Curcumin Alleviates IUGR Jejunum Damage by Increasing Antioxidant Capacity through Nrf2/Keap1 Pathway in Growing Pigs

 ,
, 

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animal and Experimental Design
2.3. Assay of Antioxidant Index in the Jejunal Mucosa
2.4. Total RNA Isolation and mRNA Quantification
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
3.1. Antioxidant Index in the Jejunal Mucosa
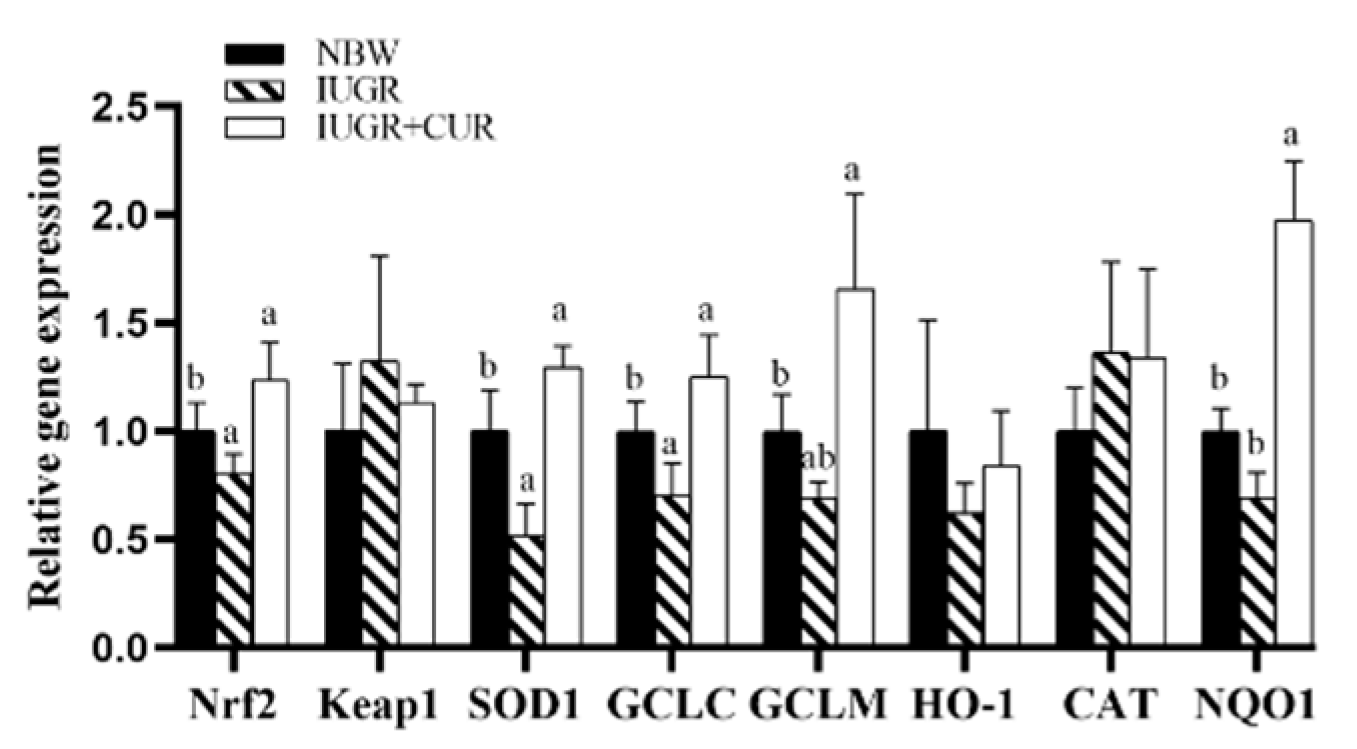
3.2. Antioxidant Enzyme Gene Expression in the Jejunal Mucosa
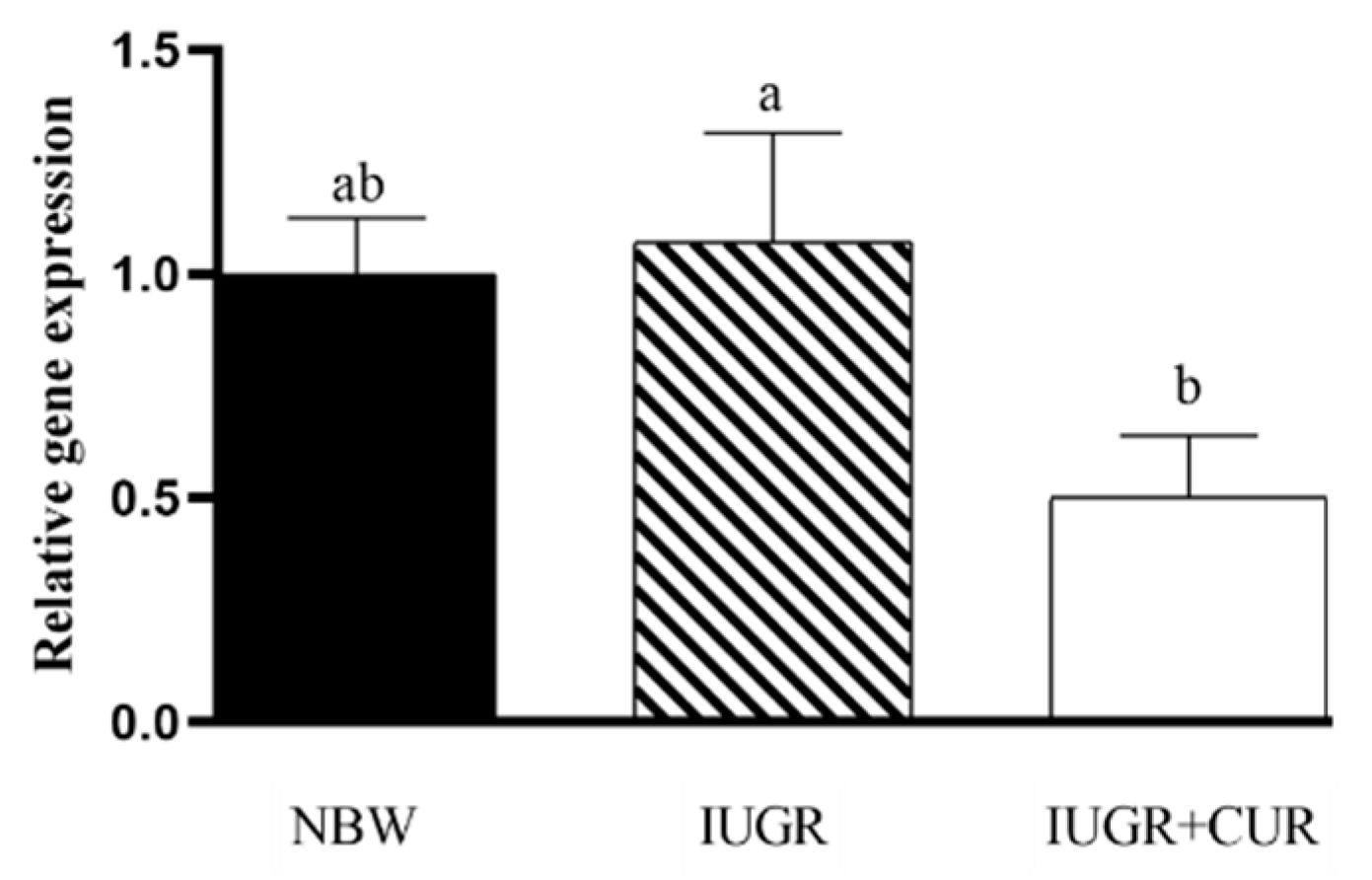
3.3. Hsp70 Expression in the Jejunal Mucosa
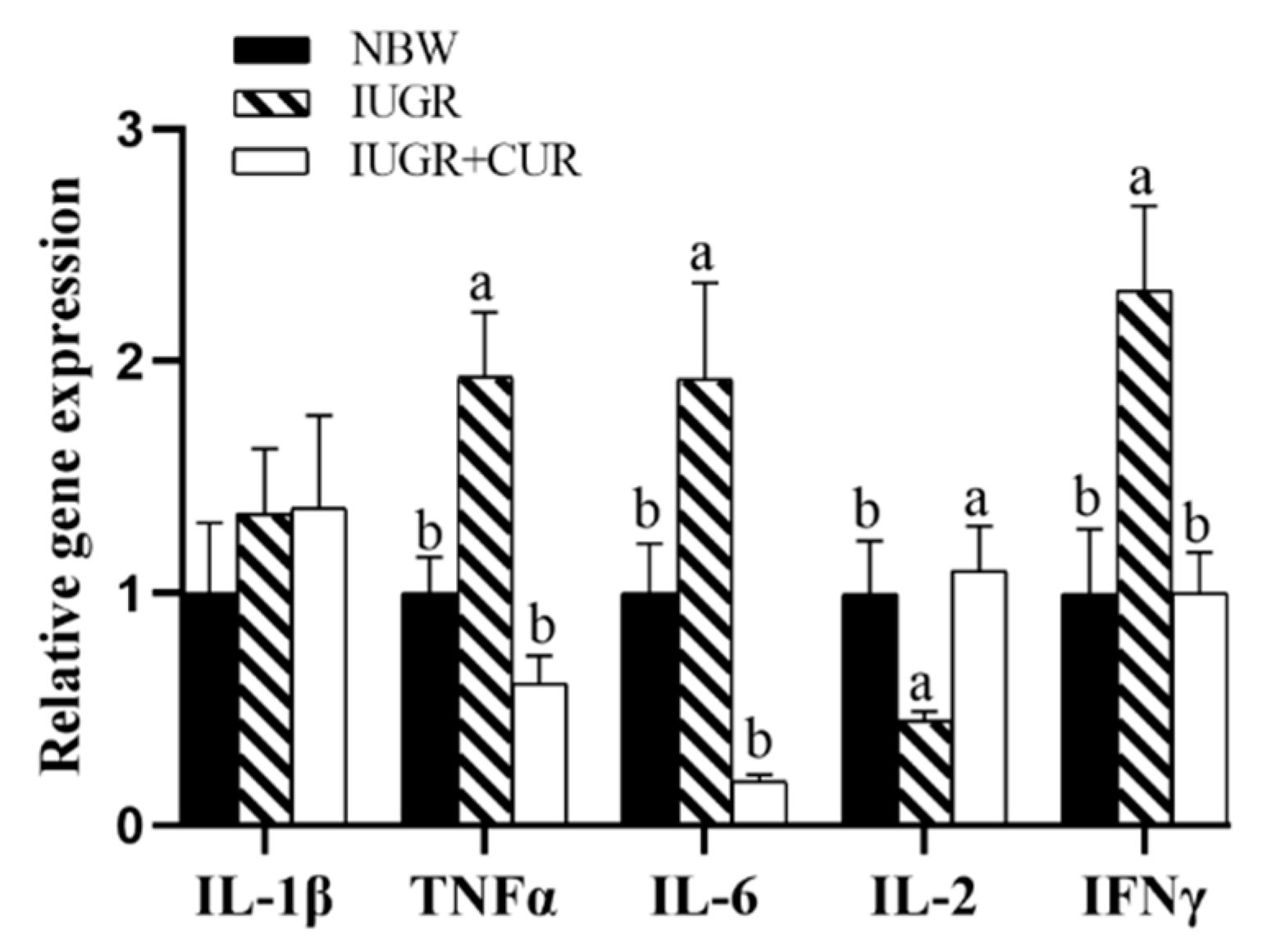
3.4. Immune-Related Gene Expression in the Jejunal Mucosa
3.5. Apoptosi- Related Gene Expression in the Jejunal Mucosa
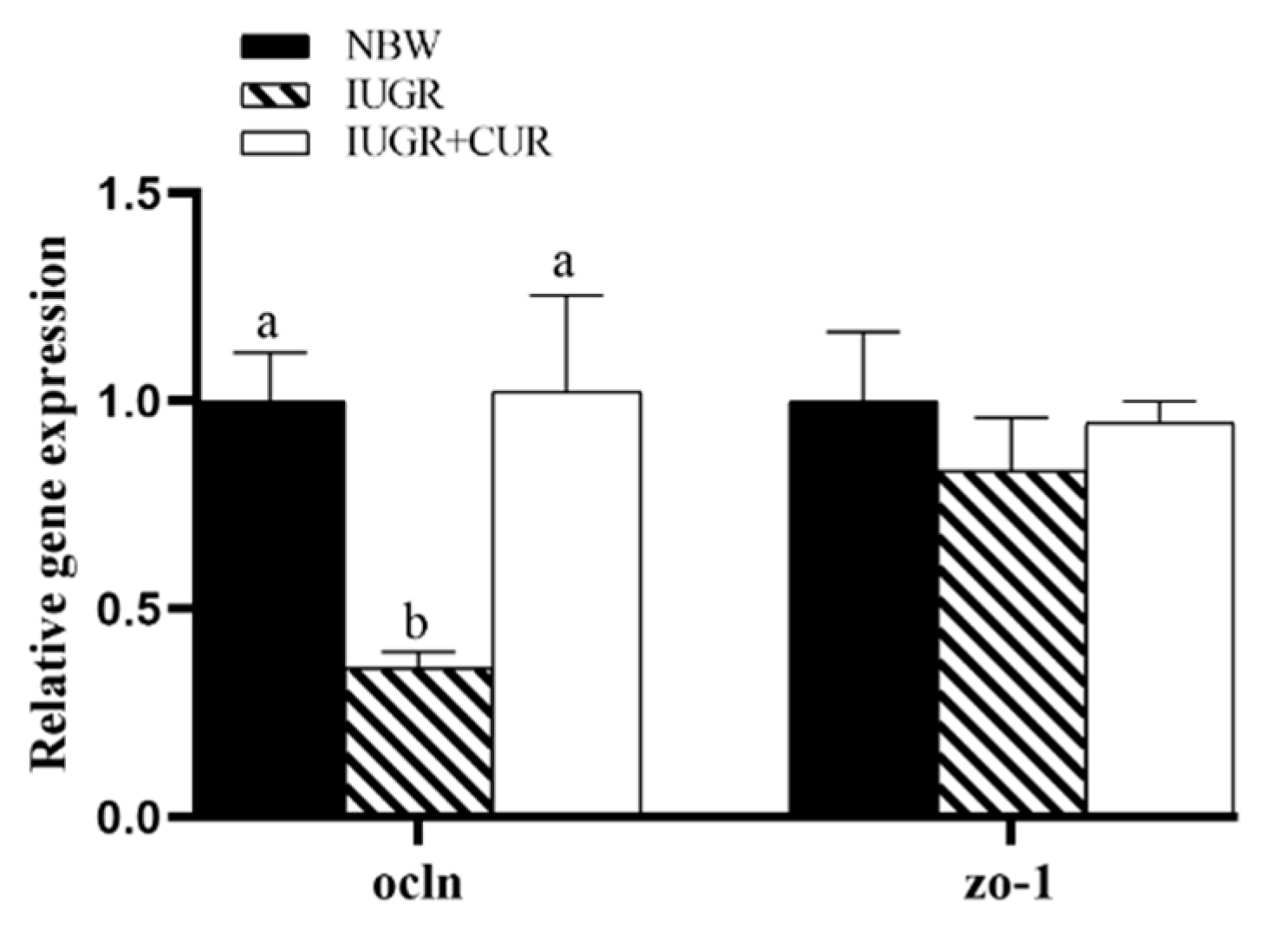
3.6. Tight Junction-Related Gene Expression in the Jejunal Mucosa
3.7. Protein Expression of Keap1/Nrf2 Signal Pathway in Jejunal Mucosa
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Xu, R.J.; Mellor, D.J.; Birtles, M.J.; Reynolds, G.W.; Simpson, H.V. Impact of intrauterine growth retardation on the gastrointestinal tract and the pancreas in newborn pigs. J. Pediatr. Gastroenterol. Nutr. 1994, 18, 231–240. [Google Scholar] [CrossRef]
- Dollberg, S.; Haklai, Z.; Mimouni, F.B.; Gorfein, L.; Gordon, E. Birth weight standards in the live-born population in Israel. Isr. Med. Assoc. J. 2005, 7, 311–314. [Google Scholar] [PubMed]
- Godfrey, K.; Cameron, I.; Hanson, M. Long-term consequences of foetal restriction. Curr. Obstet. Gynaecol. 2006, 16, 267–272. [Google Scholar] [CrossRef]
