Could Dietary Black Soldier Fly Meal Inclusion Affect the Liver and Intestinal Histological Traits and the Oxidative Stress Biomarkers of Siberian Sturgeon (Acipenser baerii) Juveniles?


 , , ,
, , ,  ,
,  ,
,  ,
,  ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets and Fish Management
2.2. Sampling and Histological Investigations
2.3. Biochemical Analyses
2.4. Statistical Analyses
3. Results
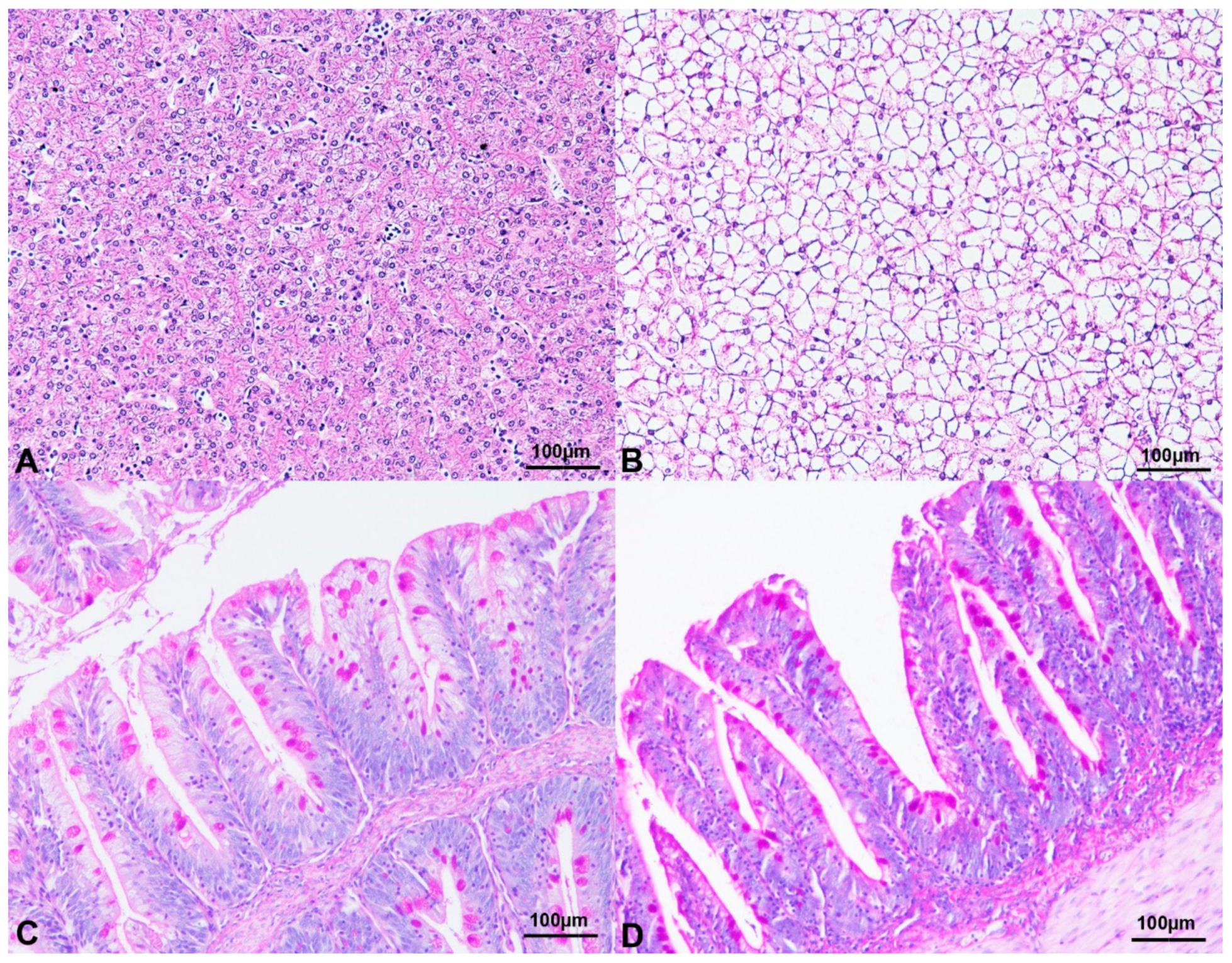
3.1. Histological Investigations
3.2. Oxidative Stress Biomarkers
4. Discussion
4.1. Histological Investigations
4.2. Oxidative Stress Biomarkers
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alltech 2019 Global Feed Survey. Available online: https://www.alltech.com/feed-survey (accessed on 28 August 2019).
