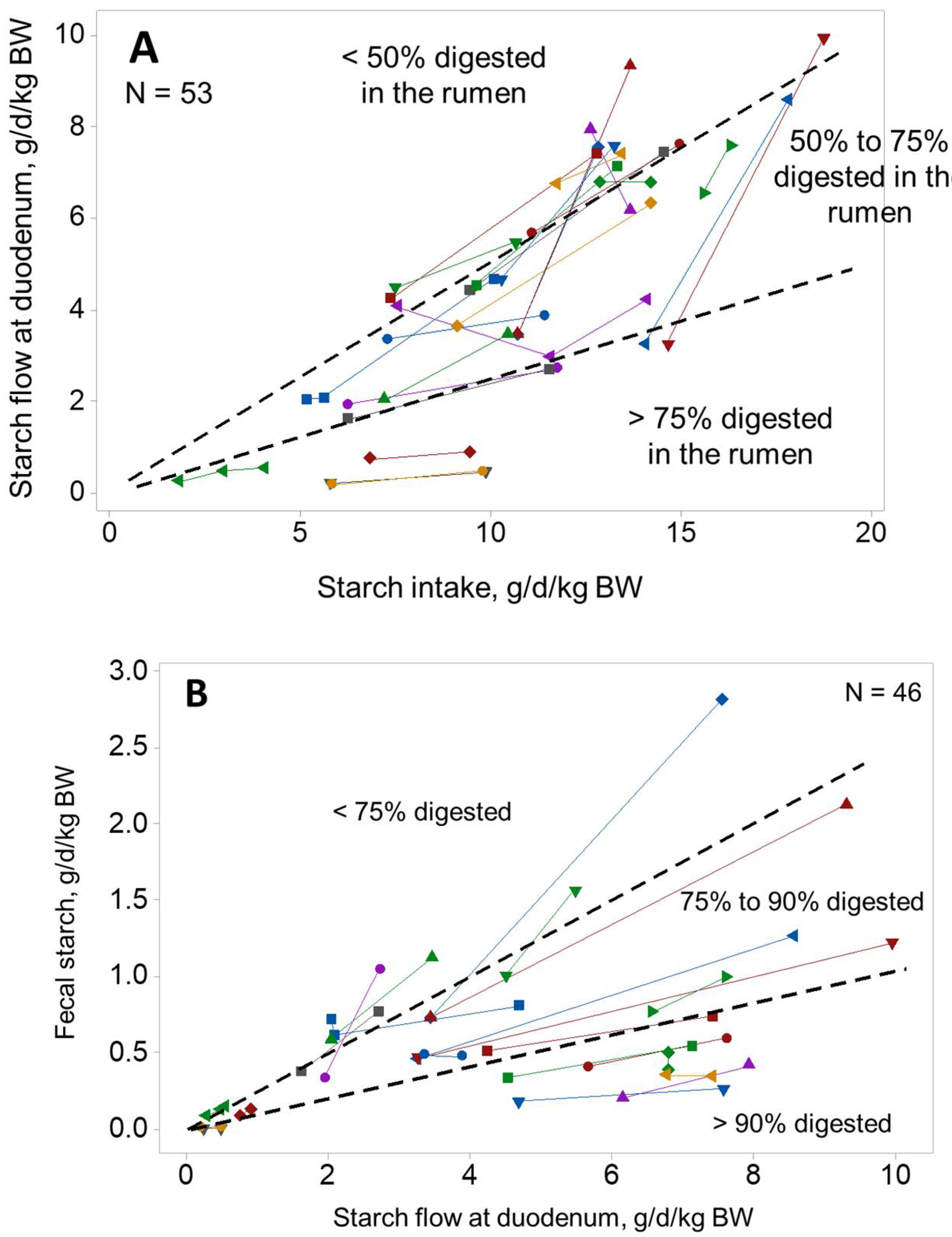
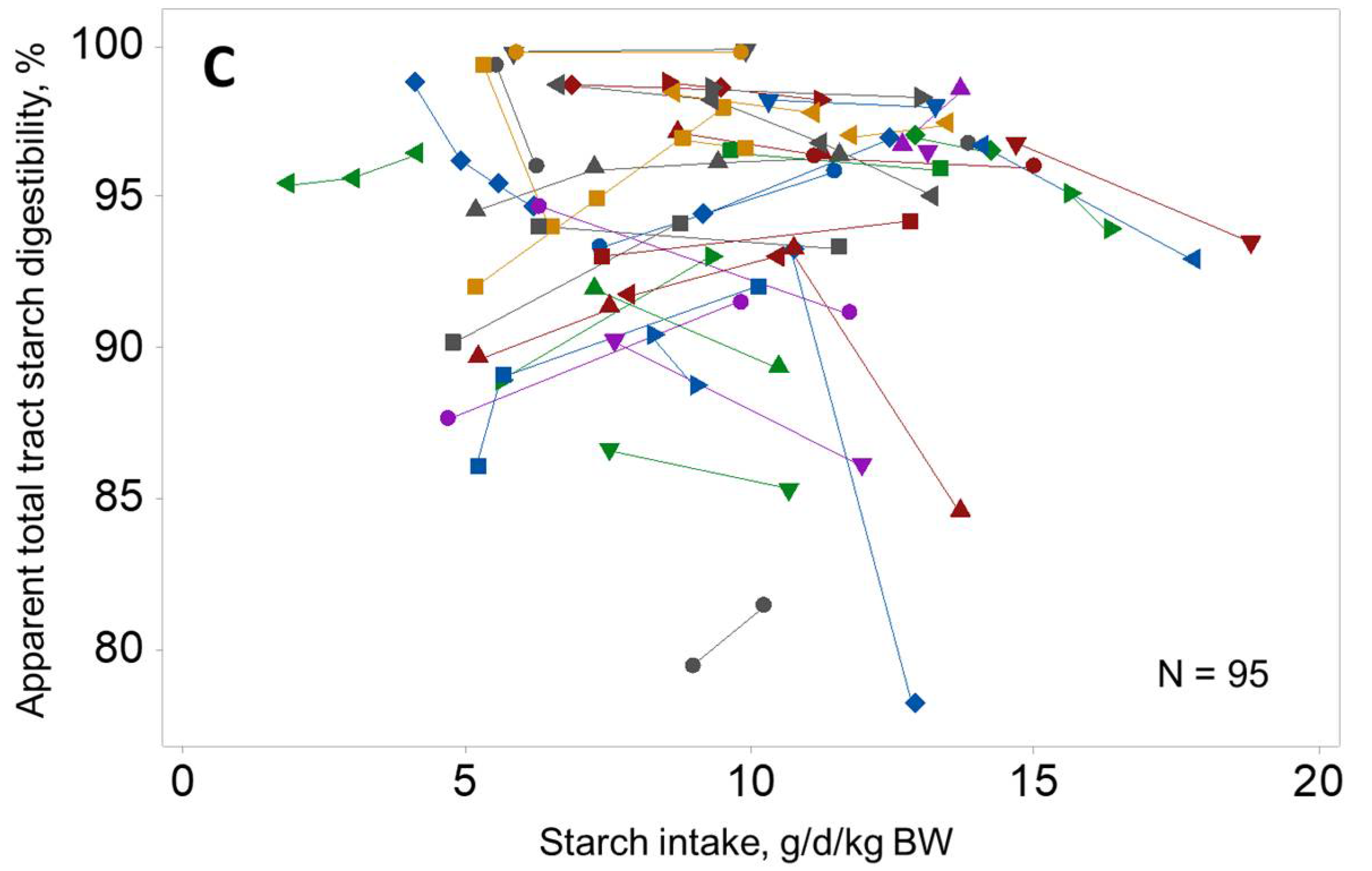
2. Grain Overload Syndrome and Considerations on Starch Digestion Site
3. Intestinal Health and Inflammation
4. Energetic Cost of Inflammation
5. Role of Hindgut Health on Metabolic Diseases in Dairy Cows
6. Role of Hindgut Health on Liver Abscesses in Beef Cattle
7. Nutritional Strategies to Support Hindgut Health
8. Conclusions and Implications
Author Contributions
Funding
Conflicts of Interest
References
- McBride, B.W.; Kelly, J.M. Energy cost of absorption and metabolism in the ruminant gastrointestinal tract and liver: A review. J. Anim. Sci. 1990, 68, 2997–3010. [Google Scholar] [CrossRef]
- Cant, J.P.; McBride, B.W.; Croom, W.J., Jr. The regulation of intestinal metabolism and its impact on whole animal energetics. J. Anim. Sci. 1996, 74, 2541–2553. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, P.J.; Stok, W.; Bianchi, A.T.J. Contribution of immunoglobulin-secreting cells in the murine small intestine to the total ‘background’ immunoglobulin production. Immunology 1987, 62, 551–555. [Google Scholar] [PubMed]
- Matsunaga, T.; Rahman, A. What brought the adaptive immune system to vertebrates?—The jaw hypothesis and the seahorse. Immunol. Rev. 1998, 166, 177–186. [Google Scholar] [CrossRef]
- Bischoff, S.C. ‘Gut health’: A new objective in medicine? BMC Med. 2011, 9, 24. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Sanz Fernandez, M.V.; Abuajamieh, M.; Ganesan, S.; Gorden, P.J.; Green, H.B.; Schoenberg, K.M.; Trout, W.E.; Keating, A.F.; et al. Characterizing effects of feed restriction and glucagon-like peptide 2 administration on biomarkers of inflammation and intestinal morphology. J. Dairy Sci. 2017, 100, 9402–9417. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Albornoz, R.I.; Aschenbach, J.R.; Barreda, D.R.; Penner, G.B. Short-term feed restriction impairs the absorptive function of the reticulo-rumen and total tract barrier function in beef cattle. J. Anim. Sci. 2013, 91, 1685–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanuytsel, T.; van Wanrooy, S.; Vanheel, H.; Vanormelingen, C.; Verschueren, S.; Houben, E.; Salim Rasoel, S.; Tόth, J.; Holvoet, L.; Farré, R.; et al. Psychological stress and corticotropin-releasing hormone increase intestinal permeability in humans by a mast cell-dependent mechanism. Gut 2014, 63, 1293–1299. [Google Scholar] [CrossRef]
- de Punder, K.; Pruimboom, L. Stress induces endotoxemia and low-grade inflammation by increasing barrier permeability. Front. Immunol. 2015, 6, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, T.R.; Smith, R.J.; O’Dwyer, S.T.; Demling, R.H.; Wilmore, D.W. Increased intestinal permeability associated with infection in burn patients. Arch. Surg. 1988, 123, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Hietbrink, F.; Besselink, M.G.; Renooij, W.; de Smet, M.B.; Draisma, A.; van der Hoeven, H.; Pickkers, P. Systemic inflammation increases intestinal permeability during experimental human endotoxemia. Shock 2009, 32, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Danesh Mesgaran, M.; Derakhshani, H.; Golder, H.; Khafipour, E.; Kleen, J.L.; Lean, I.; Loor, J.; Penner, G.B.; Zebeli, Q. Review: Enhancing gastrointestinal health in dairy cows. Animal 2018, 12, s399–s418. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.A.; Penner, G.B.; Chaucheyras-Durand, F.; Guan, L.L. Development and physiology of the rumen and the lower gut: Targets for improving gut health. J. Dairy Sci. 2016, 99, 4955–4966. [Google Scholar] [CrossRef] [Green Version]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.A.; Sanz Fernandez, M.V.; Baumgard, L.H. Glucose requirements of an activated immune system in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2360–2374. [Google Scholar] [CrossRef] [Green Version]
- Eckel, E.F.; Ametaj, B.N. Invited review: Role of bacterial endotoxins in the etiopathogenesis of periparturient diseases of transition dairy cows. J. Dairy Sci. 2016, 99, 5967–5990. [Google Scholar] [CrossRef]