- Yucel, A.F.; Erboga, M.; Guzel, A.J. The role of curcumin on intestinal oxidative stress, cell proliferation and apoptosis after ischemia/reperfusion injury in rats. J. Mol. Histol. 2011, 42, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; He, J.T.; Sheng, M.M.; Zhang, H.; Niu, Y.; Zhang, L.L.; Wang, T. Effect of Curcumin Supplementation on Intestinal Antioxidant Function in Weaning Piglets with Intrauterine Growth Retardation. Food Sci. 2019, 40, 177–183. [Google Scholar]
- Zhang, H.; Zhao, F.; Peng, A.; Dong, L.; Wang, M.; Yu, L.H.; Loor, J.J.; Wang, H.G. Correction to effects of dietary l arginine and n carbamylglutamate supplementation on intestinal integrity, immune function, and oxidative status in intrauterine-growth-retarded suckling lambs. J. Agric. Food Chem. 2018, 66, 6254. [Google Scholar] [CrossRef]
- Su, W.P.; Zhang, H.; Ying, Z.X.; Li, Y.; Zhou, L.; Wang, F.; Zhang, L.L.; Wang, T. Effects of dietary L-methionine supplementation on intestinal integrity and oxidative status in intrauterine growth-retarded weanling piglets. Eur. J. Nutr. 2018, 57, 2735–2745. [Google Scholar] [CrossRef]
- Karowicz-Bilinska, A.; Dziora-Kornatowska, K.; Bartosz, G. Indices of oxidative stress in pregnancy with fetal growth restriction. Free Radic. Res. 2007, 41, 870–873. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Wang, T. Antioxidant capacity and concentration of redox-active trace mineral in fully weaned intra-uterine growth retardation piglets. J. Anim. Sci. Biotechnol. 2015, 6, 201–207. [Google Scholar] [CrossRef]
- Noorafshan, A.; Ashkani-Esfahani, S. A Review of Therapeutic Effects of Curcumin. Curr. Pharm. Des. 2013, 19, 2032–2046. [Google Scholar] [PubMed]