- Glencross, B.D.; Baily, J.; Berntssen, M.H.; Hardy, R.; MacKenzie, S.; Tocher, D.R. Risk assessment of the use of alternative animal and plant raw material resources in aquaculture feeds. Rev. Aquac. 2019, 1–56. [Google Scholar] [CrossRef]
- Sánchez-Lozano, N.B.; Martínez-Llorens, S.; Tomás-Vidal, A.; Cerdá, M.J. Effect of high-level fish meal replacement by pea and rice concentrate protein on growth, nutrient utilization and fillet quality in gilthead seabream (Sparus aurata, L.). Aquaculture 2009, 298, 83–89. [Google Scholar] [CrossRef]
- Kokou, F.; Sarropoulou, E.; Cotou, E.; Rigos, G.; Henry, M.; Alexis, M.; Kentouri, M. Effects of fish meal replacement by a soybean protein on growth, histology, selected immune and oxidative status markers of gilthead sea bream, Sparus aurata. J. World Aquac. Soc. 2015, 46, 115–128. [Google Scholar] [CrossRef]
- Gai, F.; Gasco, L.; Daprà, F.; Palmegiano, G.B.; Sicuro, B. Enzymatic and histological evaluation of gut and liver in rainbow trout, Oncorhynchus mykiss, fed with rice protein concentrate based diets. J. World Aquac. Soc. 2012, 43, 218–229. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, Å. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Merrifield, D.L.; Olsen, R.E.; Myklebust, R.; Ringo, E.; El-Shemy, H. Dietary effect of soybean (Glycine max) products on gut histology and microbiota of fish. In Soybean and Nutrition; El-Shemy, H., Ed.; In Tech: Rijeka, Croatia, 2011; pp. 231–250. [Google Scholar]
- Zhou, J.S.; Liu, S.S.; Ji, H.; Yu, H.B. Effect of replacing dietary fish meal with black soldier fly larvae meal on growth and fatty acid composition of Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2016, 24, 424–433. [Google Scholar] [CrossRef]
- Voorhees, J.M.; Barnes, M.E.; Chipps, S.R.; Brown, M.L. Bioprocessed soybean meal replacement of fish meal in rainbow trout (Oncorhynchus mykiss) diets. Cogent Food Agric. 2019, 5, 1579482. [Google Scholar] [CrossRef]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The potential role of insects as feed: A multi-perspective review. Animals 2019, 9, 199. [Google Scholar] [CrossRef]
- Moutinho, S.; Martínez-Llorens, S.; Tomás-Vidal, A.; Jover-Cerdá, M.; Oliva-Teles, A.; Peres, H. Meat and bone meal as partial replacement for fish meal in diets for gilthead seabream (Sparus aurata) juveniles: Growth, feed efficiency, amino acid utilization, and economic efficiency. Aquaculture 2017, 468, 271–277. [Google Scholar] [CrossRef]
- EU Regulation 893/2017 of 24 May 2017 amending Annexes I and IV to Regulation (EC) No 999/2001 of the European Parliament and of the Council and Annexes X, XIV and XV to Commission Regulation (EU) No 142/2011 as regards the provisions on processed animal protein.