- Gressley, T.F.; Hall, M.B.; Armentano, L.E. Ruminant Nutrition Symposium: Productivity, digestion, and health responses to hindgut acidosis in ruminants. J. Anim. Sci. 2011, 89, 1120–1130. [Google Scholar] [CrossRef] [Green Version]
- Kreikemeier, K.K.; Harmon, D.L.; Brandt, R.T., Jr.; Nagaraja, T.G.; Cochran, R.C. Steam-rolled wheat diets for finishing cattle: Effects of dietary roughage and feed intake on finishing steer performance and ruminal metabolism. J. Anim. Sci. 1990, 68, 2130–2141. [Google Scholar] [CrossRef] [Green Version]
- Swingle, R.S.; Eck, T.P.; Theurer, C.B.; De la Llata, M.; Poore, M.H.; Moore, J.A. Flake density of steam-processed sorghum grain alters performance and sites of digestibility by growing-finishing steers. J. Anim. Sci. 1999, 77, 1055–1065. [Google Scholar] [CrossRef]
- Galyean, M.L.; Wagner, D.G.; Johnson, R.R. Site and Extent of Starch Digestion in Steers Fed Processed Corn Rations. J. Anim. Sci. 1976, 43, 1088–1094. [Google Scholar] [CrossRef]
- Armstrong, D.A.; Blaxter, K.L.; Graham, N.M. Fat synthesis from glucose by sheep. Proc. Nutr. Soc. 1960, 19, xxxi–xxxii. [Google Scholar]
- Owens, F.N.; Zinn, R.A.; Kim, Y.K. Limits to starch digestion in the ruminant small intestine. J. Anim. Sci. 1986, 63, 1634–1648. [Google Scholar] [CrossRef] [PubMed]
- McDonald, P.; Edwards, R.A.; Greenhalgh, J.F.D.; Morgan, C.A. Animal Nutrition, 5th ed.; Longman Scientific & Technical: Harlow, UK, 1995. [Google Scholar]
- Kreikemeier, K.K.; Harmon, D.L.; Brandt, R.T.; Avery, T.B.; Johnson, D.E. Small intestinal starch digestion in steers: Effect of various levels of abomasal glucose, corn starch and corn dextrin infusion on small intestinal disappearance and net glucose absorption. J. Anim. Sci. 1991, 69, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Offner, A.; Sauvant, D. Prediction of in vivo starch digestion in cattle from in situ data. Anim. Feed Sci. Technol. 2004, 111, 41–56. [Google Scholar] [CrossRef]
- Walker, J.A.; Harmon, D.L. Influence of ruminal or abomasal starch hydrolysate infusion on pancreatic exocrine secretion and blood glucose and insulin concentrations in steers. J. Anim. Sci. 1995, 73, 3766–3774. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.C.; Matthews, J.C.; Woods, C.A.; Harmon, D.L. Postruminal administration of partially hydrolyzed starch and casein influences pancreatic alpha-amylase expression in calves. J. Nutr. 2002, 132, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2017, 90, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Nagaraja, T.G.; Bartley, E.E.; Fina, L.R.; Anthony, H.D. Relationship of rumen gram-negative bacteria and free endotoxin to lactic acidosis in cattle. J. Anim. Sci. 1978, 47, 1329–1336. [Google Scholar] [CrossRef]
- Gozho, G.N.; Plaizier, J.C.; Krause, D.O.; Kennedy, A.D.; Wittenberg, K.M. Subacute ruminal acidosis induces ruminal lipopolysaccharide endotoxin release and triggers an inflammatory response. J. Dairy Sci. 2005, 88, 1399–1403. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Khafipour, E.; Krause, D.O.; Kroeker, A.; Rodriguez-Lecompte, J.C.; Gozho, G.N.; Plaizier, J.C. Effects of subacute ruminal acidosis challenges on fermentation and endotoxins in the rumen and hindgut of dairy cows. J. Dairy Sci. 2012, 95, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Zhang, R.; Wang, D.; Zhu, W. The diversity of the fecal bacterial community and its relationship with the concentration of volatile fatty acids in the feces during subacute rumen acidosis in dairy cows. BMC Vet. Res. 2012, 8, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, S.Y.; Zhang, R.Y.; Wang, D.S.; Zhu, W.Y. Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 2013, 24, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Plaizier, J.C.; Li, S.; Tun, H.M.; Khafipour, E. Nutritional models of experimentally-induced subacute ruminal acidosis (SARA) differ in their impact on rumen and hindgut bacterial communities in dairy cows. Front. Microbiol. 2017, 7, 2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Luo, Y.; Wang, H.; Liu, S.; Shen, Y.; Wang, M. Effects of glucose and starch on lactate production by newly isolated Streptococcus bovis S1 from Saanen Goats. Appl. Environ. Microbiol. 2016, 82, 5982–5989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Shen, Y.; Wang, C.; Ding, L.; Zhao, F.; Wang, M.; Fu, J.; Wang, H. Megasphaera elsdenii lactate degradation pattern shifts in rumen acidosis models. Front. Microbiol. 2019, 10, 162. [Google Scholar] [CrossRef] [Green Version]