- Kiuchi, F.; Goto, Y.; Sugimoto, N.; Akao, N.; Kondo, K.; Tsuda, Y. Nematocidal activity of turmeric: Synergistic action of curcuminoids. Chem. Pharm. Bull. (Tokyo) 1993, 41, 1640–1643. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sung, B. Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol. Sci. 2009, 30, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From kitchen to clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef]
- Trujillo, J.; Chirino, Y.I.; Molina-Jijón, E.; Andérica-Romero, A.C.; Tapia, E.; Pedraza-Chaverrí, J. Renoprotective effect of the antioxidant curcumin: Recent findings. Redox Biol. 2013, 1, 448–456. [Google Scholar] [CrossRef]
- Zhang, J.; Hu, Z.; Lu, C.; Bai, K.; Zhang, L.; Wang, T. Effect of various levels of dietary curcumin on meat quality and antioxidant profile of breast muscle in broilers. J. Agric. Food Chem. 2015, 63, 3880–3886. [Google Scholar] [CrossRef]
- Venkatesan, N.; Chandrakasan, G. Modulation of cyclophosphamide-induced early lung injury by curcumin, an anti-inflammatory antioxidant. Mol. Cell. Biochem. 1995, 142, 79–87. [Google Scholar] [CrossRef]
- Lu, N.; Qiu, J.; Ying, Z.; Zheng, Y.; Wang, T.; Zhong, X. Effects of Varying Levels of Dietary Curcumin on Growth Performance, Digestibility and Serum Parameters of Weaned Piglets. J. Domest. Anim. Ecol. 2017, 38, 30–35. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sykiotis, G.P.; Bohmann, D. Keap1/Nrf2 signaling regulates oxidative stress tolerance and lifespan in Drosophila. Dev. Cell. 2008, 14, 76–85. [Google Scholar] [CrossRef]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends. Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Sykiotis, G.P.; Habeos, I.G.; Samuelson, A.V.; Dirk, B. The role of the antioxidant and longevity-promoting Nrf2 pathway in metabolic regulation. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Aroor, A.R.; Mandavia, C.; Ren, J.; Sowers, J.R.; Pulakat, L. Mitochondria and Oxidative Stress in the Cardiorenal Metabolic Syndrome. CardioRenal Med. 2012, 2, 87–109. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, C.; Duan, S.; Yuan, X.; Liang, J.; Hou, S. Curcumin attenuates potassium oxonate-induced hyperuricemia and kidney inflammation in mice. Biomed. Pharmacother. 2019, 118, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zadrożna, M.; Gawlik, M.; Nowak, B.; Marcinek, A.; Mrowiec, H.; Walas, S.; Wietecha-Posłuszny, R.; Zagrodzki, P. Antioxidants activities and concentration of selenium, zinc and copper in preterm and IUGR human placentas. J. Trace Elem. Med. Biol. 2009, 23, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Scandalios, J.G. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef]
- Wei, B.S.; Yuan, Y.W.; Fan, S.M.; Qing, H.Z.; Jian, Y.Z.; Li, P.X.; Xin, P.Y.; Dan, D.P.; Lei, S.; Bing, X. Curcumin protects intestinal mucosal barrier function of rat enteritis via activation of MKP-1 and attenuation of p38 and NF-κB activation. PLoS ONE 2010, 5, e12969. [Google Scholar]
- Del, R.D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. PLoS ONE 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Zhang, L.L.; Wang, Y.X.; Kong, Y.L.; Zhou, G.L.; Wang, C.; Zhong, X.; Wang, T. Effects of supplement of arginine on anti-oxidation and arginine metabolism in intrauterine growth retardation piglets. J. Nanjing Agric. Univ. 2017, 6, 1111–1118. [Google Scholar]
- Pierzchalski, P.; Krawiec, A.; Ptak-Belowska, A.; Baranska, A.; Konturek, S.J.; Pawlik, W.W. The mechanism of heat-shock protein 70 gene expression abolition in gastric epithelium caused by Helicobacter pylori infection. Helicobacter 2006, 11, 96–104. [Google Scholar] [CrossRef]
