
Does Organic Farming Increase Raspberry Quality, Aroma and Beneficial Bacterial Biodiversity?

 , ,
, ,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation Method
2.2. DNA Extraction and Next-Generation Sequencing for Microbiome Analysis
2.3. Bioinformatic Analysis
2.4. Cultural Dependent Characterization of Fruit Microbiome
2.5. Assessment of Fruit Quality Parameters
2.6. Fruit Volatile Analysis
2.7. Bacterial Volatile Analysis by Proton-Transfer Reaction−Mass Spectrometry (PTR–MS) and in Silico Fruit Volatilome Assembly
2.8. Statistical Analysis
3. Results
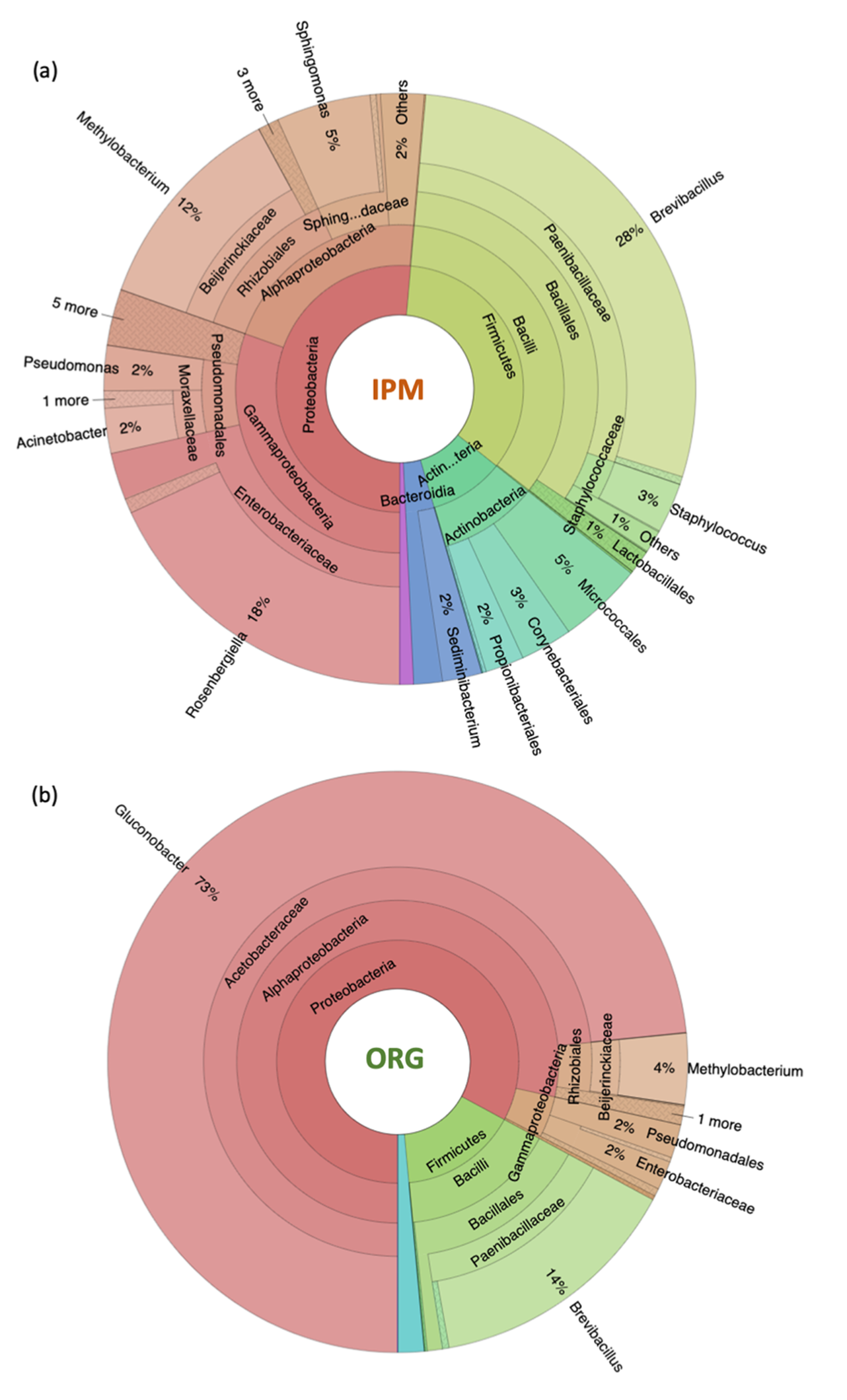
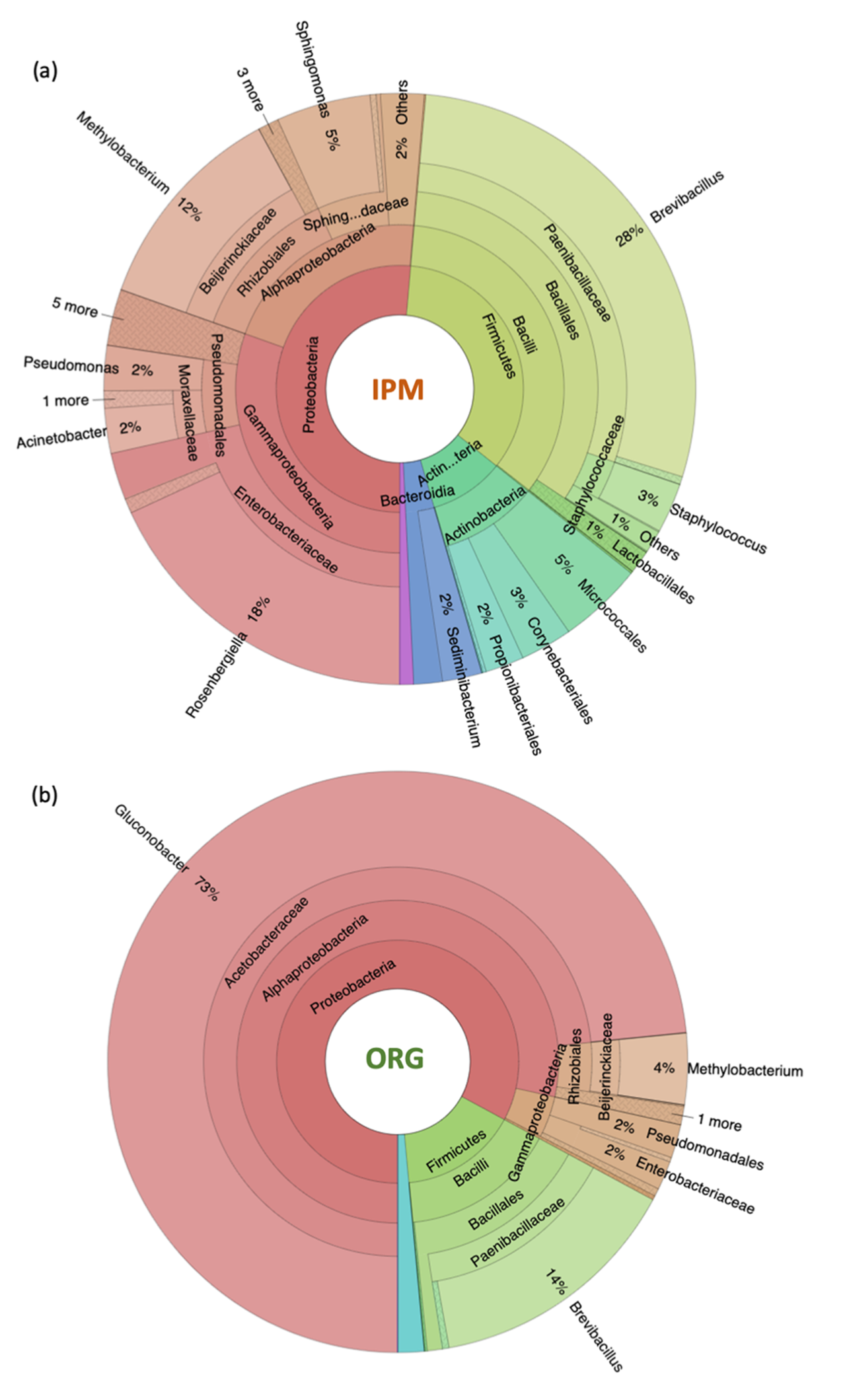
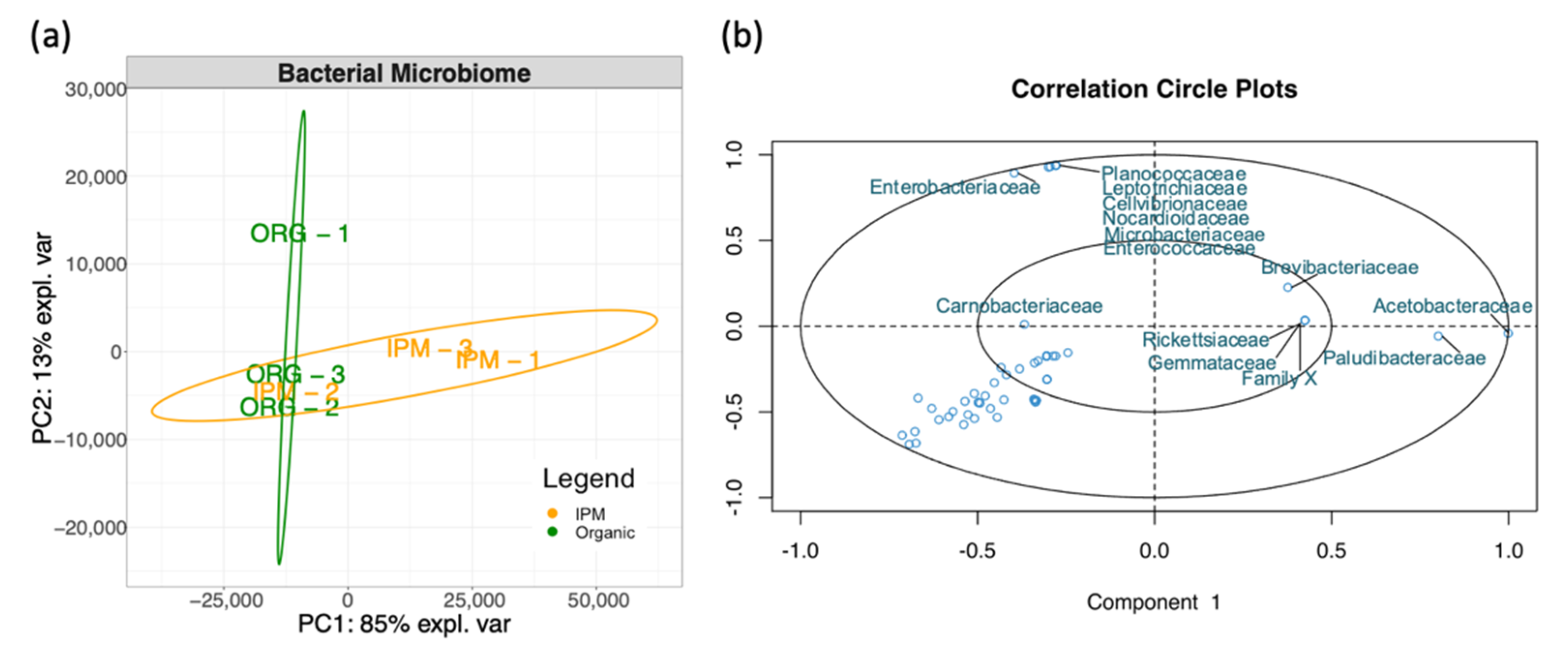
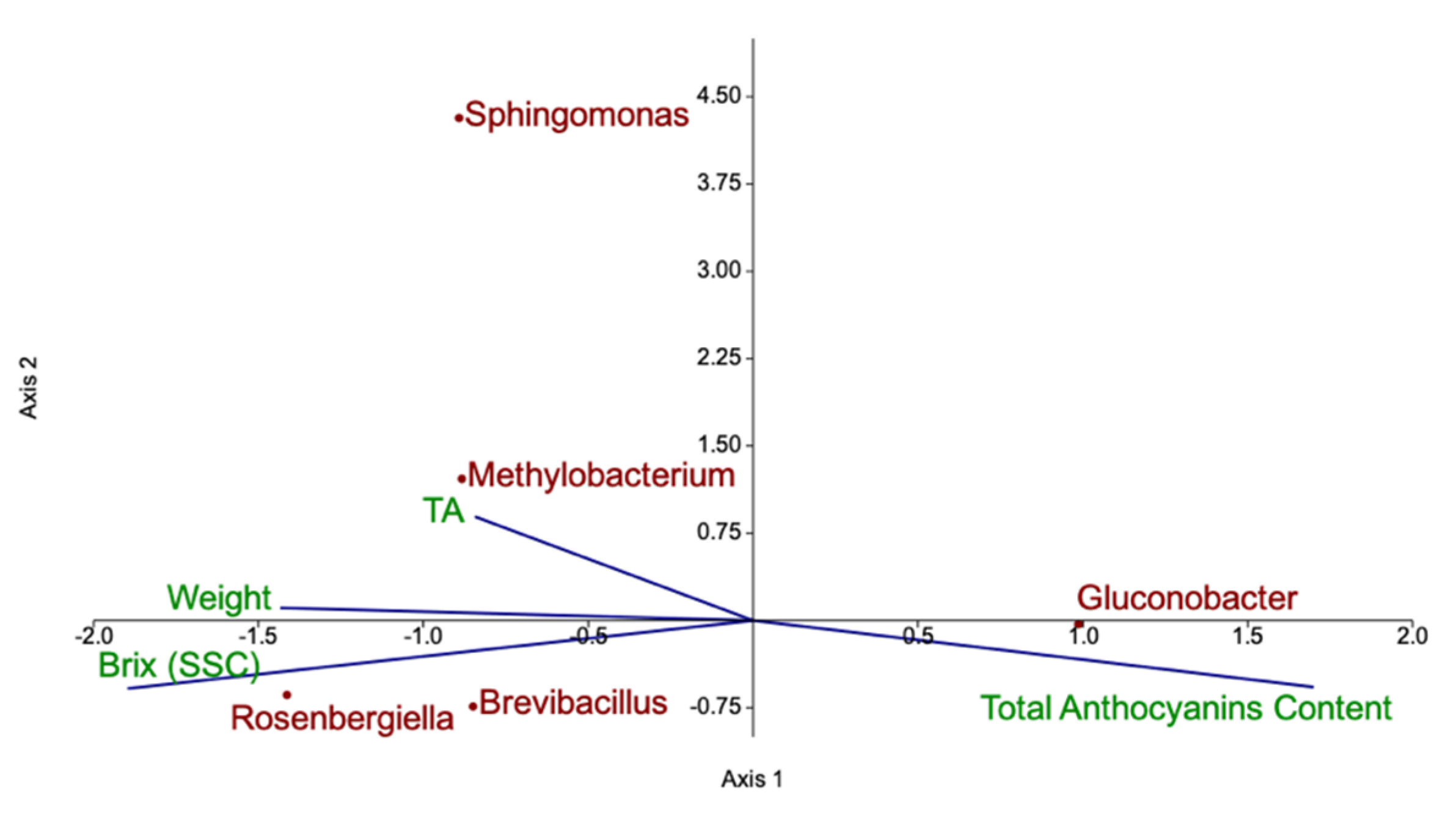
3.1. Characterization of Raspberry Culture Dependent and Independent Bacteriome
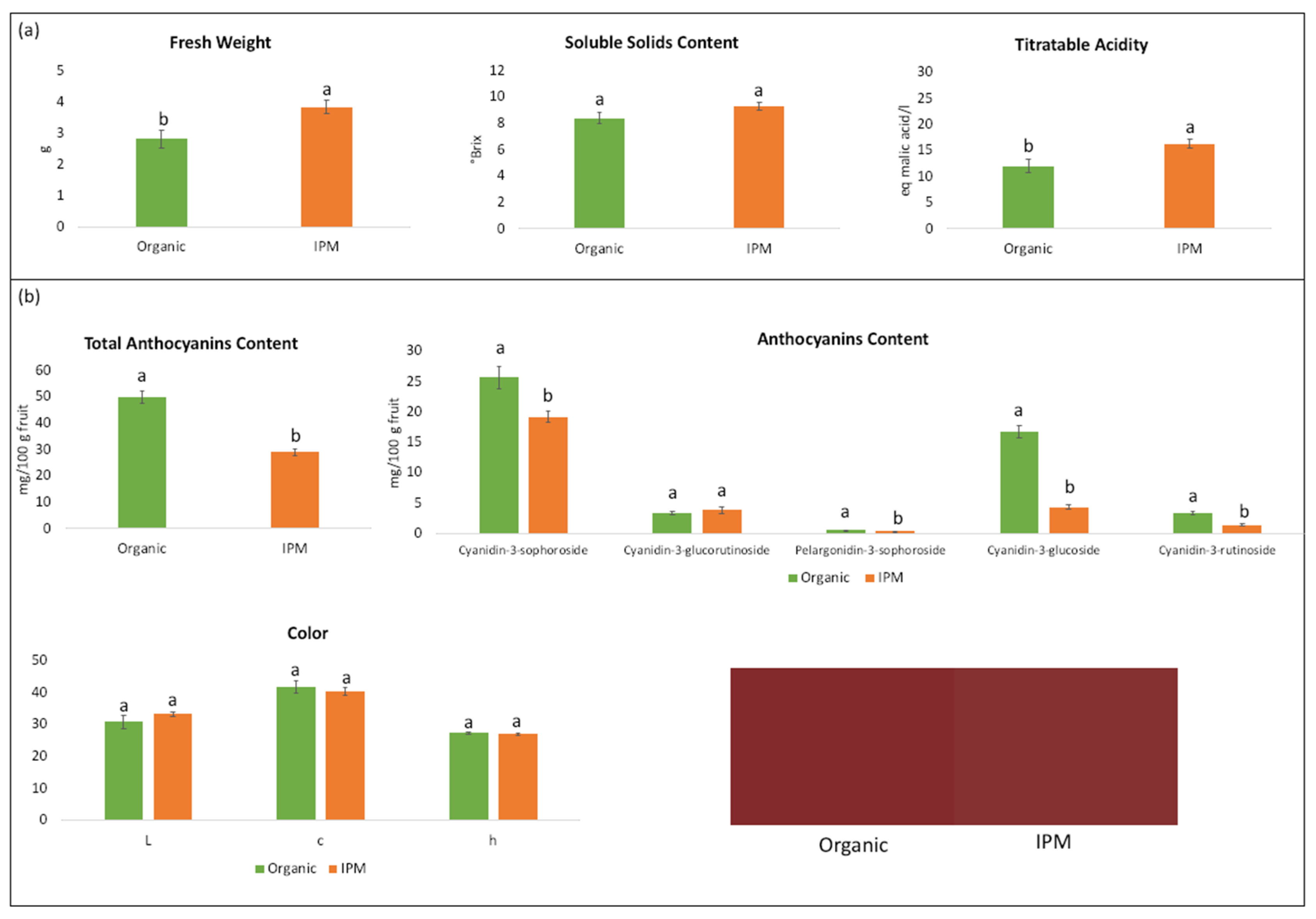