- McNeil, N.I.; Ling, K.L.; Wager, J. Mucosal surface pH of the large intestine of the rat and of normal and inflamed large intestine in man. Gut 1987, 28, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Aschenbach, J.R.; Penner, G.B.; Stumpff, F.; Gäbel, G. Ruminant Nutrition Symposium: Role of fermentation acid absorption in the regulation of ruminal pH. J. Anim. Sci. 2011, 89, 1092–1107. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, E.; Neves, A.L.; Song, Y.; Guan, L.L. The role of the gut microbiome in cattle production and health: Driver or passenger? Annu. Rev. Anim. Biosci. 2020, 8, 199–220. [Google Scholar] [CrossRef] [Green Version]
- Emmanuel, D.G.V.; Madsen, K.L.; Churchill, T.A.; Dunn, S.M.; Ametaj, B.N. Acidosis and lipopolysaccharide from Escherichia coli B:055 cause hyperpermeability of rumen and colon tissues. J. Dairy Sci. 2007, 90, 5552–5557. [Google Scholar] [CrossRef] [Green Version]
- Minuti, A.; Ahmed, S.; Trevisi, E.; Piccioli-Cappelli, F.; Bertoni, G.; Jahan, N.; Bani, P. Experimental acute rumen acidosis in sheep: Consequences on clinical, rumen, and gastrointestinal permeability conditions and blood chemistry. J. Anim. Sci. 2014, 92, 3966–3977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, M.A.; AlZahal, O.; Hook, S.E.; Croom, J.; McBride, B.W. Ruminal acidosis and the rapid onset of ruminal parakeratosis in a mature dairy cow: A case report. Acta Vet. Scand. 2009, 51, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Duanmu, Y.; Dong, H.; Tian, J.; Ni, Y.; Zhao, R. A high-concentrate diet induced colonic epithelial barrier disruption is associated with the activating of cell apoptosis in lactating goats. BMC Vet. Res. 2014, 10, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Duanmu, Y.; Dong, H.; Ni, Y.; Chen, J.; Shen, X.; Zhao, R. High concentrate diet induced mucosal injuries by enhancing epithelial apoptosis and inflammatory response in the hindgut of goats. PLoS ONE 2014, 9, e111596. [Google Scholar] [CrossRef] [Green Version]
- Garry, F.B. Indigestion in ruminants. In Large Animal Internal Medicine, Mosby-Year Book; Smith, B.P., Ed.; Mosby: St. Louis, MO, USA, 2002; pp. 722–747. [Google Scholar]
- Zebeli, Q.; Ametaj, B.N. Relationships between rumen lipopolysaccharide and mediators of inflammatory response with milk fat production and efficiency in dairy cows. J. Dairy Sci. 2009, 92, 3800–3809. [Google Scholar] [CrossRef] [Green Version]
- Plaizier, J.C.; Khafipour, E.; Li, S.; Gozho, G.N.; Krause, D.O. Subacute ruminal acidosis (SARA), endotoxins and health consequences. Anim. Feed Sci, Techno. 2012, 172, 9–21. [Google Scholar] [CrossRef]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef] [Green Version]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. Alfalfa pellet-induced subacute ruminal acidosis in dairy cows increases bacterial endotoxin in the rumen without causing inflammation. J. Dairy Sci. 2009, 92, 1712–1724. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Fernandez, M.V.; Ilg, T.; Daniel, J.B.; Martin-Tereso, J.; Mertens, C.; Doelman, J.H. Characterization of a model of leaky gut secondary to hindgut acidosis in mid-lactation cows: A pilot study. In Proceedings of the ADSA Discover Conference, Ithasca, IL, USA, 28–31 October 2019. [Google Scholar]
- Mainardi, S.R.; Hengst, B.A.; Nebzydoski, S.J.; Nemec, L.M.; Gressley, T.F. Effects of abomasal oligofructose on blood and feces of Holstein steers. J. Anim Sci. 2011, 89, 2510–2517. [Google Scholar] [CrossRef]