- Lallès, J.P.; Lacan, D.; David, J.C. A melon pulp concentrate rich in superoxide dismutase reduces stress proteins along the gastrointestinal tract of pigs. Nutrition 2011, 27, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Wu, W.Z.; Lin, G.; Li, D.F.; Wu, G.Y.; Wang, J.J. Temporal Proteomic Analysis Reveals Continuous Impairment of Intestinal Development in Neonatal Piglets with Intrauterine Growth Restriction. J. Proteome Res. 2010, 9, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T.; Fukata, M.; Arditi, M. TLR signaling in the gut in health and disease. J. Immunol. 2005, 174, 4453–4460. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Mochizuki, M.; Ishii, Y.; Ishii, T.; Shibata, T.; Kawamoto, Y.; Kelly, V.; Sekizawa, K.; Uchida, K.; Yamamoto, M. Transcription factor Nrf2 regulates inflammation by mediating the effect of 15-deoxy-Delta (12,14)-prostaglandin j (2). Mol. Cell. Biol. 2004, 24, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zong, X.; Ahmad, H.; Li, W.; Wang, Y.; Zhang, L.; Wang, T. Intrauterine Growth Restriction Impairs Small Intestinal Mucosal Immunity in Neonatal Piglets. J. Histochem. Cytochem. 2014, 62, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Hehlgans, T.; Pfeffer, K. The intriguing biology of the tumour necrosis factor/tumour necrosis factor receptor superfamily: Players, rules and the games. Immunology 2005, 115, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Yu, J.L.; Wei, X.D.; Zhang, X.P.; Li, J.C. Protective effects of curcumin on neonatal rats with necrotizing enterocolitis. Zhongguo Dang Dai Er Ke Za Zhi 2010, 12, 132–136. [Google Scholar]