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef] [PubMed]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black Soldier Fly biowaste treatment—Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals Fed Insect-Based Diets: State-of-the-Art on Digestibility, Performance and Product Quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Capucchio, M.T.; Caldaroni, B.; Magara, G.; Dörr, A.J.M.; Biasato, I.; Biasibetti, E.; Righetti, M.; Pastorino, P.; Prearo, M.; et al. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 50–57. [Google Scholar] [CrossRef]
- Rimoldi, S.; Gini, E.; Iannini, F.; Gasco, L.; Terova, G. The effects of dietary insect meal from Hermetia illucens prepupae on autochthonous gut microbiota of rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 143. [Google Scholar] [CrossRef]
- Józefiak, A.; Nogales-Mérida, S.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Effects of insect diets on the gastrointestinal tract health and growth performance of Siberian sturgeon (Acipenser baerii Brandt, 1869). BMC Vet. Res. 2019, 15, 348. [Google Scholar] [CrossRef]
- Caimi, C.; Renna, M.; Lussiana, C.; Bonaldo, A.; Gariglio, M.; Meneguz, M.; Dabbou, S.; Schiavone, A.; Gai, F.; Elia, A.C.; et al. First insights on Black Soldier Fly (Hermetia illucens L.) larvae meal dietary administration in Siberian sturgeon (Acipenser baerii Brandt) juveniles. Aquaculture 2020, 734539. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile Jian carp (Cyprinus carpio var. Jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]
- Xiao, X.; Jin, P.; Zheng, L.; Cai, M.; Yu, Z.; Yu, J.; Zhang, J. Effects of black soldier fly (Hermetia illucens) larvae meal protein as a fishmeal replacement on the growth and immune index of yellow catfish (Pelteobagrus fulvidraco). Aquac. Res. 2018, 49, 1569–1577. [Google Scholar] [CrossRef]
- Webster, C.D.; Rawles, S.D.; Koch, J.F.; Thompson, K.R.; Kobayashi, Y.; Gannam, A.L.; Twibell, R.G.; Hyde, N.M. Bio-Ag reutilization of distiller’s dried grains with solubles (DDGS) as a substrate for black soldier fly larvae, Hermetia illucens, along with poultry by-product meal and soybean meal, as total replacement of fish meal in diets for Nile tilapia, Oreochromis niloticus. Aquac. Nutr. 2016, 22, 976–988. [Google Scholar] [CrossRef]
- Devic, E.; Leschen, W.; Murray, F.; Little, D.C. Growth performance, feed utilization and body composition of advanced nursing Nile tilapia (Oreochromis niloticus) fed diets containing Black Soldier Fly (Hermetia illucens) larvae meal. Aquac. Nutr. 2018, 24, 416–423. [Google Scholar] [CrossRef]
- Rapatsa, M.M.; Moyo, N.A. Enzyme activity and histological analysis of Clarias gariepinus fed on Imbrasia belina meal used for partial replacement of fishmeal. Fish Physiol. Biochem. 2019, 45, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Sankian, Z.; Khsravi, S.; Kim, Y.O.; Lee, S.M. Effects of dietary inclusion of yellow mealworm (Tenebrio molitor) meal on growth performance, feed utilization, body composition, plasma biochemical indices, selected immune parameters and antioxidant enzyme activities of mandarin fish (Siniperca scherzeri) juvenile. Aquaculture 2018, 496, 79–87. [Google Scholar] [CrossRef]
- Magalhães, R.; Sánchez-López, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Chatzifotis, S.; Piccolo, G. Does dietary insect meal affect the fish immune system? The case of mealworm, Tenebrio molitor on European sea bass, Dicentrarchus labrax. Dev. Comp. Immunol. 2018, 81, 204–209. [Google Scholar] [CrossRef]
- Piccolo, G.; Iaconisi, V.; Marono, S.; Gasco, L.; Loponte, R.; Nizza, S.; Bovera, F.; Parisi, G. Effect of Tenebrio molitor larvae meal on growth performance, in vivo nutrients digestibility, somatic and marketable indexes of gilthead sea bream (Sparus aurata). Anim. Feed Sci. Technol. 2017, 226, 12–20. [Google Scholar] [CrossRef]
- Wang, G.; Peng, K.; Hu, J.; Yi, C.; Chen, X.; Wu, H.; Huang, Y. Evaluation of defatted black soldier fly (Hermetia illucens L.) larvae meal as an alternative protein ingredient for juvenile Japanese seabass (Lateolabrax japonicus) diets. Aquaculture 2019, 507, 144–154. [Google Scholar] [CrossRef]