- Bissell, H.A. Post-ruminal starch infusion in dairy cattle: Implications for inflammatory response and animal health. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2002. [Google Scholar]
- Boosman, R.; Mutsaers, C.W.; Klarenbeek, A. The role of endotoxin in the pathogenesis of acute bovine laminitis. Vet. Q. 1991, 13, 155–162. [Google Scholar] [CrossRef]
- Geor, R.J. Current concepts on the pathophysiology of pasture-associated laminitis. Vet. Clin. North. Am. Equine Pract. 2010, 26, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Suagee-Bedore, J.K.; Linden, D.R.; Bennett-Wimbush, K.; Splan, R.K. Feeding grass hay before concentrate mitigates the effect of grain-based concentrates on postprandial plasma interleukin-1β. J. Equine Vet. Sci. 2020, 86, 102899. [Google Scholar] [CrossRef] [PubMed]
- Lochmiller, R.L.; Deerenberg, C. Trade-offs in evolutionary immunology: Just what is the cost of immunity? Oikos 2000, 88, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Graves, R.J. Lecture IV. In Clinical Lectures, 2nd ed.; Barrington & Haswell: San Francisco, CA, USA, 1842; pp. 278–285. [Google Scholar]
- Calder, P.C.; Dimitriadis, G.; Newsholme, P. Glucose metabolism in lymphoid and inflammatory cells and tissues. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 531–540. [Google Scholar] [CrossRef]
- MacIver, N.J.; Jacobs, S.R.; Wieman, H.L.; Wofford, J.A.; Coloff, J.L.; Rathmell, J.C. Glucose metabolism in lymphocytes is a regulated process with significant effects on immune cell function and survival. J. Leukoc. Biol. 2008, 84, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Palsson-McDermott, E.M.; O’Neill, L.A. The Warburg effect then and now: From cancer to inflammatory diseases. Bioessays 2013, 35, 965–973. [Google Scholar] [CrossRef]
- Lang, C.H.; Dobrescu, C. Sepsis-induced increases in glucose uptake by macrophage-rich tissues persist during hypoglycemia. Metabolism 1991, 40, 585–593. [Google Scholar] [CrossRef]
- Lang, C.H.; Spolarics, Z.; Ottlakan, A.; Spitzer, J.J. Effect of high-dose endotoxin on glucose production and utilization. Metabolism 1993, 42, 1351–1358. [Google Scholar] [CrossRef]
- Plank, L.D.; Connolly, A.B.; Hill, G.L. Sequential changes in the metabolic response in severely septic patients during the first 23 days after the onset of peritonitis. Ann. Surg. 1998, 228, 146–158. [Google Scholar] [CrossRef]
- Mészáros, K.; Bojta, J.; Bautista, A.P.; Lang, C.H.; Spitzer, J.J. Glucose utilization by Kupffer cells, endothelial cells, and granulocytes in endotoxemic rat liver. Am. J. Physiol. 1991, 260, G7-12. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.A.; Sanz Fernandez, M.V.; Baumgard, L.H. Technical note: A procedure to estimate glucose requirements of an activated immune system in steers. J. Anim. Sci. 2016, 94, 4591–4599. [Google Scholar] [CrossRef] [PubMed]
- Kvidera, S.K.; Horst, E.A.; Mayorga, E.A.; Sanz Fernandez, M.V.; Abuajamieh, M.; Baumgard, L.H. Estimating glucose requirements of an activated immune system in growing pigs. J. Anim Sci. 2017, 95, 5020–5029. [Google Scholar] [CrossRef] [PubMed]
- van Engelen, E.; de Groot, M.W.; Breeveld-Dwarkasing, V.N.A.; Everts, M.E.; van der Weyden, G.C.; Taverne, M.A.M.; Rutten, V.P.M.G. Cervical ripening and parturition in cows are driven by a cascade of pro-inflammatory cytokines. Reprod. Domest. Anim. 2009, 44, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Bradford, B.J.; Yuan, K.; Farney, J.K.; Mamedova, L.K.; Carpenter, A.J. Invited review: Inflammation during the transition to lactation: New adventures with an old flame. J. Dairy Sci. 2015, 98, 6631–6650. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Bradford, B.J.; Bobe, G.; Nafikov, R.A.; Lu, Y.; Young, J.W.; Beitz, D.C. Strong relationships between mediators of the acute phase response and fatty liver in dairy cows. Can. J. Anim. Sci. 2005, 85, 65–175. [Google Scholar] [CrossRef]