- Xun, W.J.; Zhou, H.L.; Hou, G.Y.; Cao, T.; Shi, L.G.; Zhao, C.P. Effects of Curcumin on Ileum Mucosal Morphology, Gene Expression of Tight Junction Proteins and Inflammatory Cytokines, and Serum Immunoglobulin Levels of Early Weaning Piglets. Chin. J. Anim. Nutr. 2016, 28, 826–833. [Google Scholar]
- Naoki, M.; Keigyou, Y.; Ken, I.; Aki, H.; Akio, K.; Masayuki, Y.; Satoru, T. Nrf2 regulates the sensitivity of death receptor signals by affecting intracellular glutathione levels. Oncogene 2003, 22, 9275–9281. [Google Scholar]
- Zhu, L.H.; Xu, J.X.; Zhu, S.W.; Cai, X.; Yang, S.F.; Chen, X.L.; Gao, Q. Gene expression profiling analysis reveals weaning-induced cell cycle arrest and apoptosis in the small intestine of pigs. J. Anim. Sci. 2014, 92, 996–1006. [Google Scholar] [CrossRef]
- Samali, A.; Zhivotovsky, B.; Jones, D.; Nagata, S.; Orrenius, S. Apoptosis: Cell death defined by caspase activation. Cell Death Differ. 1999, 6, 495–496. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.R.; Deng, H.; Zhang, X.L.; Zen, Y.; Xiao, D.F.; Sun, W.Z.; Zhang, Z. Technology, Effects of orally administering the antimicrobial peptide buforin II on small intestinal mucosal membrane integrity, the expression of tight junction proteins and protective factors in weaned piglets challenged by enterotoxigenic Escherichia coli. Anim. Feed Sci. Technol. 2013, 186, 177–185. [Google Scholar]
- Furuse, M.; Sasaki, H.; Fujimoto, K.; Tsukita, S. A single gene product, claudin-1 or -2, reconstitutes tight junction strands and recruits occludin in fibroblasts. J. Cell Biol. 1998, 143, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Berkes, J.; Viswanathan, V.K.; Savkovic, S.D.; Hecht, G. Intestinal epithelial responses to enteric pathogens: Effects on the tight junction barrier, ion transport, and inflammation. Gut 2003, 52, 439–451. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No | Sequence (5′ to 3′) | Product Length (bp) |