- Lock, E.J.; Arsiwalla, T.; Waagbø, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) post smolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, A.; Lock, E.J. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Biancarosa, I.; Sele, V.; Belghit, I.; Ørnsrud, R.; Lock, E.J.; Amlund, H. Replacing fish meal with insect meal in the diet of Atlantic salmon (Salmo salar) does not impact the amount of contaminants in the feed and it lowers accumulation of arsenic in the fillet. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2019. [Google Scholar] [CrossRef]
- Guerreiro, I.; Castro, C.; Antunes, B.; Coutinho, F.; Rangel, F.; Couto, A.; Serra, C.R.; Peres, H.; Pousão-Ferreira, P.; Matos, E.; et al. Catching black soldier fly for meagre: Growth, whole-body fatty acid profile and metabolic responses. Aquaculture 2020, 516, 734613. [Google Scholar] [CrossRef]
- Rašković, B.S.; Stanković, M.B.; Marković, Z.Z.; Poleksić, V.D. Histological methods in the assessment of different feed effects on liver and intestine of fish. J. Agric. Sci. 2011, 56, 87–100. [Google Scholar] [CrossRef]
- Baeza-Ariño, R.; Martínez-Llorens, S.; Nogales-Mérida, S.; Jover-Cerda, M.; Tomás-Vidal, A. Study of liver and gut alterations in sea bream, Sparus aurata L., fed a mixture of vegetable protein concentrates. Aquac. Res. 2014, 47, 460–471. [Google Scholar] [CrossRef]
- Petrinec, Z.; Nejedli, S.; Kuzir, S.; Opacak, A. Mucosubstances of the digestive tract mucosa in northern pike (Esox lucius L.) and European catfish (Silurus glanis L.). Vet. Arh. 2005, 75, 317. [Google Scholar]
- Murray, H.M.; Wright, G.M.; Goff, G.P. A comparative histological and histochemical study of the post-gastric alimentary canal from three species of pleuronectid, the Atlantic halibut, the yellowtail flounder and the winter flounder. J. Fish Biol. 1996, 48, 187–206. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant defenses in fish: Biotic and abiotic factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Olsvik, P.A.; Torstensen, B.E.; Hemre, G.I.; Sanden, M.; Waagbø, R. Hepatic oxidative stress in Atlantic salmon (Salmo salar L.) transferred from a diet based on marine feed ingredients to a diet based on plant ingredients. Aquac. Nutr. 2011, 17, 424–436. [Google Scholar] [CrossRef]
- Morrow, M.D.; Higgs, D.; Kennedy, C.J. The effects of diet composition and ration on biotransformation enzymes and stress parameters in rainbow trout, Oncorhynchus mykiss. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 137, 143–154. [Google Scholar] [CrossRef]
- Rueda-Jasso, R.; Conceiçao, L.E.; Dias, J.; De Coen, W.; Gomes, E.; Rees, J.F.; Sorgeloos, P. Effect of dietary non-protein energy levels on condition and oxidative status of Senegalese sole (Solea senegalensis) juveniles. Aquaculture 2004, 231, 417–433. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Peña-Llopis, S.; Gómez-Requeni, P.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J. Effect of fish meal replacement by plant protein sources on non-specific defence mechanisms and oxidative stress in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 387–400. [Google Scholar] [CrossRef]
- Fontagné, S.; Bazin, D.; Brèque, J.; Vachot, C.; Bernarde, C.; Rouault, T.; Bergot, P. Effects of dietary oxidized lipid and vitamin A on the early development and antioxidant status of Siberian sturgeon (Acipenser baeri) larvae. Aquaculture 2006, 257, 400–411. [Google Scholar] [CrossRef]
- Ogunji, J.O.; Nimptsch, J.; Wiegand, C.; Schulz, C.; Rennert, B. Effect of housefly maggot meal (magmeal) diets on catalase, and glutathione S-transferase in the liver and gills of carp Cyprinus carpio fingerling. Int. Aquat. Res. 2011, 3, 11–20. [Google Scholar] [CrossRef]
- Alegbeleye, W.O.; Obasa, S.O.; Olude, O.O.; Otubu, K.; Jimoh, W. Preliminary evaluation of the nutritive value of the variegated grasshopper (Zonocerus variegatus L.) for African catfish Clarias gariepinus (Burchell. 1822) fingerlings. Aquac. Res. 2012, 43, 412–420. [Google Scholar] [CrossRef]
- Taufek, N.M.; Aspani, F.; Muin, H.; Raji, A.A.; Razak, S.A.; Alias, Z. The effect of dietary cricket meal (Gryllus bimaculatus) on growth performance, antioxidant enzyme activities, and haematological response of African catfish (Clarias gariepinus). Fish Physiol. Biochem. 2016, 42, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Directive 2010/63/EU of the European Parliament and of the council of 22 September 2010 on the protection of animals used for scientific purposes.