- Humblet, M.-F.; Guyot, H.; Boudry, B.; Mbayahi, F.; Hanzen, C.; Rollin, F.; Godeau, J.-M. Relationship between haptoglobin, serum amyloid A, and clinical status in a survey of dairy herds during a 6-month period. Vet. Clin. Pathol. 2006, 35, 188–193. [Google Scholar] [CrossRef]
- Bionaz, M.; Trevisi, E.; Calamari, L.; Librandi, F.; Ferrari, A.; Bertoni, G. Plasma paraoxonase, health, inflammatory conditions, and liver function in transition dairy cows. J. Dairy Sci. 2007, 90, 1740–1750. [Google Scholar] [CrossRef] [Green Version]
- Mullins, C.R.; Mamedova, L.K.; Brouk, M.J.; Moore, C.E.; Green, H.B.; Perfield, K.L.; Smith, J.F.; Harner, J.P.; Bradford, B.J. Effects of monensin on metabolic parameters, feeding behavior, and productivity of transition dairy cows. J. Dairy Sci. 2012, 95, 1323–1336. [Google Scholar] [CrossRef] [Green Version]
- Huzzey, J.M.; Duffield, T.F.; LeBlanc, S.J.; Veira, D.M.; Weary, D.M.; von Keyserlingk, M.A.G. Short communication: Haptoglobin as an early indicator of metritis. J. Dairy Sci. 2009, 92, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Goldansaz, S.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Alterations in innate immunity reactants and carbohydrate and lipid metabolism precede occurrence of metritis in transition dairy cows. Res. Vet. Sci. 2016, 104, 30–39. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Dunn, S.M.; Ametaj, B.N. Innate immunity and carbohydrate metabolism alterations precede occurrence of subclinical mastitis in transition dairy cows. J. Anim. Sci. Technol. 2015, 57, 46–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Deng, Q.; Goldansaz, S.A.; Dunn, S.M.; Ametaj, B.N. Alterations of innate immunity reactants in transition dairy cows before clinical signs of lameness. Animals 2015, 5, 717–747. [Google Scholar] [CrossRef] [PubMed]
- Abuajamieh, M.; Stoakes, S.K.; Sanz Fernandez, M.V.; Nayeri, A.; Upah, N.C.; Nolan, E.A.; Lei, S.M.; DeFrain, J.M.; Green, H.B.; Schoenberg, K.M.; et al. Inflammatory biomarkers are closely associated with ketosis in periparturient Holstein cows. Res. Vet. Sci. 2016, 109, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Dairy cows affected by ketosis show alterations in innate immunity and lipid and carbohydrate metabolism during the dry off period and postpartum. Res. Vet. Sci. 2016, 107, 246–256. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Dunn, S.M.; Ametaj, B.N. Occurrence of retained placenta is preceded by an inflammatory state and alterations of energy metabolism in transition dairy cows. J. Anim. Sci. Biotechnol. 2016, 7, 6–39. [Google Scholar] [CrossRef] [Green Version]
- Huber, K.; Dänicke, S.; Rehage, J.; Sauerwein, H.; Otto, W.; Rolle-Kampczyk, U.; Von Bergen, M. Metabotypes with properly functioning mitochondria and anti-inflammation predict extended productive life span in dairy cows. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Mezzetti, M.; Minuti, A.; Piccioli-Cappelli, F.; Amadori, M.; Bionaz, M.; Trevisi, E. The role of altered immune function during the dry period in promoting the development of subclinical ketosis in early lactation. J. Dairy Sci. 2019, 102, 9241–9258. [Google Scholar] [CrossRef]
- Sanz-Fernandez, M.V.; Pesantez-Pacheco, J.L.; Torres-Rovira, L.; Vazquez-Gomez, M.; Garcia-Contreras, C.; Heras-Molina, A.; Perez Villalobos, N.; Hernandez, F.; Gonzalez-Martin, J.V.; Gonzalez-Bulnes, A.; et al. Gestational toxemia in lactating sheep is associated with alterations in circulating inflammatory biomarkers. In Proceedings of the 30th World Buiatrics Congress, Sapporo, Japan, 28 August–1 September 2018. [Google Scholar]
- Nagaraja, T.G.; Laudert, S.B.; Parrott, J.C. Liver abscesses in feedlot cattle. Part 2. Incidence, economic importance and prevention. Comp. Cont. Edu. Pract. Vet. 1996, 18, S264–S273. [Google Scholar]
- Reinhardt, C.D.; Hubbert, M.E. Control of liver abscesses in feedlot cattle: A review. The Prof. Am. Sci. 2015, 31, 101–108. [Google Scholar] [CrossRef]
- Smith, H.R. Beef liver condemnations. J. Anim. Sci. 1940, 1940, 272–276. [Google Scholar]