|---|---|---|---|
| β-actin | XM_003124280.4 | F: CACGCCATCCTGCGTCTGGA R: AGCACCGTGTTGGCGTAGAG | 380 |
| hsp70 | X68213 | F: GCCCTGAATCCGCAGAATA R: TCCCCACGGTAGGAAACG | 281 |
| p53 | NM_213824.3 | F: CACTGGATGGCGAGTATTTCAC R: CTTAGACTTCAGGTGGCTGGA | 152 |
| IFNγ | AY188090.1 | F: TCAGCTTTGCGTGACTTTGTG R: GCTCTCTGGCCTTGGAACAT | 251 |
| ocln | NM_001163647.2 | F: ATGGCTGCCTTCTGCTTCAT R: TCACTTTCCCGTTGGACGAG | 75 |
| caspase3 | NM_214131.1 | F: ATAATTCAGGCCTGCCGAAG R: TGTTTCAGCGCTGCACAAAG | 200 |
| caspase9 | XM_003127618. | F: ACCCCTGACATGATCGAGGA R: CTGGCTTGAGTTCCACTGGT | 256 |
| bax | XM_003127290.4 | F: AGCATGCGGCCTCTATTTGA R: GGCCCGTGGACTTCACTTAT | 200 |
| IL-2 | NM_213861.1 | F: TGCACTAACCCTTGCACTCA R: GCAATGGCTCCAGTTGTTTCT | 83 |
| TNFα | NM_214022.1 | F: ATCGGCCCCCAGAAGGAAGAG R: GATGGCAGAGAGGAGGTTGAC | 351 |
| IL-6 | NM_214399.1 | F: AAATGTCGAGGCTGTGCAGA R: CTCAGGCTGAACTGCAGGAA | 207 |
| IL-1β | NM_214029.1 | F: TGCCAGCTATGAGCCACTTCC R: TGACGGGTCTCGAATGATGCT | 337 |
| Nrf2 | NM_001114671.1 | F: GACAAACCGCCTCAACTCAG R: GTCTCCACGTCGTAGCGTTC | 183 |
| Keap1 | XM_021076667.1 | F: CGTGGAGACAGAAACGTGGA R: CAATCTGCTTCCGACAGGGT | 239 |
| NQO1 | NM_001159613.1 | F: GATCATACTGGCCCACTCCG R: GAGCAGTCTCGGCAGGATAC | 200 |
| GCLC | XM_003482164.4 | F: GGCGACGAGGTGGAATACAT R: GTTTGGGTTTGTCCTTTCCCC | 123 |
| GCLM | XM_001926378.4 | F: GCATCTACAGCCTTACTGGGA R: GTTAAATCGGGCGGCATCAC | 180 |
| HO-1 | NM_001004027.1 | F: CAAGCAGAAAATCCTCGAAG R: GCTGAGTGTCAGGACCCATC | 241 |