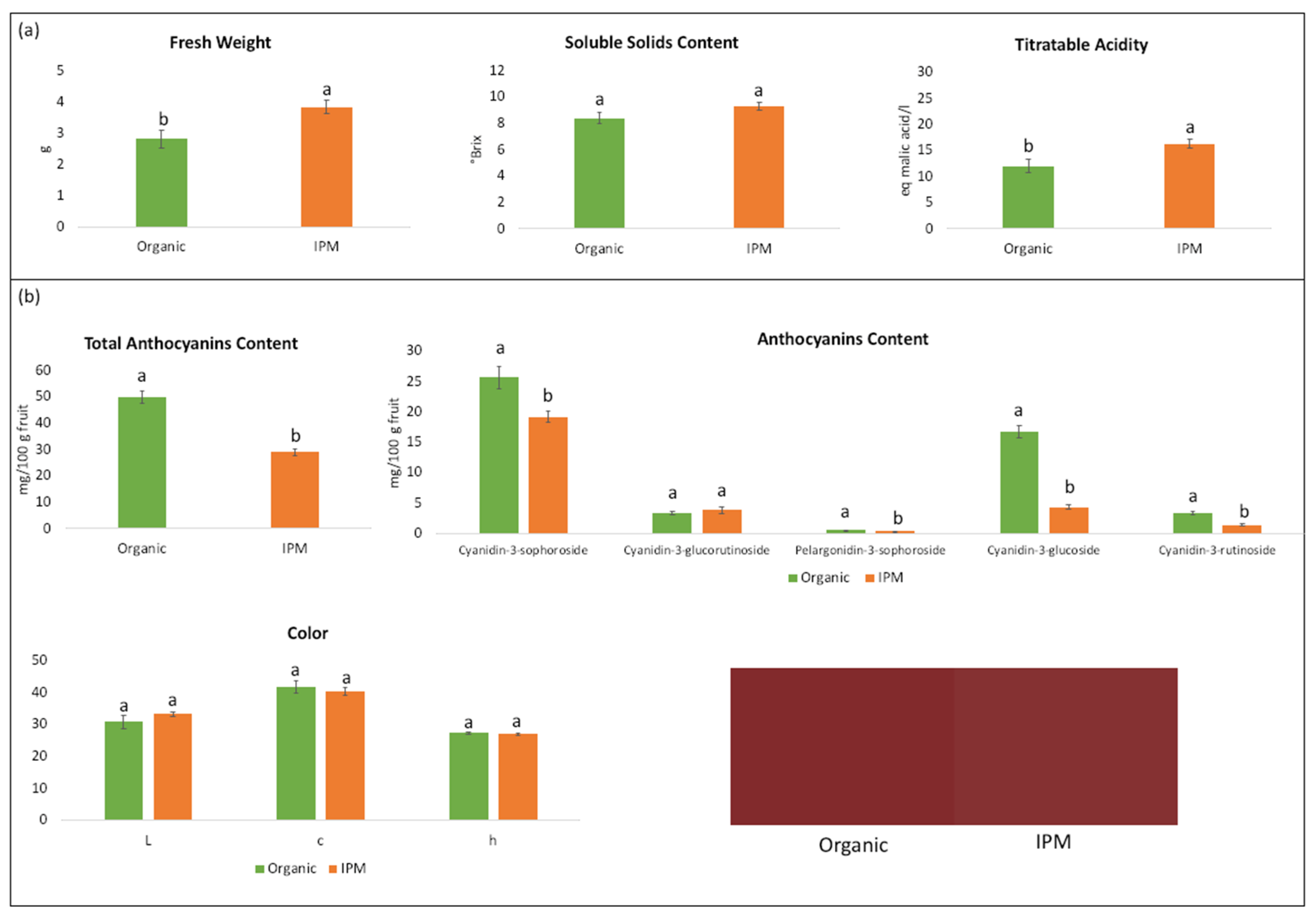
3.2. Fruit Quality Characteristics
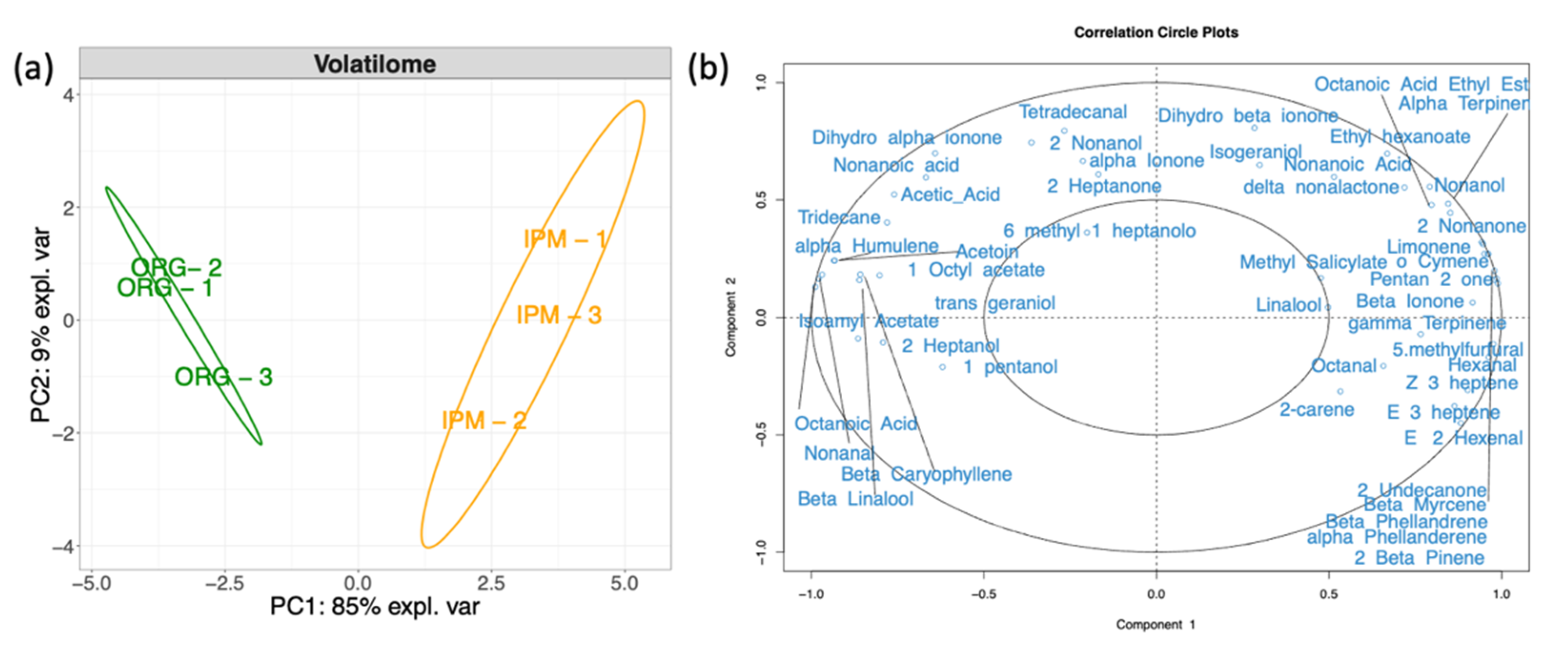
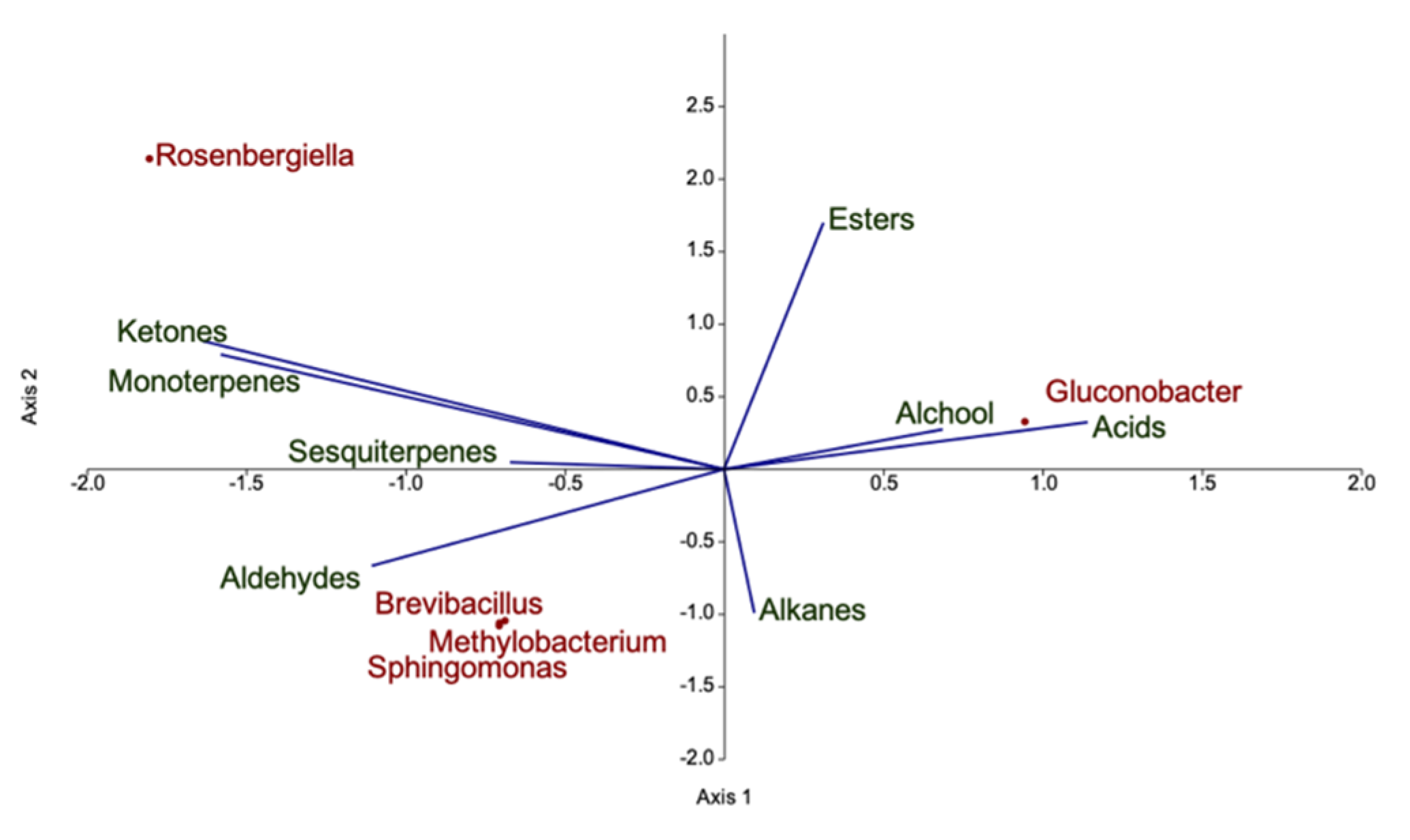
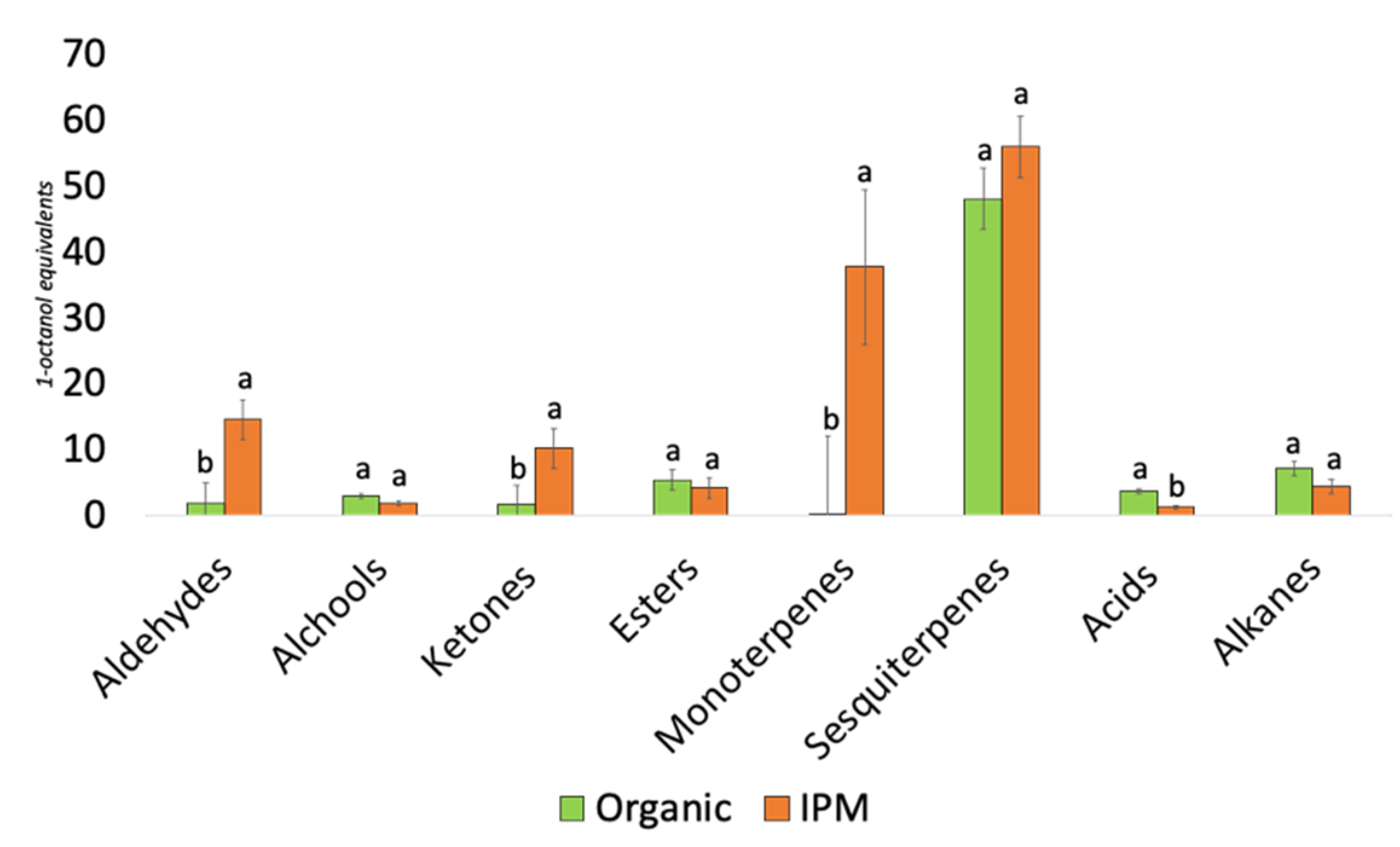
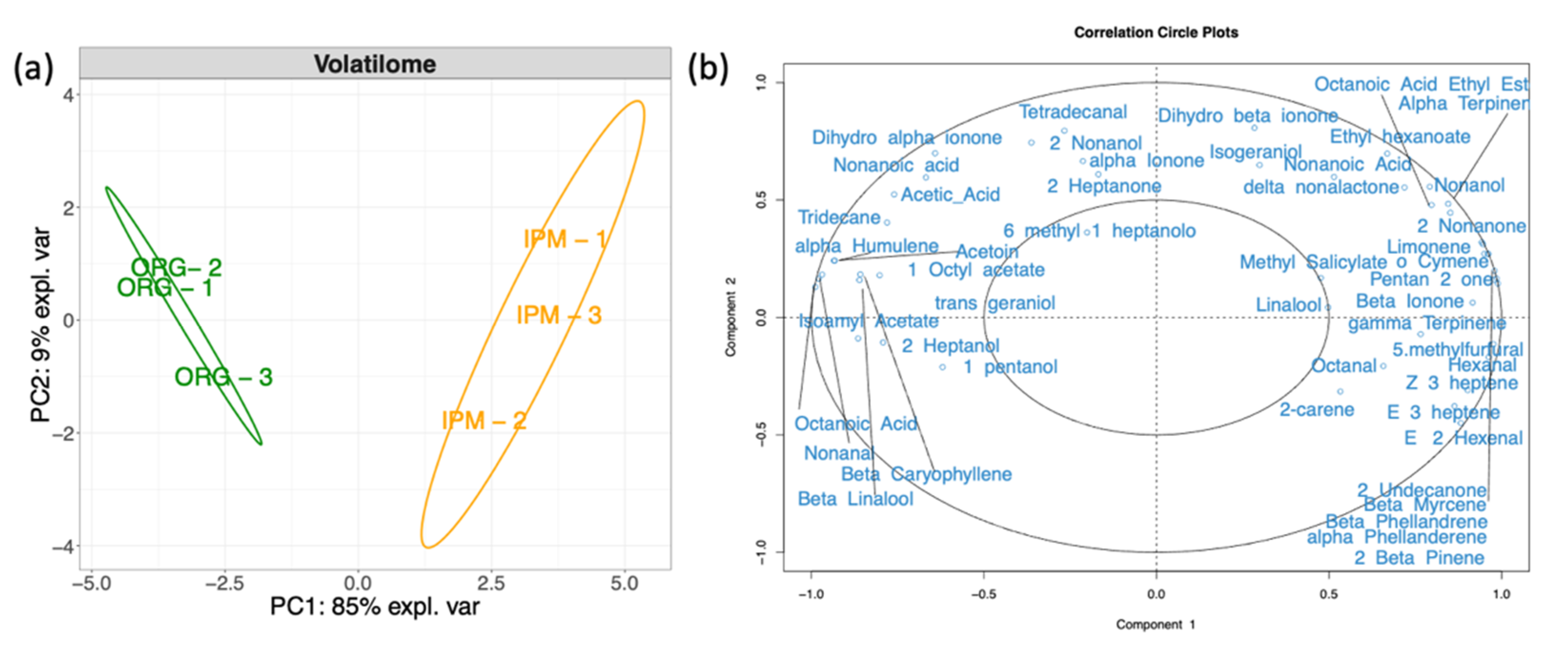
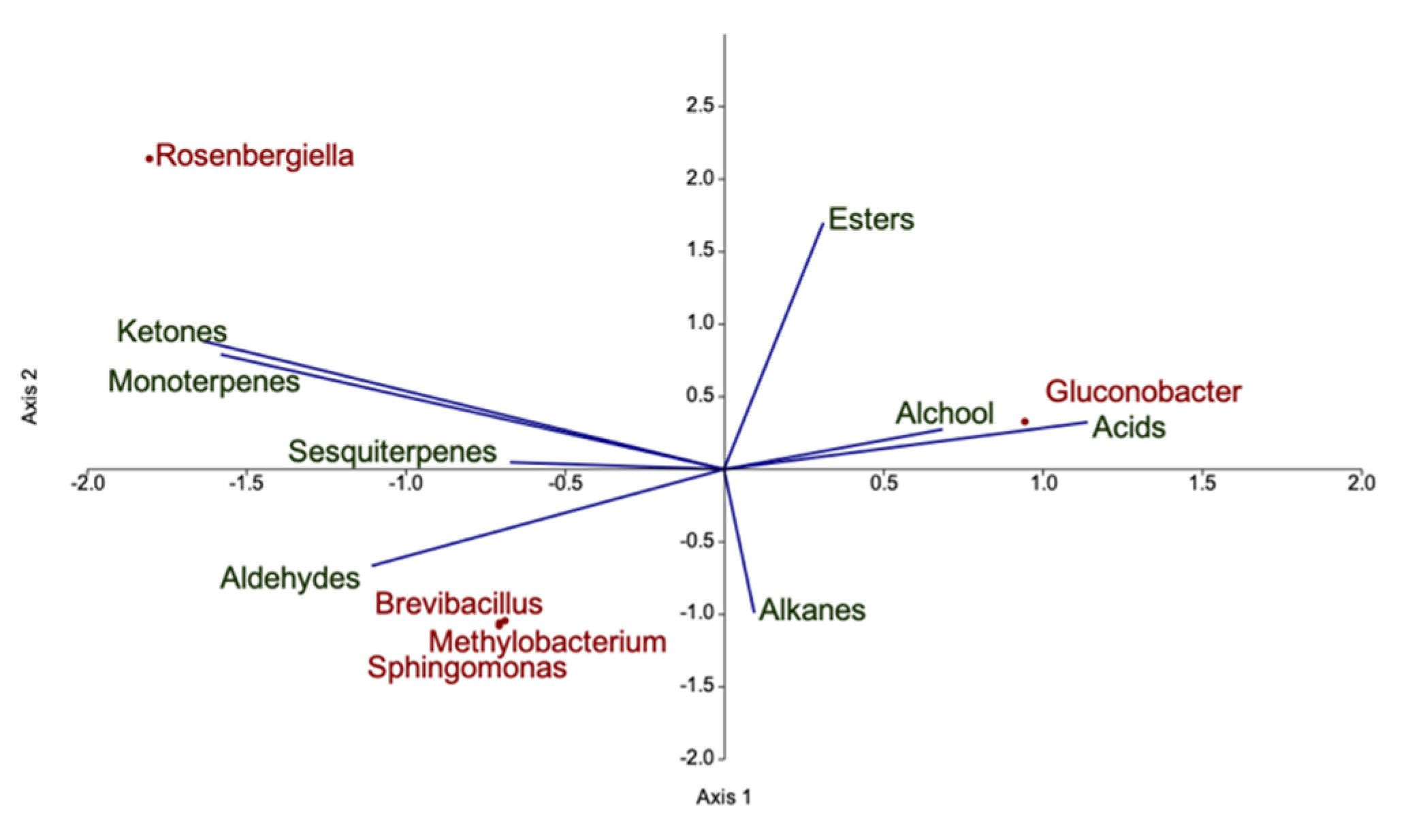
3.3. Characterization of the Volatilome of Organic and Integrated Pest Management (IPM) Raspberry Fruits
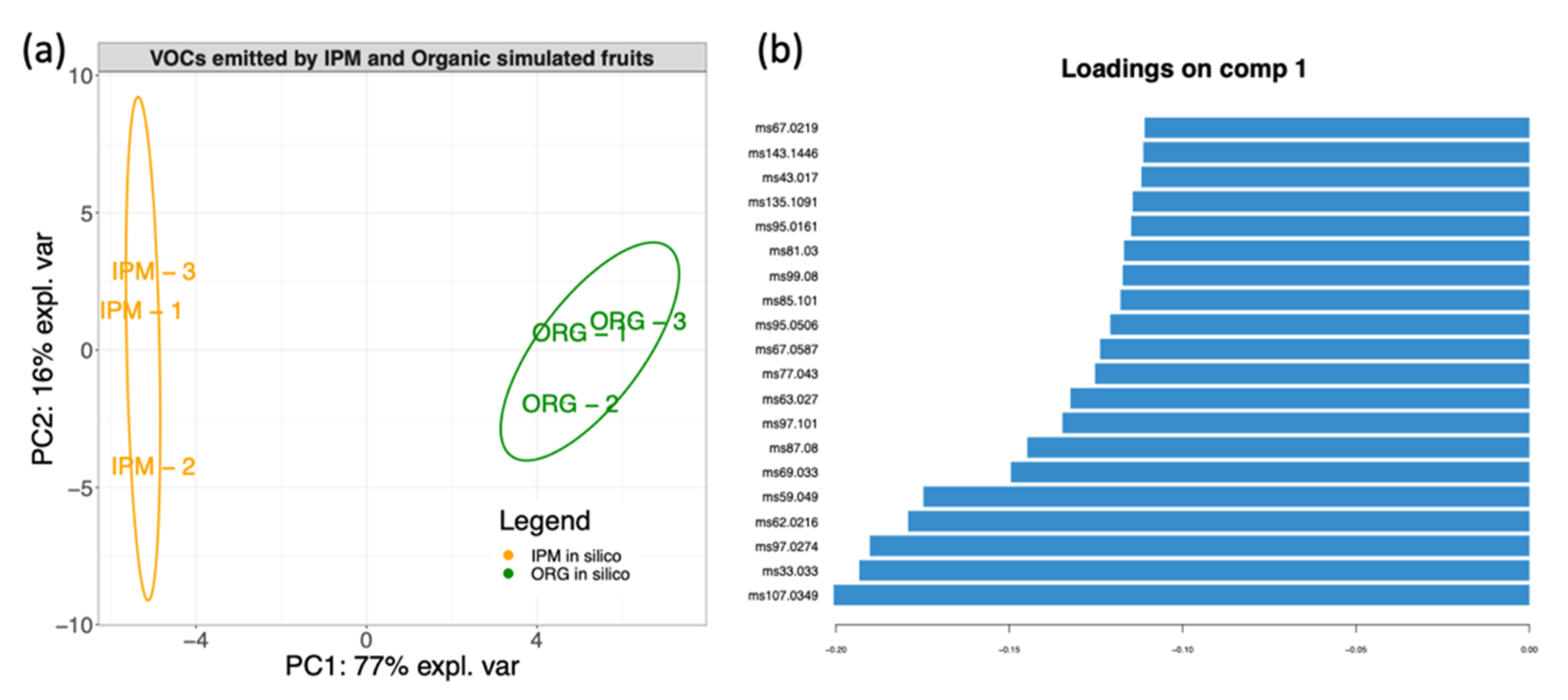
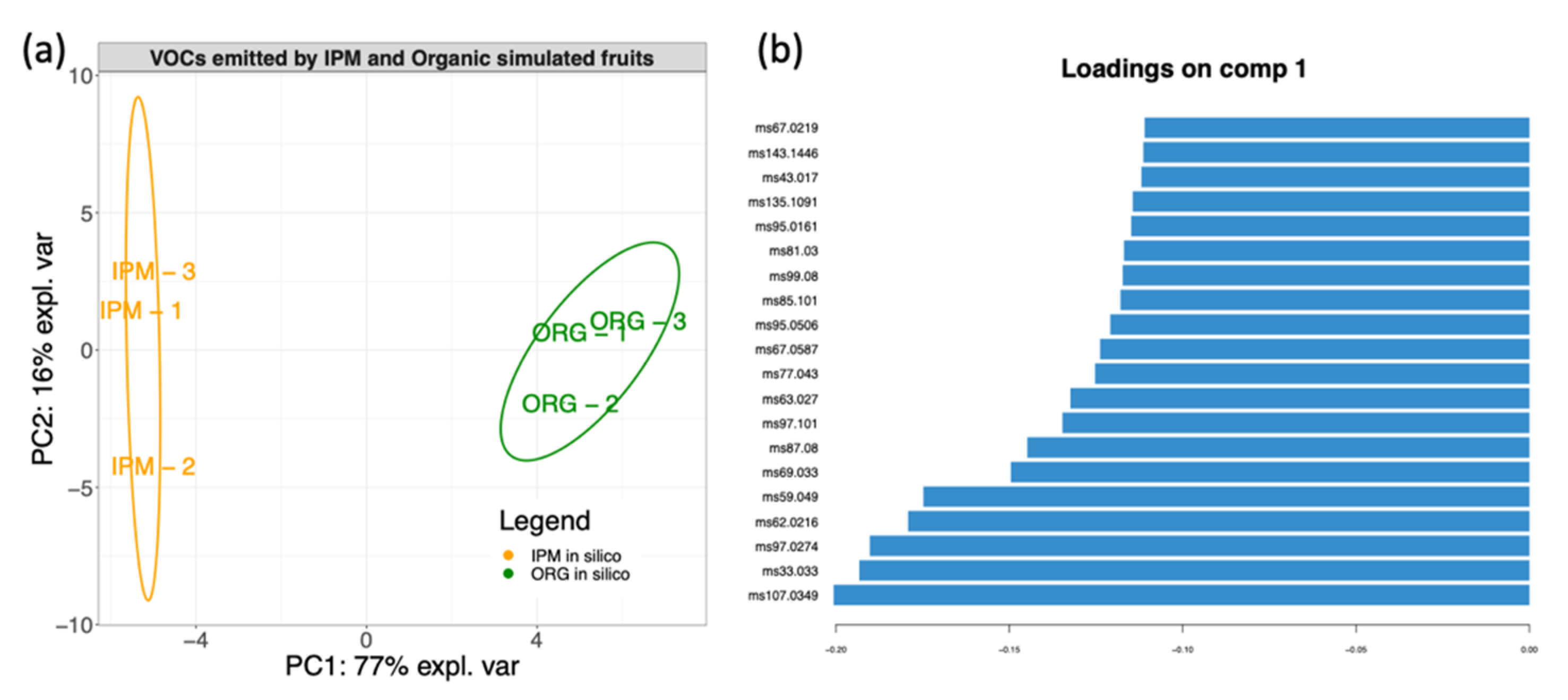
3.4. In Silico Fruit Volatilome Assembly
4. Discussion
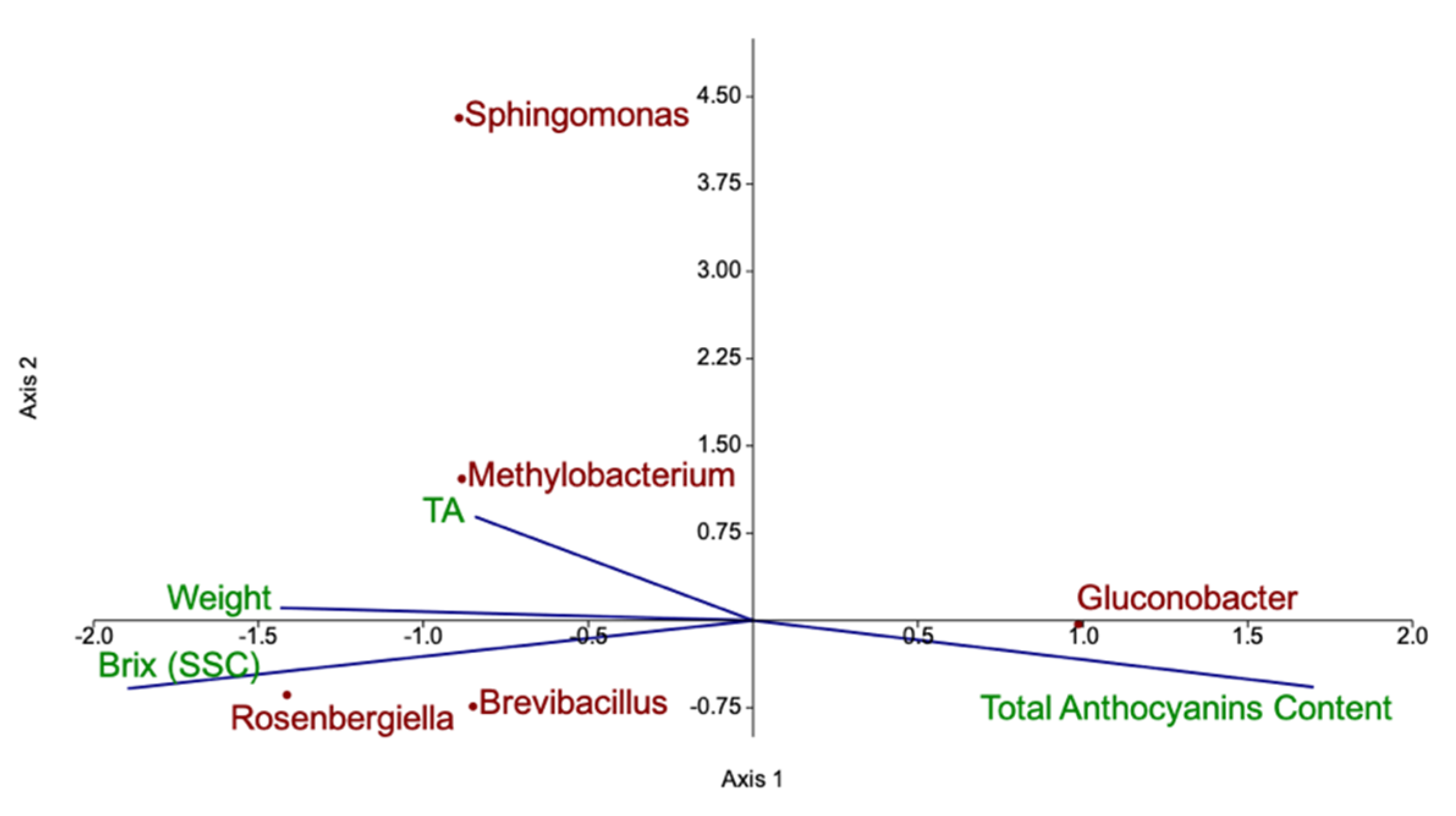
4.1. Diversity of Fruit-Associated Microbiomes
4.2. Quality Parameters of Organic and IPM Raspberry Fruits
4.3. Volatilome and Bacterial Contribution to Fruit Aroma
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT Database. Available online: http://www.fao.org/faostat/en/#home (accessed on 15 January 2021).