- Finke, M.D. Estimate of chitin in raw whole insects. Zoo Biol. 2007, 26, 105–115. [Google Scholar] [CrossRef]
- Cao, X.J.; Wang, W.M. Histology and mucin histochemistry of the digestive tract of yellow catfish, Pelteobagrus fulvidraco. Anat. Histol. Embryol. 2009, 38, 254–261. [Google Scholar] [CrossRef]
- Pryor, G.S.; Royes, J.B.; Chapman, F.A.; Miles, R.D. Mannanoligosaccharides in fish nutrition: Effects of dietary supplementation on growth and gastrointestinal villi structure in Gulf of Mexico sturgeon. N. Am. J. Aquac. 2003, 65, 106–111. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Zarantoniello, M.; Bruni, L.; Randazzo, B.; Vargas, A.; Gioacchini, G.; Truzzi, C.; Annibaldi, A.; Riolo, P.; Parisi, G.; Cardinaletti, G.; et al. Partial dietary inclusion of Hermetia illucens (Black Soldier Fly) full-fat prepupae in zebrafish feed: Biometric, histological, biochemical, and molecular implications. Zebrafish 2018, 15, 519–532. [Google Scholar] [CrossRef]
- Olsson, C. Gut anatomy and morphology: Gut anatomy. In Enciclopedia of Fish Phisiology—From Genome to Environment; Farrel, A.P., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 1270–1275. [Google Scholar]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Urán, P.A.; Schrama, J.W.; Rombout, J.H.W.M.; Obach, A.; Jensen, L.; Koppe, W.; Verreth, J.A.J. Soybean meal-induced enteritis in Atlantic salmon (Salmo salar L.) at different temperatures. Aquac. Nutr. 2008, 14, 324–330. [Google Scholar] [CrossRef]
- Glencross, B.D.; Evans, D.; Dods, K.; McCafferty, P.; Hawkins, W.; Maas, R.; Sipsas, S. Evaluation of the digestible value of lupin and soybean protein concentrates and isolates when fed to rainbow trout, Oncorhynchus mykiss, using either stripping or settlement faecal collection methods. Aquaculture 2005, 245, 211–220. [Google Scholar] [CrossRef]
- Kokou, F.; Sarropoulou, E.; Cotou, E.; Kentouri, M.; Alexis, M.; Rigos, G. Effects of graded dietary levels of soy protein concentrate supplemented with methionine and phosphate on the immune and antioxidant responses of gilthead sea bream (Sparus aurata L.). Fish Shellfish Immunol. 2017, 64, 111–121. [Google Scholar] [CrossRef]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R.M. Soybean meal-induced enteritis in Altantic salmon (Salmo salar) and Chinook salmon (Oncorhynchus tshawytscha) but not in pink salmon (O. gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- Kittel, E.C.; Sealey, W.M.; Small, B.C. Preliminary investigation of dietary soy sensitivity in Shovelnose Sturgeon. J. Appl. Aquac. 2014, 26, 356–369. [Google Scholar] [CrossRef]
- Elia, A.C.; Dörr, A.J.M.; Galarini, R. Comparison of organochlorine pesticides, PCBs, and heavy metal contamination and of detoxifying response in tissues of Ameiurus melas from Corbara, Alviano, and Trasimeno lakes, Italy. Bull. Environ. Contam. Toxicol. 2007, 78, 463–468. [Google Scholar] [CrossRef]
- Elia, A.C.; Dörr, A.J.M.; Abete, M.C.; Prearo, M. Seasonal variability of detoxificant response and heavy metal accumulation in tissues of both sexes in Tinca tinca (L.) from Lake Trasimeno. Rev. Fish Biol. Fish. 2010, 20, 425–434. [Google Scholar] [CrossRef]