| SOD1 | NM_001190422.1 | F: CATTCCATCATTGGCCGCAC R: TTACACCACAGGCCAAACGA | 118 |
| ZO-1 | XM005659811.1 | F-ACCCCCTACATGCTGACTCT R-TGGCTGCTTCAAGACATGGT | 167 |
| CAT | XM_021081498.1 | F: AGCTTTGCCCTTGCACAAAC R: ACATCCTGAACAAGAAGGGGC | 119 |
| Item | NBW | IUGR | IUGR + CUR |
|---|---|---|---|
| MDA (nmol/mgprot) | 1.57 ± 0.09 | 2.01 ± 0.15 * | 1.23 ± 0.13 # |
| GPx (U/mgprot) | 147.88 ± 24.69 | 123.20 ± 13.31 | 150.52 ± 31.86 |
| T-SOD (U/mgprot) | 16.54 ± 2.28 | 15.82 ± 3.04 | 28.65 ± 4.83 *,# |
| T-AOC (U/mgprot) | 1.58 ± 0.15 | 1.62 ± 0.08 | 1.92 ± 0.13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, E.; Zhang, J.; Han, H.; Wu, J.; Gan, Z.; Wei, C.; Zhang, L.; Wang, C.; Wang, T. Curcumin Alleviates IUGR Jejunum Damage by Increasing Antioxidant Capacity through Nrf2/Keap1 Pathway in Growing Pigs. Animals 2020, 10, 41. https://doi.org/10.3390/ani10010041
Yan E, Zhang J, Han H, Wu J, Gan Z, Wei C, Zhang L, Wang C, Wang T. Curcumin Alleviates IUGR Jejunum Damage by Increasing Antioxidant Capacity through Nrf2/Keap1 Pathway in Growing Pigs. Animals. 2020; 10(1):41. https://doi.org/10.3390/ani10010041
Chicago/Turabian StyleYan, Enfa, Jiaqi Zhang, Hongli Han, Jiamin Wu, Zhending Gan, Chengheng Wei, Lili Zhang, Chao Wang, and Tian Wang. 2020. "Curcumin Alleviates IUGR Jejunum Damage by Increasing Antioxidant Capacity through Nrf2/Keap1 Pathway in Growing Pigs" Animals 10, no. 1: 41. https://doi.org/10.3390/ani10010041
APA StyleYan, E., Zhang, J., Han, H., Wu, J., Gan, Z., Wei, C., Zhang, L., Wang, C., & Wang, T. (2020). Curcumin Alleviates IUGR Jejunum Damage by Increasing Antioxidant Capacity through Nrf2/Keap1 Pathway in Growing Pigs. Animals, 10(1), 41. https://doi.org/10.3390/ani10010041