
Gut Dysbiosis during COVID-19 and Potential Effect of Probiotics

 ,
, 
Abstract
:1. Introduction
2. The Interaction between Respiratory Tract Diseases and Gut Microbiota
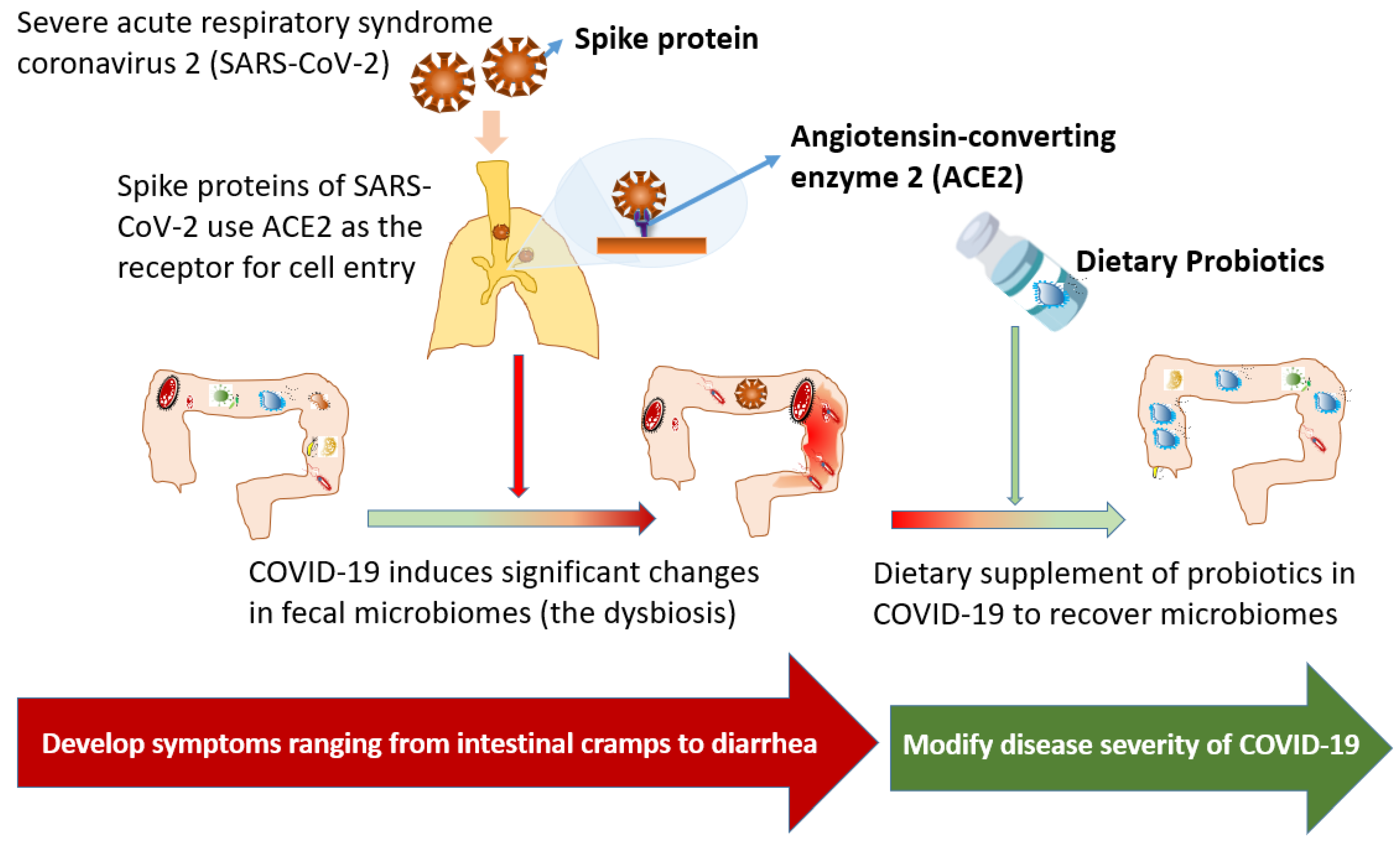
3. Gut Dysbiosis during COVID-19
4. Therapeutic Effects of Dietary Supplement of Probiotics for COVID-19
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lamers, M.M.; Beumer, J.; van der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.G.; van Schayck, J.P.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease In China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Suryana, K.D.; Simadibrata, M.; Renaldi, K. Impact of COVID-19 on the Gut: A review of the manifestations, pathology, management, and challenges. Acta Med. Indones. 2021, 53, 96–104. [Google Scholar]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B Betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Li, C.; Liu, X.; Chiu, M.C.; Zhao, X.; Wang, D.; Wei, Y.; Lee, A.; Zhang, A.J.; Chu, H.; et al. Infection of bat and human intestinal organoids by SARS-CoV-2. Nat. Med. 2020, 26, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef]
- de Oliveira, A.P.; Lopes, A.L.F.; Pacheco, G.; de Sa Guimaraes Noleto, I.R.; Nicolau, L.A.D.; Medeiros, J.V.R. Premises among SARS-CoV-2, dysbiosis and diarrhea: Walking through the ACE2/mTOR/autophagy route. Med. Hypotheses 2020, 144, 110243. [Google Scholar] [CrossRef] [PubMed]
- Bas, S.; Zarbaliyev, E. The role of dual-energy computed tomography in locating gastrointestinal tract perforations. Cureus 2021, 13, e15265. [Google Scholar]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical characteristics of 138 hospitalized patients with 2019 novel coronavirus-infected pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef]
- Monkemuller, K.; Fry, L.C.; Rickes, S. Systemic inflammatory response and thrombosis due to alterations in the gut microbiota in COVID-19. Rev. Esp. Enferm. Dig. 2020, 112, 584–585. [Google Scholar] [PubMed]
- Finlay, B.B.; Amato, K.R.; Azad, M.; Blaser, M.J.; Bosch, T.C.G.; Chu, H.; Dominguez-Bello, M.G.; Ehrlich, S.D.; Elinav, E.; Geva-Zatorsky, N.; et al. The hygiene hypothesis, the COVID pandemic, and consequences for the human microbiome. Proc. Natl. Acad. Sci. USA 2021, 118, e2010217118. [Google Scholar] [CrossRef]
- Groves, H.T.; Higham, S.L.; Moffatt, M.F.; Cox, M.J.; Tregoning, J.S. Respiratory viral infection alters the gut microbiota by inducing inappetence. Mbio 2020, 11, e03236-19. [Google Scholar] [CrossRef] [Green Version]
- Ichinohe, T.; Pang, I.K.; Kumamoto, Y.; Peaper, D.R.; Ho, J.H.; Murray, T.S.; Iwasaki, A. Microbiota regulates immune defense against respiratory tract influenza A virus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 5354–5359. [Google Scholar] [CrossRef] [Green Version]
- Enaud, R.; Prevel, R.; Ciarlo, E.; Beaufils, F.; Wieers, G.; Guery, B.; Delhaes, L. The gut-lung axis in health and respiratory diseases: A place for inter-organ and inter-kingdom crosstalks. Front. Cell Infect. Microbiol. 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [Green Version]