- Herrick, R.; Rogers, C.; Jones, T.; McEvers, T.; Brown, T.; Maxwell, C.; Lawrence, T. 481 Association of liver abscess presence and severity with trim loss, harvest yield, carcass grading performance, lung lesions, and value of fed Holsteins. J. Anim. Sci. 2018, 96 (Suppl. 3), 269. [Google Scholar] [CrossRef]
- Smith, H.A. Ulcerative lesions of the bovine rumen and their possible relation to hepatic abscesses. Am. J. Vet. Res. 1944, 5, 235–242. [Google Scholar]
- Jensen, R.; Frey, P.R. Telangiectasis, sawdust, and abscesses in the livers of beef cattle. J. Am. Vet. Med. Assoc. 1947, 110, 256–261. [Google Scholar] [PubMed]
- Matsushima, J.; Dowe, T.W.; Adams, C.H. Effect of aureomycin in preventing liver abscess in cattle. Proc. Soc. Exp. Biol. Med. 1954, 85, 18–20. [Google Scholar] [CrossRef]
- Jensen, R.; Deane, H.M.; Cooper, L.J.; Miller, V.A.; Graham, W.R. The rumenitis-liver abscess complex in beef cattle. Am. J. Vet. Res. 1954, 15, 202. [Google Scholar]
- Nagaraja, T.G.; Beharka, A.B.; Chengappa, M.M.; Carroll, L.H.; Raun, A.P.; Laudert, S.B.; Parrott, J.C. Bacterial flora of liver abscesses in feedlot cattle fed tylosin or no tylosin. J. Anim. Sci. 1999, 77, 973–978. [Google Scholar] [CrossRef]
- Brink, D.R.; Lowry, S.R.; Stock, R.A.; Parrott, J.C. Severity of liver abscesses and efficiency of feed utilization of feedlot cattle. J. Anim. Sci. 1990, 68, 1201–1207. [Google Scholar] [CrossRef]
- Owens, F.N.; Secrist, D.S.; Hill, W.J.; Gill, D.R. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef]
- Nagaraja, T.G.; Lechtenberg, K.F. Liver abscesses in feedlot cattle. Vet. Clin. North. Am. Food Anim. Pract. 2007, 23, 333–350. [Google Scholar] [CrossRef]
- Wadhwa, M.; Bakshi, M.P.; Makkar, H.P. Modifying gut microbiomes in large ruminants: Opportunities in non-intensive husbandry systems. Anim. Front. 2016, 6, 27–36. [Google Scholar] [CrossRef]
- Scanlan, C.M.; Hathcock, T.L. Bovine rumenitis-liver abscess complex: A bacteriological review. The Cornell Vet. 1983, 73, 288–297. [Google Scholar] [PubMed]
- Nagaraja, T.G.; Chengappa, M.M. Liver abscesses in feedlot cattle: A review. J. Anim. Sci. 1998, 76, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadepalli, S.; Narayanan, S.K.; Stewart, G.C.; Chengappa, M.M.; Nagaraja, T.G. Fusobacterium necrophorum: A ruminal bacterium that invades liver to cause abscesses in cattle. Anaerobe 2009, 15, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.L.; Nagaraja, T.G.; Chengappa, M.M. Fusobacterium necrophorum infections: Virulence factors, pathogenic mechanism, and control measures. Vet. Res. Commun. 1996, 20, 113–140. [Google Scholar] [CrossRef]
- Narayanan, S.; Nagaraja, T.G.; Wallace, N.; Staats, J.; Chengappa, M.M.; Oberst, R.D. Biochemical and ribotypic comparison of Actinomyces pyogenes and A pyogenes-like organisms from liver abscesses, ruminal wall, and ruminal contents of cattle. Am. J. Vet. Res. 1998, 59, 271–276. [Google Scholar]
- Rasmussen, M.A.; Carlson, S.A.; Franklin, S.K.; McCuddin, Z.P.; Wu, M.T.; Sharma, V.K. Exposure to rumen protozoa leads to enhancement of pathogenicity of and invasion by multiple-antibiotic-resistant Salmonella enterica bearing SGI1. Infect. Immun. 2005, 73, 4668–4675. [Google Scholar] [CrossRef] [Green Version]
- Wieser, M.F.; Preston, T.R.; Macdearmid, A.; Rowland, A.C. Intensive beef production. 8. The effect of chlortetracycline on growth, feed utilisation and incidence of liver abscesses in barley beef cattle. Anim. Sci. 1966, 8, 411–423. [Google Scholar] [CrossRef]
- Bester, Z.; Hubbert, M.E.; Carey, R.E.; Samuelson, K.L.; Loest, C.A. 1671WS Shifting the paradigm of liver abscess dogma in USA feedlots. J. Anim. Sci. 2016, 94 (Suppl. 5), 814. [Google Scholar] [CrossRef]
- Amachawadi, R.G.; Nagaraja, T.G. First report of anaerobic isolation of Salmonella enterica from liver abscesses of feedlot cattle. J. Clin. Microbiol. 2015, 53, 3100–3101. [Google Scholar] [CrossRef] [Green Version]