- Girgenti, V.; Peano, C.; Bounous, M.; Baudino, C. A life cycle assessment of non-renewable energy use and greenhouse gas emissions associated with blueberry and raspberry production in northern Italy. Sci. Total. Environ. 2013, 458, 414–418. [Google Scholar] [CrossRef] [Green Version]
- Valentinuzzi, F.; Pii, Y.; Mimmo, T.; Savini, G.; Curzel, S.; Cesco, S. Fertilization strategies as a tool to modify the organoleptic properties of raspberry (Rubus idaeus L.) fruits. Sci. Hortic. 2018, 240, 205–212. [Google Scholar] [CrossRef]
- Stavang, J.A.; Freitag, S.; Foito, A.; Verrall, S.; Heide, O.M.; Stewart, D.; Sønsteby, A. Raspberry fruit quality changes during ripening and storage as assessed by colour, sensory evaluation and chemical analyses. Sci. Hortic. 2015, 195, 216–225. [Google Scholar] [CrossRef]
- Aprea, E.; Biasioli, F.; Gasperi, F. Volatile Compounds of Raspberry Fruit: From Analytical Methods to Biological Role and Sensory Impact. Molecules 2015, 20, 2445–2474. [Google Scholar] [CrossRef] [Green Version]
- Aprea, E.; Biasioli, F.; Carlin, S.; Endrizzi, I.; Gasperi, F. Investigation of Volatile Compounds in Two Raspberry Cultivars by Two Headspace Techniques: Solid-Phase Microextraction/Gas Chromatography−Mass Spectrometry (SPME/GC−MS) and Proton-Transfer Reaction−Mass Spectrometry (PTR−MS). J. Agric. Food Chem. 2009, 57, 4011–4018. [Google Scholar] [CrossRef]
- Estrada-Beltran, A.; Salas-Salazar, N.A.; Parra-Quezada, R.A.; Gonzalez-Franco, A.C.; Soto-Caballero, M.C.; Rodriguez-Roque, M.J.; Flores-Cordova, M.A.; Chavez-Martinez, A. Effect of conventional and organic fertilizers on volatile compounds of raspberry fruit. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 862–870. [Google Scholar] [CrossRef]
- Zhang, W.; Lao, F.; Bi, S.; Pan, X.; Pang, X.; Hu, X.; Wu, J. Insights into the major aroma-active compounds in clear red raspberry juice (Rubus idaeus L. cv. Heritage) by molecular sensory science approaches. Food Chem. 2021, 336, 127721. [Google Scholar] [CrossRef] [PubMed]
- Lupatini, M.; Korthals, G.W.; De Hollander, M.; Janssens, T.K.S.; Kuramae, E.E. Soil Microbiome Is More Heterogeneous in Organic Than in Conventional Farming System. Front. Microbiol. 2017, 7, 2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyinade, G.A.; Mushunje, A.; Yusuf, S.F.G. The willingness to consume organic food: A review. Food Agric. Immunol. 2021, 32, 78–104. [Google Scholar] [CrossRef]
- Eurostat Database. Available online: https://ec.europa.eu/eurostat (accessed on 15 January 2021).