- Pacini, N.; Prearo, M.; Abete, M.C.; Brizio, P.; Dörr, A.J.M.; Reimschuessel, R.; Andersen, W.; Gasco, L.; Righetti, M.; Elia, A.C. Antioxidant responses and renal crystal formation in rainbow trout treated with melamine administered individually or in combination with cyanuric acid. J. Toxicol. Environ. Health A 2013, 76, 491–508. [Google Scholar] [CrossRef]
- Elia, A.C.; Ciccotelli, V.; Pacini, N.; Dörr, A.J.M.; Gili, M.; Natali, M.; Gasco, L.; Prearo, M.; Abete, M.C. Transferability of oxytetracycline (OTC) from feed to carp muscle and evaluation of the antibiotic effects on antioxidant systems in liver and kidney. Fish Physiol. Biochem. 2014, 40, 1055–1068. [Google Scholar] [CrossRef]
- Furné, M.; García-Gallego, M.; Hidalgo, M.C.; Sanz, A. Effect of dietary macronutrient proportion on intermediate metabolism and oxidative status in sturgeon (Acipenser naccarii) and trout (Oncorhynchus mykiss): Comparative study. Fish Physiol. Biochem. 2016, 42, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Babaei, S.; Abedian, A.; Email, K.; Hedayati, M.; Ali Sadati, M.A.Y.; Metón, I. Effect of diet composition on growth performance, hepatic metabolism and antioxidant activities in Siberian sturgeon (Acipenser baerii, Brandt, 1869) submitted to starvation and refeeding. Fish Physiol. Biochem. 2016, 42, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- Khoushab, F.; Yamabhai, M. Chitin research revisited. Mar. Drugs 2010, 8, 1988–2012. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.H.; Kim, S.K. Antioxidant effects of chitin, chitosan and their derivatives. Adv. Food Nutr. Res. 2014, 73, 15–31. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, S.; Chen, J.; Qin, C.; Lin, H.; Li, W. Stimulatory effects of chitinase on growth and immune defense of orange-spotted grouper (Epinephelus coioides). Fish Shellfish Immunol. 2012, 32, 844–854. [Google Scholar] [CrossRef]
- Pacini, N.; Abete, M.C.; Dörr, A.J.M.; Prearo, M.; Natali, M.; Elia, A.C. Detoxifying response in juvenile tench fed by selenium diet. Environ. Toxicol. Pharmacol. 2012, 33, 46–52. [Google Scholar] [CrossRef]
- Pacini, N.; Elia, A.C.; Abete, M.C.; Dörr, A.J.M.; Brizio, P.; Gasco, L.; Righetti, M.; Prearo, M. Antioxidant response versus selenium accumulation in the liver and kidney of the Siberian sturgeon (Acipenser baeri). Chemosphere 2013, 93, 2405–2412. [Google Scholar] [CrossRef]
- Elia, A.C.; Abete, M.C.; Pacini, N.; Dörr, A.J.M.; Scanzio, T.; Prearo, M. Antioxidant biomarker survey ensuing long-term selenium withdrawal in Acipenser baeri fed Se-cysteine diets. Environ. Toxicol. Pharmacol. 2014, 37, 1131–1139. [Google Scholar] [CrossRef]
- Hao, Y.; Zhan, Z.F.; Guo, P.F. Soybean β-conglycinin-induced gut hypersensitivity reaction in a piglet model. Arch. Anim. Nutr. 2009, 63, 188–202. [Google Scholar] [CrossRef]
- Zhang, J.X.; Guo, L.G.; Feng, L.; Jiang, W.D.; Kuang, S.Y.; Liu, Y.; Hu, K.; Jiang, J.; Li, S.H.; Tang, L.; et al. Soybean β-conglycinin induces inflammation and oxidation and causes dysfunction of intestinal digestion and absorption in fish. PLoS ONE 2013, 8, e58115. [Google Scholar] [CrossRef]
- Peng, S.; Chen, L.; Qin, J.G.; Hou, J.; Yu, N.; Long, Z.; Ye, J.; Sun, X. Effects of replacement of dietary fish oil by soybean oil on growth performance and liver biochemical composition in juvenile black seabream, Acanthopagrus schlegeli. Aquaculture 2008, 276, 154–161. [Google Scholar] [CrossRef]