- Gragg, S.E.; Loneragan, G.H.; Brashears, M.M.; Arthur, T.M.; Bosilevac, J.M.; Kalchayanand, N.; Wang, R.; Schmidt, J.W.; Brooks, J.C.; Shackelford, S.D.; et al. Cross-sectional study examining Salmonella enterica carriage in subiliac lymph nodes of cull and feedlot cattle at harvest. Foodborne Pathog. Dis. 2013, 10, 368–374. [Google Scholar] [CrossRef] [Green Version]
- United States Department of Agriculture (USDA). Part IV: Health and health management on U.S. feedlots with capacity of 1000 or more head. In Feedlot 2011; USDA–APHIS–VS–CEAH–NAHMS: Fort Collins, CO, USA, 2013. [Google Scholar]
- Nagaraja, T.G.; Sun, Y.; Wallace, N.; Kemp, K.E.; Parrott, C.J. Effects of tylosin on concentrations of Fusobacterium necrophorum and fermentation products in the rumen of cattle fed a high-concentrate diet. Am. J. Vet. Res. 1999, 60, 1061–1065. [Google Scholar] [PubMed]
- Elanco. Elanco Liver Check System; Elanco Animal Health: Greenfield, IN, USA, 2014. [Google Scholar]
- Brown, T.R.; Lawrence, T.E. Association of liver abnormalities with carcass grading performance and value. J. Anim. Sci. 2010, 88, 4037–4043. [Google Scholar] [CrossRef] [PubMed]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef] [PubMed]
- Hills, R.D.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut microbiome: Profound implications for diet and disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, S.C.; Sanz Fernandez, M.V.; Torrison, J.; Wilson, M.E.; Baumgard, L.H.; Gabler, N.K. Dietary organic zinc attenuates heat stress–induced changes in pig intestinal integrity and metabolism. J. Anim. Sci. 2015, 93, 4702–4713. [Google Scholar] [CrossRef] [PubMed]
- Abuajamieh, M.; DeFrain, J.; Rhoads, R.P.; Baumgard, L.H.; Kvidera, S.K.; Horst, E.A.; Mayorga, E.J.; Seibert, J.T.; Johnson, J.S.; Ross, J.W.; et al. The effects of zinc amino acid complex on biomarkers of gut integrity and metabolism in heat-stressed steers. J. Anim. Sci. 2016, 94 (Suppl. 5), 564. [Google Scholar] [CrossRef]
- Spears, J.W. Zinc methionine for ruminants: Relative bioavailability of zinc in lambs and effects of growth and performance of growing heifers. J. Anim. Sci. 1989, 67, 835–843. [Google Scholar] [CrossRef]
- Shaeffer, G.L.; Lloyd, K.E.; Spears, J.W. Bioavailability of zinc hydroxychloride relative to zinc sulfate in growing cattle fed a corn-cottonseed hull-based diet. Anim. Feed Sci. Technol. 2017, 232, 1–5. [Google Scholar] [CrossRef]
- De Lange, C.F.; Pluske, J.; Gong, J.; Nyachoti, C.M. Strategic use of feed ingredients and feed additives to stimulate gut health and development in young pigs. Livest. Sci. 2010, 134, 124–134. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Updating the concept of prebiotics. Nutr. Res. Rev. 2004, 17, 259–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, H.L.; Campbell, B.J. Dietary fibre–microbiota interactions. Aliment. Pharmacol. Ther. 2015, 42, 158–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamer, H.M.; Jonkers, D.M.; Venema, K.; Vanhoutvin, S.A.; Troost, F.J.; Brummer, R.J. The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, N.B.; Harmon, D.L. Effect of increasing ruminal butyrate absorption on splanchnic metabolism of volatile fatty acids absorbed from the washed reticulorumen of steers. J. Anim. Sci. 2004, 82, 3549–3559. [Google Scholar] [CrossRef]
- Uyeno, Y.; Shigemori, S.; Shimosato, T. Effect of probiotics/prebiotics on cattle health and productivity. Microbes Environ. 2015, 30, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Tsukahara, T.; Koyama, H.; Okada, M.; Ushida, K. Stimulation of butyrate production by gluconic acid in batch culture of pig cecal digesta and identification of butyrate-producing bacteria. J. Nutr. 2002, 132, 2229–2234. [Google Scholar] [CrossRef]
- Tsukahara, T.; Hashizume, K.; Koyama, H.; Ushida, K. Stimulation of butyrate production through the metabolic interaction among lactic acid bacteria, Lactobacillus acidophilus, and lactic acid--utilizing bacteria, Megasphaera elsdenii, in porcine cecal digesta. Anim. Sci. J. 2006, 77, 454–461. [Google Scholar] [CrossRef]