- Bodiroga, R.; Sredojević, Z. Economic Validity of Organic Raspberry Production as a Challenge for Producers in Bosnia and Herzegovina. Econ. Insights-Trends Chall. 2017, 69, 5–15. [Google Scholar]
- Frias-Moreno, M.N.; Olivas-Orozco, G.I.; Gonzalez-Aguilar, G.A.; Benitez-Enriquez, Y.E.; Paredes-Alonso, A.; Jacobo-Cuellar, J.L.; Salas-Salazar, N.A.; Ojeda-Barrios, D.L.; Parra-Quezada, R.A. Yield, Quality and Phytochemicals of Organic and Conventional Raspberry Cultivated in Chihuahua, Mexico. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 47, 522–530. [Google Scholar] [CrossRef] [Green Version]
- Barrel, I.; Diemoz, P. La Coltivazione del Lampone—Manuale Tecnico-Pratico; Insitut Agricole Regional: Aosta, Italy, 2011. [Google Scholar]
- Dolan, A.; Macfarlane, S.; Jennings, S.N. Pathogens in Raspberry and Other Rubus spp. Raspberry 2018, 41–61. [Google Scholar] [CrossRef]
- Perpetuini, G.; Donati, I.; Cellini, A.; Orrú, L.; Giongo, L.; Farneti, B.; Spinelli, F. Genetic and functional characterization of the bacterial community on fruit of three raspberry (Rubus idaeus) cultivars. J. Berry Res. 2019, 9, 227–247. [Google Scholar] [CrossRef]
- Hemmerling, S.; Hamm, U.; Spiller, A. Consumption behaviour regarding organic food from a marketing perspective—a literature review. Org. Agric. 2015, 5, 277–313. [Google Scholar] [CrossRef]
- Orsini, F.; Maggio, A.; Rouphael, Y.; De Pascale, S. Physiological quality” of organically grown vegetables. Sci. Hortic. 2016, 208, 131–139. [Google Scholar] [CrossRef]
- Funk, C.; Kennedy, B. The New Food Fights: US Public Divides over Food Science; Pew Research Center: Washington, DC, USA, 2016. [Google Scholar]
- Seufert, V.; Ramankutty, N.; Mayerhofer, T. What is this thing called organic? How organic farming is codified in regulations. Food Policy 2017, 68, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Di Vittori, L.; Mazzoni, L.; Battino, M.; Mezzetti, B. Pre-harvest factors influencing the quality of berries. Sci. Hortic. 2018, 233, 310–322. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Lemanceau, P.; Blouin, M.; Muller, D.; Moënne-Loccoz, Y. Let the Core Microbiota Be Functional. Trends Plant Sci. 2017, 22, 583–595. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Rocha-Granados, M.D.C.; Glick, B.R.; Santoyo, G. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol. Res. 2018, 208, 25–31. [Google Scholar] [CrossRef]
- Andreote, F.D.; Pereira, M.D.C.; Silva, M.D.C.P. Microbial communities associated with plants: Learning from nature to apply it in agriculture. Curr. Opin. Microbiol. 2017, 37, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Sangiorgio, D.; Cellini, A.; Donati, I.; Pastore, C.; Onofrietti, C.; Spinelli, F. Facing Climate Change: Application of Microbial Biostimulants to Mitigate Stress in Horticultural Crops. Agronomy 2020, 10, 794. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Malacrinò, A.; Wisniewski, M.; Cacciola, S.O.; Schena, L. Metabarcoding: A powerful tool to investigate microbial communities and shape future plant protection strategies. Biol. Control. 2018, 120, 1–10. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; Celador-Lera, L.; Fradejas-Bayón, M.; Rivas, R. Plant probiotic bacteria enhance the quality of fruit and horticultural crops. AIMS Microbiol. 2017, 3, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Wassermann, B.; Müller, H.; Berg, G. An Apple a Day: Which Bacteria Do We Eat with Organic and Conventional Apples? Front. Microbiol. 2019, 10, 1629. [Google Scholar] [CrossRef] [Green Version]
- Mditshwa, A.; Magwaza, L.; Tesfay, S.Z.; Mbili, N. Postharvest quality and composition of organically and conventionally produced fruits: A review. Sci. Hortic. 2017, 216, 148–159. [Google Scholar] [CrossRef]
- Ishaq, S.; Johnson, S.P.; Miller, Z.J.; Lehnhoff, E.A.; Olivo, S.; Yeoman, C.J.; Menalled, F.D. Impact of Cropping Systems, Soil Inoculum, and Plant Species Identity on Soil Bacterial Community Structure. Microb. Ecol. 2016, 73, 417–434. [Google Scholar] [CrossRef]
- Suzuki, K.; Takemura, M.; Miki, T.; Nonaka, M.; Harada, N. Differences in soil bacterial community compositions in paddy fields under organic and IPM farming conditions. Microbes Environ. 2019, 34, 108–111. [Google Scholar] [CrossRef]
- Bill, M.; Chidamba, L.; Gokul, J.K.; Labuschagne, N.; Korsten, L. Bacterial community dynamics and functional profiling of soils from IPM and organic cropping systems. Appl. Soil Ecol. 2021, 157, 103734. [Google Scholar] [CrossRef]
- Maguire, T.L.; Collins, G.G.; Sedgley, M. A modified CTAB DNA extraction procedure for plants belonging to the family proteaceae. Plant Mol. Biol. Rep. 1994, 12, 106–109. [Google Scholar] [CrossRef]
- Bushnell, B. BBMap: A Fast, Accurate, Splice-Aware Aligner. No. LBNL-7065E; Lawrence Berkeley National Lab. (LBNL): Berkeley, CA, USA, 2014. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- De Urraza, P.J.; Gómez-Zavaglia, A.; Lozano, M.E.; Romanowski, V.; De Antoni, G.L. DNA fingerprinting of thermophilic lactic acid bacteria using repetitive sequence-based polymerase chain reaction. J. Dairy Res. 2000, 67, 381–392. [Google Scholar] [CrossRef]
- Gevers, D.; Huys, G.; Swings, J. Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. FEMS Microbiol. Lett. 2001, 205, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Filippetti, I.; Intrieri, C.; Centinari, M.; Bucchetti, B.; Pastore, C. Molecular characterization of officially registered Sangiovese clones and of other Sangiovese-like biotypes in Tuscany, Corsica and Emilia-Romagna. Vitis-Geilweilerhof 2005, 44, 167. [Google Scholar]
- Bringel, F.; Castioni, A.; Olukoya, D.K.; Felis, G.E.; Torriani, S.; Dellaglio, F. Lactobacillus plantarum subsp. argentoratensis subsp. nov., isolated from vegetable matrices. Int. J. Syst. Evol. Microbiol. 2005, 55, 1629–1634. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Ahmad, I.; Khan, M. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Blomqvist, K.; Nikkola, M.; Lehtovaara, P.; Suihko, M.L.; Airaksinen, U.; Stråby, K.B.; Penttilä, M.E. Characterization of the genes of the 2, 3-butanediol operons from Klebsiella terrigena and Enterobacter aerogenes. J. Bacteriol. 1993, 175, 1392–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappuccino, J.C.; Sherman, N. Microbiology: A Laboratory Manual, 3rd ed.; Benjamin/cummings Pub. Co.: New York, NY, USA, 1992. [Google Scholar]
- Pérez-Miranda, S.; Cabirol, N.; George-Téllez, R.; Zamudio-Rivera, L.; Fernández, F. O-CAS, a fast and universal method for siderophore detection. J. Microbiol. Methods 2007, 70, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Mattivi, F.; Guzzon, R.; Vrhovsek, U.; Stefanini, A.M.; Velasco, R. Metabolite Profiling of Grape: Flavonols and Anthocyanins. J. Agric. Food Chem. 2006, 54, 7692–7702. [Google Scholar] [CrossRef]
- De Ancos, B.; Gonzalez, E.; Cano, M.P. Differentiation of raspberry varieties according to anthocyanin composition. Z. Lebensm. Forschung A 1999, 208, 33–38. [Google Scholar] [CrossRef]
- Farneti, B.; Khomenko, I.; Grisenti, M.; Ajelli, M.; Betta, E.; Algarra, A.A.; Cappellin, L.; Aprea, E.; Gasperi, F.; Biasioli, F.; et al. Exploring Blueberry Aroma Complexity by Chromatographic and Direct-Injection Spectrometric Techniques. Front. Plant Sci. 2017, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Rohart, F.; Gautier, B.; Singh, A.; Cao, K.-A.L. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Hartman, K.; Van Der Heijden, M.G.A.; Wittwer, R.A.; Banerjee, S.; Walser, J.-C.; Schlaeppi, K. Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef]
- Remus-Emsermann, M.N.P.; Schlechter, R.O. Phyllosphere microbiology: At the interface between microbial individuals and the plant host. New Phytol. 2018, 218, 1327–1333. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S. The postharvest microbiome: The other half of sustainability. Biol. Control. 2019, 137, 104025. [Google Scholar] [CrossRef]
- Lenaerts, M.; Álvarez-Pérez, S.; De Vega, C.; Van Assche, A.; Johnson, S.D.; Willems, K.A.; Herrera, C.M.; Jacquemyn, H.; Lievens, B. Rosenbergiella australoborealis sp. nov., Rosenbergiella collisarenosi sp. nov. and Rosenbergiella epipactidis sp. nov., three novel bacterial species isolated from floral nectar. Syst. Appl. Microbiol. 2014, 37, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Hommel, R. Gluconobacter. In Encyclopedia of Food Microbiology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 2, pp. 99–105. [Google Scholar]