- Ji, H.; Zhang, J.L.; Huang, J.Q.; Cheng, X.F.; Liu, C. Effect of replacement of dietary fish meal with silkworm pupae meal on growth performance, body composition, intestinal protease activity and health status in juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Res. 2015, 46, 1209–1221. [Google Scholar] [CrossRef]


{kind=link}
| Ingredients (%, as Fed) | Experimental Diets | ||||
|---|---|---|---|---|---|
| HIM | HIM0 | HIM25 | HIM50 | VEG | |
| Fish meal (Chile, super prime) a | 70 | 52.5 | 35 | 32 | |
| HI larva meal b | 0 | 18.5 | 37.5 | 0 | |
| Wheat meal | 14 | 12 | 10 | 0 | |
| Corn gluten meal | 0 | 0 | 0 | 15 | |
| Soybean protein concentrate | 0 | 0 | 0 | 20 | |
| Soybean meal | 0 | 0 | 0 | 14 | |
| Starch gelatinized, D500 | 8 | 8 | 8 | 8 | |
| Fish oil | 6 | 7 | 7.5 | 9 | |
| Vitamin mixture c | 1 | 1 | 1 | 1 | |
| Mineral mixture d | 1 | 1 | 1 | 1 | |
| Chemical Composition (% as Fed) e | |||||
| DM | 94.94 | 96.41 | 96.39 | 96.29 | 97.37 |
| CP | 62.51 | 50.29 | 50.65 | 50.2 | 50.87 |
| EE | 4.03 | 12.68 | 12.62 | 12.1 | 12.81 |
| Ash | 8.2 | 13.15 | 11.71 | 10.24 | 9.91 |
| Chitin f | 4.97 | nd | 0.72 | 1.92 | nd |
| NFE g | 20.29 | 23.88 | 24.15 | 25.55 | 26.41 |
| GE (MJ kg−1) | 20.76 | 19.77 | 19.65 | 20.64 | 20.44 |
| Alteration | Score 0 | Score 1 | Score 2 | Score 3 |
|---|---|---|---|---|
| Liver | ||||
| Vacuolization | Absent | Mild, focal to multifocal vacuolization of the hepatocytes (<25% of the cells is affected) | Moderate, multifocal to diffuse vacuolization of the hepatocytes (>25%, but <50% of the cells is affected) | Severe, multifocal to diffuse vacuolization of the hepatocytes (>50% of the cells is affected) |
| Nuclear displacement | Absent | The nucleus of the hepatocytes is mildly displaced towards the cell membrane | The nucleus of the hepatocytes is moderately displaced towards the cell membrane | The nucleus of the hepatocytes is severely displaced towards (and adhered to) the cell membrane |
| Lymphocytic infiltrates | Absent | Mild, focal to multifocal interstitial and/or perivascular lymphocytic infiltrates (<25% of the liver parenchyma is affected) | Moderate, multifocal to diffuse interstitial and/or perivascular lymphocytic infiltrates (>25%, but <50% of the liver parenchyma is affected) | Severe, multifocal to diffuse interstitial and/or perivascular lymphocytic infiltrates (>50% of the liver parenchyma is affected) |
| Necrotic tissue area | Absent | Small, focal to multifocal areas of hepatocyte necrosis | Small to large, multifocal areas of hepatocyte necrosis | Large, multifocal to diffuse areas of hepatocyte necrosis |
| Spiral valve | ||||
| Lymphocytic infiltrates | Absent | Mild, focal to multifocal mucosal, submucosal and/or muscular lymphocytic infiltrates (<25% of the intestinal wall is affected) | Moderate, multifocal to diffuse mucosal, submucosal and/or muscular lymphocytic infiltrates (>25%, but <50% of the intestinal wall is affected) | Severe, multifocal to diffuse mucosal, submucosal and/or muscular lymphocytic infiltrates (>50% of the intestinal wall is affected) |
| Items | Experimental Diets | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM50 | VEG | |||
| Liver | ||||||
| Vacuolization | 2.67 | 2.58 | 2.83 | 2.75 | 0.073 | 0.663 |