- Biagi, G.; Piva, A.; Moschini, M.; Vezzali, E.; Roth, F.X. Effect of gluconic acid on piglet growth performance, intestinal microflora, and intestinal wall morphology. J. Anim. Sci. 2006, 84, 370–378. [Google Scholar] [CrossRef]
- Poeikhampha, T.; Bunchasak, C. Comparative effects of sodium gluconate, mannan oligosaccharide and potassium diformate on growth performances and small intestinal morphology of nursery pigs. Asian-Australas. J. Anim. Sci. 2011, 24, 844–850. [Google Scholar] [CrossRef]
- Cañete-Rodriguez, A.M.; Santos-Duenas, I.M.; Jimenez-Hornero, J.E.; Ehrenreich, A.; Liebl, W.; Garcia-Garcia, I. Gluconic acid: Properties, production methods and applications—An excellent opportunity for agro-industrial by-products and waste bio-valorization. Process. Biochem. 2016, 51, 1891–1903. [Google Scholar]
- Asano, T.; Yuasa, K.; Kunugita, K.; Teraji, T.; Mitsuoka, T. Effects of gluconic acid on human faecal bacteria. Microb. Ecol. Health Dis. 1994, 7, 247–256. [Google Scholar]
- Sokatch, J.T.; Gunsalus, I.C. Aldonic acid metabolism. I. Pathway of carbon in an inducible gluconate fermentation by Streptococcus faecalis. J. Bacteriol. 1957, 73, 452–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kameue, C.; Tsukahara, T.; Yamada, K.; Koyama, H.; Iwasaki, Y.; Nakayama, K.; Ushida, K. Dietary sodium gluconate protects rats from large bowel cancer by stimulating butyrate production. J. Nutr. 2004, 134, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Michiels, J.; Truffin, D.; Majdeddin, M.; Van Liefferinge, E.; Van Noten, N.; Vandaele, M.; Van Kerschaver, C.; Degroote, J.; Linder, P. L’acide gluconique améliore les performances des porcelets nouvellement sevrés associées avec des modifications au niveau du microbiote intestinal et de la fermentation. In Proceedings of the 52emes Journées de la Recherche Porcine, Paris, France, 4–5 February 2020; pp. 177–178. [Google Scholar]
- McKnight, L.L.; Doelman, J.; Carson, M.; Waterman, D.F.; Metcalf, J.A. Feeding and postruminal infusion of calcium gluconate to lactating dairy cows. Can. J. Anim. Sci. 2019, 99, 563–569. [Google Scholar] [CrossRef] [Green Version]
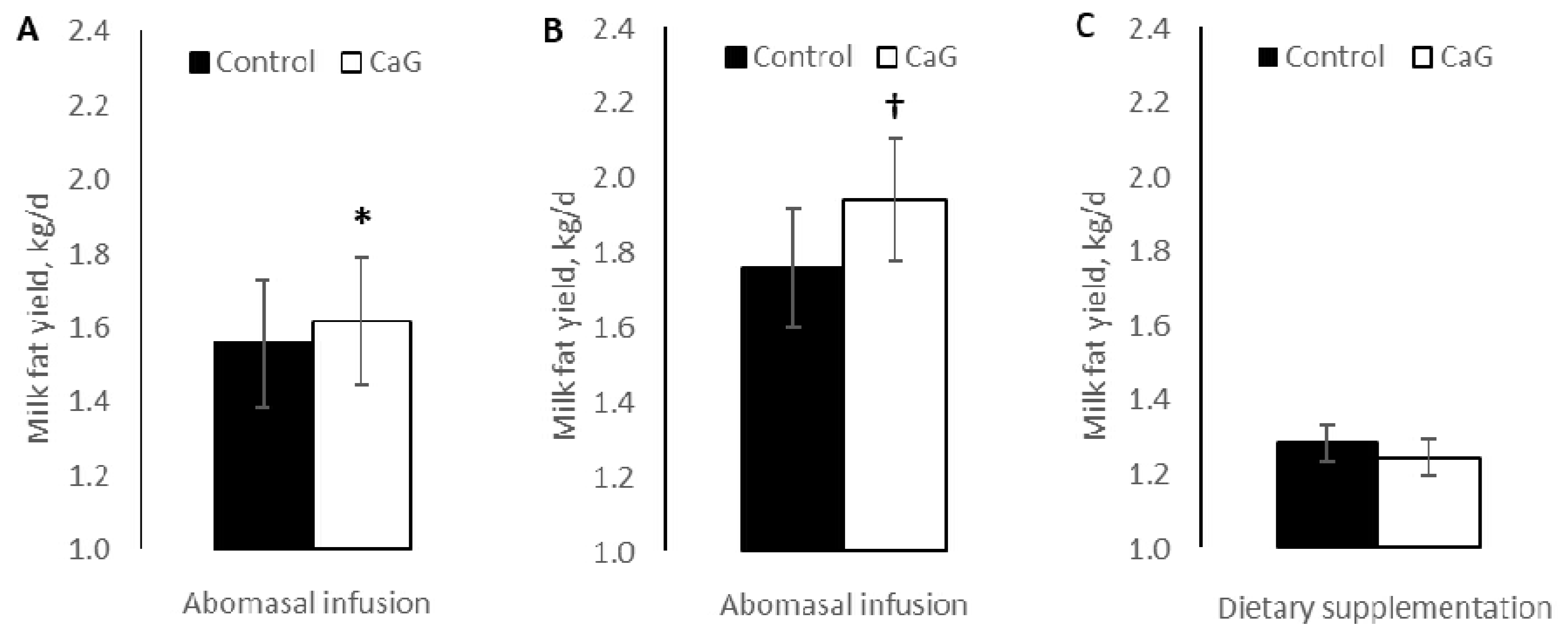
- Doelman, J.; McKnight, L.L.; Carson, M.; Nichols, K.; Waterman, D.F.; Metcalf, J.A. Postruminal infusion of calcium gluconate increases milk fat production and alters fecal volatile fatty acid profile in lactating dairy cows. J. Dairy Sci. 2019, 102, 1274–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, D.J.; Daniel, J.B.; Martín-Tereso, J.; Doelman, J. Effect of fat-embedded calcium gluconate on lactation performance and metabolism in dairy cattle. J. Dairy. Sci. 2020, 103 (Suppl. 1), 70–71. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Gluconate Affinity 2 |
|---|---|
| Bifidobacterium | |
| adolescentis | ++ |
| pseudocatenulatum | +++ |
| catenulatum | +++ |
| dentium | +++ |
| Clostridium | |
| clostridifome | + |
| innocuum | + |
| Cutibacterium acnes3 | + |
| Enterococcus faecium | ++ |
| Klebsiella pneumoniae | + |
| Megasphaera elsdenii | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanz-Fernandez, M.V.; Daniel, J.-B.; Seymour, D.J.; Kvidera, S.K.; Bester, Z.; Doelman, J.; Martín-Tereso, J. Targeting the Hindgut to Improve Health and Performance in Cattle. Animals 2020, 10, 1817. https://doi.org/10.3390/ani10101817
Sanz-Fernandez MV, Daniel J-B, Seymour DJ, Kvidera SK, Bester Z, Doelman J, Martín-Tereso J. Targeting the Hindgut to Improve Health and Performance in Cattle. Animals. 2020; 10(10):1817. https://doi.org/10.3390/ani10101817
Chicago/Turabian StyleSanz-Fernandez, M. Victoria, Jean-Baptiste Daniel, Dave J. Seymour, Sara K. Kvidera, Zeno Bester, John Doelman, and Javier Martín-Tereso. 2020. "Targeting the Hindgut to Improve Health and Performance in Cattle" Animals 10, no. 10: 1817. https://doi.org/10.3390/ani10101817
APA StyleSanz-Fernandez, M. V., Daniel, J.-B., Seymour, D. J., Kvidera, S. K., Bester, Z., Doelman, J., & Martín-Tereso, J. (2020). Targeting the Hindgut to Improve Health and Performance in Cattle. Animals, 10(10), 1817. https://doi.org/10.3390/ani10101817