- Van Keer, C.; Abeele, P.V.; Swings, J.; Gosselé, F.; De Ley, J. Acetic acid bacteria as causal agents of browning and rot of apples and pears. Zentralbl. Bakteriol. Parasitenkd. Hyg. 1981, 2, 197–204. [Google Scholar] [CrossRef]
- Kemperman, R.A.; Gross, G.; Mondot, S.; Possemiers, S.; Marzorati, M.; Van de Wiele, T.; Doré, J.; Vaughan, E.E. Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome. Food Res. Int. 2013, 53, 659–669. [Google Scholar] [CrossRef]
- Kremen, C.; Miles, A. Ecosystem Services in Biologically Diversified versus Conventional Farming Systems: Benefits, Externalities, and Trade-Offs. Ecol. Soc. 2012, 17, 174. [Google Scholar] [CrossRef]
- Lori, M.; Symnaczik, S.; Mäder, P.; De Deyn, G.; Gattinger, A. Organic farming enhances soil microbial abundance and activity—A meta-analysis and meta-regression. PLoS ONE 2017, 12, e0180442. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, V.K.; Qazi, G.N.; Kumar, A. Gluconobacter oxydans: Its biotechnological applications. J. Mol. Microbiol. Biotechnol. 2001, 3, 445–456. [Google Scholar] [PubMed]
- Cernava, T.; Chen, X.; Krug, L.; Li, H.; Yang, M.; Berg, G. The tea leaf microbiome shows specific responses to chemical pesticides and biocontrol applications. Sci. Total. Environ. 2019, 667, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Crecente-Campo, J.; Nunes-Damaceno, M.; Romero-Rodríguez, M.; Vázquez-Odériz, M. Color, anthocyanin pigment, ascorbic acid and total phenolic compound determination in organic versus conventional strawberries (Fragaria×ananassa Duch, cv Selva). J. Food Compos. Anal. 2012, 28, 23–30. [Google Scholar] [CrossRef]
- Dastogeer, K.M.; Tumpa, F.H.; Sultana, A.; Akter, M.A.; Chakraborty, A. Plant microbiome–an account of the factors that shape community composition and diversity. Curr. Plant Biol. 2020, 23, 100161. [Google Scholar] [CrossRef]
- Ponder, A.; Hallmann, E. The effects of organic and conventional farm management and harvest time on the polyphenol content in different raspberry cultivars. Food Chem. 2019, 301, 125295. [Google Scholar] [CrossRef]
- Bernales, M.; Monsalve, L.; Ayala-Raso, A.; Valdenegro, M.; Martinez, J.P.; Travisany, D.; Defilippi, B.; González-Agüero, M.; Cherian, S.; Fuentes, L. Expression of two indole-3-acetic acid (IAA)-amido synthetase (GH3) genes during fruit development of raspberry (Rubus idaeus Heritage). Sci. Hortic. 2019, 246, 168–175. [Google Scholar] [CrossRef]
- Fuentes, L.; Monsalve, L.; Morales-Quintana, L.; Valdenegro, M.; Martínez, J.-P.; Defilippi, B.G.; González-Agüero, M. Differential expression of ethylene biosynthesis genes in drupelets and receptacle of raspberry (Rubus idaeus). J. Plant Physiol. 2015, 179, 100–105. [Google Scholar] [CrossRef]
- Iannetta, P.; Berg, J.V.D.; Wheatley, R.; McMillan, G.; McNicol, R.; Davies, H. A CAUSAL ROLE FOR ETHENE IN RASPBERRY FRUIT RIPENING. Acta Hortic. 1999, 393–400. [Google Scholar] [CrossRef]
- Teribia, N.; Tijero, V.; Munné-Bosch, S. Linking hormonal profiles with variations in sugar and anthocyanin contents during the natural development and ripening of sweet cherries. New Biotechnol. 2016, 33, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Moro, L.; Hassimotto, N.M.A.; Purgatto, E. Postharvest Auxin and Methyl Jasmonate Effect on Anthocyanin Biosynthesis in Red Raspberry (Rubus idaeus L.). J. Plant Growth Regul. 2017, 36, 773–782. [Google Scholar] [CrossRef]
- Faller, A.; Fialho, E. The antioxidant capacity and polyphenol content of organic and conventional retail vegetables after domestic cooking. Food Res. Int. 2009, 42, 210–215. [Google Scholar] [CrossRef]
- Anjos, R.; Cosme, F.; Gonçalves, A.; Nunes, F.M.; Vilela, A.; Pinto, T. Effect of agricultural practices, conventional vs organic, on the phytochemical composition of ‘Kweli’and ‘Tulameen’raspberries (Rubus idaeus L.). Food Chem. 2020, 328, 126833. [Google Scholar] [CrossRef] [PubMed]
- Rembiałkowska, E. Quality of plant products from organic agriculture. J. Sci. Food Agric. 2007, 87, 2757–2762. [Google Scholar] [CrossRef]
- Wang, N.; Qu, C.; Jiang, S.; Chen, Z.; Xu, H.; Fang, H.; Su, M.; Zhang, J.; Wang, Y.; Liu, W.; et al. The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples. Plant J. 2018, 96, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-F.; An, J.-P.; Liu, X.; Su, L.; You, C.-X.; Hao, Y.-J. The Nitrate-Responsive Protein MdBT2 Regulates Anthocyanin Biosynthesis by Interacting with the MdMYB1 Transcription Factor. Plant Physiol. 2018, 178, 890–906. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Nakayama, M.; Shigeta, J.-I. Culturing conditions affecting the production of anthocyanin in suspended cell cultures of strawberry. Plant Sci. 1996, 113, 91–98. [Google Scholar] [CrossRef]
- Prinsi, B.; Negrini, N.; Morgutti, S.; Espen, L. Nitrogen Starvation and Nitrate or Ammonium Availability Differently Affect Phenolic Composition in Green and Purple Basil. Agronomy 2020, 10, 498. [Google Scholar] [CrossRef] [Green Version]
- Etienne, A.; Génard, M.; Lobit, P.; Mbeguié-A-Mbéguié, D.; Bugaud, C. What controls fleshy fruit acidity? A review of malate and citrate accumulation in fruit cells. J. Exp. Bot. 2013, 64, 1451–1469. [Google Scholar] [CrossRef] [Green Version]
- Arikan, Ş.; Pirlak, L. Effects of Plant Growth Promoting Rhizobacteria (PGPR) on Growth, Yield and Fruit Quality of Sour Cherry (Prunus cerasus L.). Erwerbs-Obstbau 2016, 58, 221–226. [Google Scholar] [CrossRef]
- Todeschini, V.; AitLahmidi, N.; Mazzucco, E.; Marsano, F.; Gosetti, F.; Robotti, E.; Bona, E.; Massa, N.; Bonneau, L.; Marengo, E.; et al. Impact of Beneficial Microorganisms on Strawberry Growth, Fruit Production, Nutritional Quality, and Volatilome. Front. Plant Sci. 2018, 9, 1611. [Google Scholar] [CrossRef]
- Kutschera, U. Plant-associated methylobacteria as co-evolved phytosymbionts: A hypothesis. Plant Signal. Behav. 2007, 2, 74–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.L.; Waqas, M.; Kang, S.M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.-Y.; et al. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, D.N.; Ekimova, G.A.; Doronina, N.V.; Trotsenko, Y.A. 1-Aminocyclopropane-1-carboxylate (ACC) deaminases from Methylobacterium radiotolerans and Methylobacterium nodulans with higher specificity for ACC. FEMS Microbiol. Lett. 2013, 343, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Lidstrom, M.E. Metabolite profiling analysis ofMethylobacterium extorquens AM1 by comprehensive two-dimensional gas chromatography coupled with time-of-flight mass spectrometry. Biotechnol. Bioeng. 2008, 99, 929–940. [Google Scholar] [CrossRef]
- Laviad-Shitrit, S.; Izhaki, I.; Whitman, W.B.; Shapiro, N.; Woyke, T.; Kyrpides, N.C.; Halpern, M. Draft genome of Rosenbergiella nectarea strain 8N4T provides insights into the potential role of this species in its plant host. PeerJ 2020, 8, e8822. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.; Simpson, C. Developmental Transitions to Fruiting in Red Raspberry. In Compendium of Plant Genomes; Springer Science and Business Media LLC.: Berlin/Heidelberg, Germany, 2018; pp. 199–212. [Google Scholar]
- Ray, S.; Amin, D.; Patel, N. Microbial Mitigation of Salinity and Water Stress in Legume Crops. In Microbial Mitigation of Stress Response of Food Legumes; Informa UK Limited: London, UK, 2020; pp. 165–175. [Google Scholar]
- Ratón, T.D.L.M.O.; Vinent, S.N.; Díaz, M.D.J.S.; Delgado, E.O.; Barbosa, H.R. The New Rhizospheric Bacteria Brevibacillus Benefits Eggplant and Peeper Growth and Productivity Under Organoponic System. Agric. Res. 2014, 3, 395–398. [Google Scholar] [CrossRef]
- Halpern, M.; Fridman, S.; Atamna-Ismaeel, N.; Izhaki, I. Rosenbergiella nectarea gen. nov., sp. nov., in the family Enterobacteriaceae, isolated from floral nectar. Int. J. Syst. Evol. Microbiol. 2013, 63, 4259–4265. [Google Scholar] [CrossRef]
- Vannette, R.L. The Floral Microbiome: Plant, Pollinator, and Microbial Perspectives. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 363–386. [Google Scholar] [CrossRef]
- Dong, C.-J.; Wang, L.-L.; Li, Q.; Shang, Q.-M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Torres-Cortés, G.; Bonneau, S.; Bouchez, O.; Genthon, C.; Briand, M.; Jacques, M.-A.; Barret, M. Functional Microbial Features Driving Community Assembly During Seed Germination and Emergence. Front. Plant Sci. 2018, 9, 902. [Google Scholar] [CrossRef] [Green Version]