| Nuclear displacement | 1.50 | 1.83 | 1.92 | 2.08 | 0.100 | 0.215 |
| Lymphocytic infiltrates | 1.42 | 1.33 | 1.33 | 1.67 | 0.098 | 0.601 |
| Necrotic tissue area | Absent | Absent | Absent | Absent | ||
| Spiral valve | ||||||
| Lymphocytic infiltrates | Absent | Absent | Absent | Absent | ||
| Items | Experimental Diets | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM50 | VEG | |||
| Vh (µm) | 330.93 | 313.16 | 338.04 | 314.54 | 4.29 | 0.105 |
| GC number | 61.28 | 77.08 | 68.76 | 67.44 | 2.909 | 0.293 |
| Items | Experimental Diets | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| HIM0 | HIM25 | HIM50 | VEG | |||
| Liver | ||||||
| MDA | 150.73 | 140.81 | 159.64 | 129.13 | 4.53 | 0.094 |
| SOD | 23.53 b | 20.18 b | 33.22 a | 26.84 a,b | 1.25 | 0.001 |
| CAT | 614.13 b | 644.23 b | 734.23 a,b | 836.06 a | 20.51 | 0.001 |
| GPx | 69.75 a | 55.71 a,b | 50.53 b | 57.16 a,b | 2.48 | 0.037 |
| GR | 30.07 b | 30.42 b | 46.63 a | 34.35 b | 1.59 | 0.001 |
| EROD | 13.22 | 17.96 | 10.6 | 15.01 | 1.06 | 0.077 |
| GST | 246.50 a | 239.30 a | 274.39 a | 155.80 b | 11.16 | 0.001 |
| Kidney | ||||||
| MDA | 108.49 | 111.55 | 108.75 | 121.64 | 5.47 | 0.816 |
| SOD | 12.61 b | 18.51 a,b | 22.95 a | 18.82 a,b | 1.02 | 0.001 |
| CAT | 85.31 b | 81.10 b | 109.18 a,b | 134.15 a | 5.59 | 0.003 |
| GPx | 53.55 b,c | 67.78 b | 36.24 c | 83.64 a | 4.18 | 0.001 |
| GR | 16.64 b | 26.63 a | 25.94 a | 29.52 a | 1.36 | 0.003 |
| EROD | 6.34 c | 38.07 a | 22.42 b | 12.32 b,c | 3.49 | 0.001 |
| GST | 48.05 b | 62.55 b | 95.22 a | 52.60 b | 4.86 | 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caimi, C.; Gasco, L.; Biasato, I.; Malfatto, V.; Varello, K.; Prearo, M.; Pastorino, P.; Bona, M.C.; Francese, D.R.; Schiavone, A.; et al. Could Dietary Black Soldier Fly Meal Inclusion Affect the Liver and Intestinal Histological Traits and the Oxidative Stress Biomarkers of Siberian Sturgeon (Acipenser baerii) Juveniles? Animals 2020, 10, 155. https://doi.org/10.3390/ani10010155
Caimi C, Gasco L, Biasato I, Malfatto V, Varello K, Prearo M, Pastorino P, Bona MC, Francese DR, Schiavone A, et al. Could Dietary Black Soldier Fly Meal Inclusion Affect the Liver and Intestinal Histological Traits and the Oxidative Stress Biomarkers of Siberian Sturgeon (Acipenser baerii) Juveniles? Animals. 2020; 10(1):155. https://doi.org/10.3390/ani10010155
Chicago/Turabian StyleCaimi, Christian, Laura Gasco, Ilaria Biasato, Vanda Malfatto, Katia Varello, Marino Prearo, Paolo Pastorino, Maria Cristina Bona, Danila Raffaella Francese, Achille Schiavone, and et al. 2020. "Could Dietary Black Soldier Fly Meal Inclusion Affect the Liver and Intestinal Histological Traits and the Oxidative Stress Biomarkers of Siberian Sturgeon (Acipenser baerii) Juveniles?" Animals 10, no. 1: 155. https://doi.org/10.3390/ani10010155
APA StyleCaimi, C., Gasco, L., Biasato, I., Malfatto, V., Varello, K., Prearo, M., Pastorino, P., Bona, M. C., Francese, D. R., Schiavone, A., Elia, A. C., Dörr, A. J. M., & Gai, F. (2020). Could Dietary Black Soldier Fly Meal Inclusion Affect the Liver and Intestinal Histological Traits and the Oxidative Stress Biomarkers of Siberian Sturgeon (Acipenser baerii) Juveniles? Animals, 10(1), 155. https://doi.org/10.3390/ani10010155