- Caneschi, W.L.; Felestrino, É.B.; Fonseca, N.; Villa, M.M.; Lemes, C.G.D.C.; Cordeiro, I.F.; Assis, R.D.A.B.; Sanchez, A.B.; Vieira, I.T.; Kamino, L.H.Y.; et al. Brazilian Ironstone Plant Communities as Reservoirs of Culturable Bacteria with Diverse Biotechnological Potential. Front. Microbiol. 2018, 9, 1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhou, J.; Li, Y.; Wen, J.; Wang, R. Bacterial endophytes from Lycoris radiata promote the accumulation of Amaryllidaceae alkaloids. Microbiol. Res. 2020, 239, 126501. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, L.; Figueroa, C.R.; Valdenegro, M. Recent Advances in Hormonal Regulation and Cross-Talk during Non-Climacteric Fruit Development and Ripening. Horticulturae 2019, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.; Poll, L.; Callesen, O.; Lewis, M. Relations between the Content of Aroma Compounds and the Sensory Evaluation of 10 Raspberry Varieties (Rubus idaeus L.). Acta Agric. Scand. 1991, 41, 447–454. [Google Scholar] [CrossRef]
- Barnard, D.R.; Xue, R.-D. Laboratory Evaluation of Mosquito Repellents AgainstAedes albopictus, Culex nigripalpus, andOchlerotatus triseriatus(Diptera: Culicidae). J. Med. Entomol. 2004, 41, 726–730. [Google Scholar] [CrossRef] [Green Version]
- Ukeh, D.A.; Umoetok, S.B. Repellent effects of five monoterpenoid odours against Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.) in Calabar, Nigeria. Crop. Prot. 2011, 30, 1351–1355. [Google Scholar] [CrossRef]
- Cappellin, L.; Aprea, E.; Granitto, P.; Romano, A.; Gasperi, F.; Biasioli, F. Multiclass methods in the analysis of metabolomic datasets: The example of raspberry cultivar volatile compounds detected by GC–MS and PTR-MS. Food Res. Int. 2013, 54, 1313–1320. [Google Scholar] [CrossRef] [Green Version]
- Landi, M.; Araniti, F.; Flamini, G.; Piccolo, E.L.; Trivellini, A.; Abenavoli, M.R.; Guidi, L. “Help is in the air”: Volatiles from salt-stressed plants increase the reproductive success of receivers under salinity. Planta 2020, 251, 48. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Leifert, C.; Sanderson, R.; Seal, C.J. Agroecosystem Management and Nutritional Quality of Plant Foods: The Case of Organic Fruits and Vegetables. Crit. Rev. Plant Sci. 2011, 30, 177–197. [Google Scholar] [CrossRef]
- Cellini, A.; Spinelli, F.; Donati, I.; Ryu, C.-M.; Kloepper, J.W. Bacterial volatile compound-based tools for crop management and quality. Trends Plant Sci. 2021. [Google Scholar] [CrossRef]
- Amaresan, N.; Kumar, M.S.; Annapurna, K.; Kumar, K.; Sankaranarayanan, A. Beneficial Microbes in Agro-Ecology: Bacteria and Fungi; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Verginer, M.; Siegmund, B.; Cardinale, M.; Müller, H.; Choi, Y.; Míguez, C.B.; Leitner, E.; Berg, G. Monitoring the plant epiphyte Methylobacterium extorquens DSM 21961 by real-time PCR and its influence on the strawberry flavor. FEMS Microbiol. Ecol. 2010, 74, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Pohjanen, J.; Koskimäki, J.J.; Zabetakis, I.; Pirttilä, A.M. Localization of strawberry (Fragaria x ananassa) and Methylobacterium extorquens genes of strawberry flavor biosynthesis in strawberry tissue by in situ hybridization. J. Plant Physiol. 2014, 171, 1099–1105. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fruit Type | Bacterial Isolate |
|---|---|
| Both IPM and Organic | Bacillus subtilis, Burkholderia spp., Cellulomonas spp., Enterobacter asburiae, Enterobacter spp.—strain 1, Erwinia aphidicola, Erwinia rhapontici, Erwinia toletana, Lactobacillus plantarum, Methylobacterium extorquens, Ochrobactrum intermedium, Ochrobactrum pseudogrignonense, Paenibacillus alvei, Paenibacillus macerans, Paenibacillus polymixa, Pantoea agglomerans.—strain 1, Pantoea ananatis, Pantoea rwadensis, Pseudomonas fluorescens—strain 1, Pseudomonas fluorescens—strain 2, Pseudomonas stutzeri, Sphingomonas camponoticapitis, Sphingomonas paucimobilis, |
| IPM | Rosenbergiella spp. Enterobacter spp—strain 2 and 3 |
| Organic | Pantoea agglomerans—strain 2 Tatumella punctata Tatumella terrea Cronobacter spp. Klebsiella oxytoca |
| Cultivation Method | Bacterium | IAA Production | Acetoin Production | NH4+ Production | Siderphores Production | ACC Deaminase Activity |
|---|---|---|---|---|---|---|
| IPM | Rosenbergiella spp. | - | - | - | - | - |
| Enterobacter strain 2 | - | + | - | + | - | |
| Enterobacter strain 3 | - | - | + | + | - | |
| ORG | Pantoea agglomerans strain 2 | ++ | - | + | - | - |
| Cronobacter spp. | - | - | + | - | - | |
| Klebsiella oxytoca | - | - | + | + | - | |
| Tatumella punctata | - | - | + | - | - | |
| Tatumella terrea | ++ | - | + | - | - |
| Class | Compound | RetentionTime (min) | Linear Retention Index | Mean of Volatile Compounds Concentration (1-Octanol Equivalents) | |
|---|---|---|---|---|---|
| Org | IPM | ||||
| Aldehydes | (E)-2-Hexenal | 11.89 | 1192 | 0.25 ± 0.25 a | 6.82 ± 2.35 a |
| Hexanal | 5.34 | 1067 | nd | 5.65 ± 0.75 | |
| Octanal | 15.95 | 1274 | 0.47 ± 0.07 b | 0.76 ± 0.11 a | |
| Tetradecanal | 37.71 | 1942 | 0.63 ± 0.14 a | 0.44 ± 0.22 a | |
| 5-Methylfurfural | 27.51 | 1578 | nd | 0.81 ± 0.03 | |
| Nonanal | 20.46 | 1376 | 0.45 ± 0.03 | nd | |
| Alchools | 6-Methyl-1-heptanol | 18.11 | 1319 | 0.06 ± 0.03 a | 0.03 ± 0.03 a |
| 1-Pentanol | 14.27 | 1241 | 0.14 ± 0.07 | nd | |
| 2-Nonanol | 29.29 | 1508 | 0.10 ± 0.01 a | 0.05 ± 0.05 a | |
| 2-Heptanol | 17.82 | 1312 | 0.84 ± 0.14 a | 0.43 ± 0.08 b | |
| β -Linalool | 26.13 | 1535 | 0.55 ± 0.14 a | 0.05 ± 0.05 a | |
| Isogeraniol | 40.33 | 2039 | 0.21 ± 0.11 a | 0.33 ± 0.24 a | |
| Nonanol | 29.56 | 1645 | nd | 0.18 ± 0.09 | |
| Trans-geraniol | 34.71 | 1827 | 0.99 ± 0.08 a | 0.67 ± 0.09 b | |
| Ketones | Acetoin | 15.23 | 1259 | 0.31 ± 0.11 | nd |
| 2-Heptanone | 9.99 | 1136 | 0.60 ± 0.20 a | 0.45 ± 0.16 a | |
| 2-Nonanone | 20.27 | 1372 | 0.38 ± 0.10 b | 1.08 ± 0.27 a | |
| 2-Undecanone | 27.58 | 1581 | 0.18 ± 0.04 b | 1.15 ± 0.24 a | |
| Pentan-2-one | 3.91 | 1012 | nd | 7.35 ± 2.35 | |
| Esters | 1-Octyl acetate | 23.71 | 1463 | 3.05 ± 0.19 a | 0.75 ± 0.15 b |
| Ethyl hexanoate | 13.47 | 1225 | 0.71 ± 0.08 a | 2.26 ± 1.13 a | |
| Isoamyl Acetate | 6.97 | 1130 | 0.84 ± 0.24 | nd | |
| Methyl Salicylate | 32.75 | 1756 | 0.66 ± 0.33 a | 1.02 ± 0.31 a | |
| Sesquiterpenes | β-Caryophyllene | 27.21 | 1569 | 5.48 ± 1.38 | nd |
| α-Humulene | 29.39 | 1640 | 1.34 ± 0.43 | nd | |
| α-Ionone | 34.5 | 1819 | 13.99 ± 1.16 a | 13.35 ± 0.80 a | |
| β-Ionone | 36.79 | 1906 | 19.86 ± 1.97 b | 35.00 ± 2.49 a | |
| Dihydro α-ionone | 33.59 | 1786 | 1.57 ± 0.09 a | 0.96 ± 0.24 b | |
| Dihydro β-ionone | 34.05 | 1802 | 5.63 ± 0.86 a | 6.53 ± 1.33 a | |
| Monoterpenes | 2-Carene | 15.47 | 1264 | 0.07 ± 0.03 a | 0.38 ± 0.24 a |
| 2-β-Pinene | 6.19 | 1092 | nd | 0.54 ± 0.16 | |
| α-Phellanderene | 8.79 | 1124 | 0.01 ± 0.01 a | 15.33 ± 4.72 a | |
| α-Terpinene | 9.65 | 1133 | nd | 0.86 ± 0.43 | |
| β-Myrcene | 9.18 | 1128 | nd | 4.91 ± 1.90 | |
| β-Phellandrene | 11.32 | 1177 | nd | 9.65 ± 2.96 | |
| γ-Terpinene | 13.65 | 1229 | nd | 0.67 ± 0.37 | |
| Limonene | 10.85 | 1164 | 0.08 ± 0.01 b | 2.54 ± 0.79 a | |
| o-Cymene | 14.81 | 1251 | nd | 2.69 ± 0.90 | |
| Acids | Acetic acid | 22.28 | 1424 | 2.38 ± 0.62 a | 1.02 ± 0.22 a |
| Nonanoic acid | 42.45 | 2119 | 0.41 ± 0.18 a | 0.08 ± 0.08 a | |
| Octanoic acid | 39.94 | 2025 | 0.78 ± 0.15 | nd | |
| Alkanes | Tridecane | 17.38 | 1302 | 7.02 ± 3.55 a | 0.02 ± 0.02 a |
| Z-3-Heptene | 7.55 | 1131 | nd | 2.34 ± 0.58 | |
| E-3-Heptene | 7.77 | 1113 | nd | 1.89 ± 0.56 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangiorgio, D.; Cellini, A.; Spinelli, F.; Farneti, B.; Khomenko, I.; Muzzi, E.; Savioli, S.; Pastore, C.; Rodriguez-Estrada, M.T.; Donati, I. Does Organic Farming Increase Raspberry Quality, Aroma and Beneficial Bacterial Biodiversity? Microorganisms 2021, 9, 1617. https://doi.org/10.3390/microorganisms9081617
Sangiorgio D, Cellini A, Spinelli F, Farneti B, Khomenko I, Muzzi E, Savioli S, Pastore C, Rodriguez-Estrada MT, Donati I. Does Organic Farming Increase Raspberry Quality, Aroma and Beneficial Bacterial Biodiversity? Microorganisms. 2021; 9(8):1617. https://doi.org/10.3390/microorganisms9081617
Chicago/Turabian StyleSangiorgio, Daniela, Antonio Cellini, Francesco Spinelli, Brian Farneti, Iuliia Khomenko, Enrico Muzzi, Stefano Savioli, Chiara Pastore, María Teresa Rodriguez-Estrada, and Irene Donati. 2021. "Does Organic Farming Increase Raspberry Quality, Aroma and Beneficial Bacterial Biodiversity?" Microorganisms 9, no. 8: 1617. https://doi.org/10.3390/microorganisms9081617
APA StyleSangiorgio, D., Cellini, A., Spinelli, F., Farneti, B., Khomenko, I., Muzzi, E., Savioli, S., Pastore, C., Rodriguez-Estrada, M. T., & Donati, I. (2021). Does Organic Farming Increase Raspberry Quality, Aroma and Beneficial Bacterial Biodiversity? Microorganisms, 9(8), 1617. https://doi.org/10.3390/microorganisms9081617