- Kuss, S.K.; Best, G.T.; Etheredge, C.A.; Pruijssers, A.J.; Frierson, J.M.; Hooper, L.V.; Dermody, T.S.; Pfeiffer, J.K. Intestinal microbiota promote enteric virus replication and systemic pathogenesis. Science 2011, 334, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Wilks, J.; Golovkina, T. Influence of microbiota on viral infections. PLoS Pathog. 2012, 8, e1002681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Hu, J.; Feng, J.W.; Hu, X.T.; Wang, T.; Gong, W.X.; Huang, K.; Guo, Y.X.; Zou, Z.; Lin, X.; et al. Influenza infection elicits an expansion of gut population of endogenous Bifidobacterium animalis which protects mice against infection. Genome Biol. 2020, 21, 99. [Google Scholar] [CrossRef] [PubMed]
- Negi, S.; Das, D.K.; Pahari, S.; Nadeem, S.; Agrewala, J.N. Potential role of gut microbiota in induction and regulation of innate immune memory. Front. Immunol. 2019, 10, 2441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, D.; Mohanty, A. Gut microbiota and Covid-19- possible link and implications. Virus Res. 2020, 285, 198018. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. USA 2010, 107, 12204–12209. [Google Scholar] [CrossRef] [Green Version]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in gut microbiota of patients with COVID-19 during time of hospitalization. Gastroenterology 2020, 159, 944–955.e8. [Google Scholar] [CrossRef]
- Tang, L.; Gu, S.; Gong, Y.; Li, B.; Lu, H.; Li, Q.; Zhang, R.; Gao, X.; Wu, Z.; Zhang, J.; et al. Clinical significance of the correlation between changes in the major intestinal bacteria species and COVID-19 severity. Eng. Beijing 2020, 6, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Liu, Q.; Zhang, F.; Lui, G.C.; Tso, E.Y.; Yeoh, Y.K.; Chen, Z.; Boon, S.S.; Chan, F.K.; Chan, P.K.; et al. Depicting SARS-CoV-2 faecal viral activity in association with gut microbiota composition in patients with COVID-19. Gut 2021, 70, 276–284. [Google Scholar] [CrossRef]
- Gu, S.; Chen, Y.; Wu, Z.; Chen, Y.; Gao, H.; Lv, L.; Guo, F.; Zhang, X.; Luo, R.; Huang, C.; et al. Alterations of the gut microbiota in patients with coronavirus disease 2019 or H1N1 influenza. Clin. Infect. Dis. 2020, 71, 2669–2678. [Google Scholar] [CrossRef]
- Lahti, L.; Salojarvi, J.; Salonen, A.; Scheffer, M.; de Vos, W.M. Tipping elements in the human intestinal ecosystem. Nat. Commun. 2014, 5, 4344. [Google Scholar] [CrossRef] [Green Version]
- McNabney, S.M.; Henagan, T.M. Short chain fatty acids in the colon and peripheral tissues: A focus on butyrate, colon cancer, obesity and insulin resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsome, R.C.; Gauthier, J.; Hernandez, M.C.; Abraham, G.E.; Robinson, T.O.; Williams, H.B.; Sloan, M.; Owings, A.; Laird, H.; Christian, T.; et al. The gut microbiome of COVID-19 recovered patients returns to uninfected status in a minority-dominated United States cohort. Gut Microbes 2021, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Cai, H.; Shen, Y.; Ni, Q.; Chen, Y.; Hu, S.; Li, J.; Wang, H.; Yu, L.; Huang, H.; et al. Management of corona virus disease-19 (COVID-19): The Zhejiang experience. Zhejiang Exp. J. Zhejiang Univ. Med. Sci. 2020, 49, 147–157. [Google Scholar]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Zuo, T.; Zhan, H.; Zhang, F.; Liu, Q.; Tso, E.Y.K.; Lui, G.C.Y.; Chen, N.; Li, A.; Lu, W.; Chan, F.K.L.; et al. Alterations in fecal fungal microbiome of patients with COVID-19 during time of hospitalization until discharge. Gastroenterology 2020, 159, 1302–1310. [Google Scholar] [CrossRef]
- Chen, Y.; Gu, S.; Chen, Y.; Lu, H.; Shi, D.; Guo, J.; Wu, W.R.; Yang, Y.; Li, Y.; Xu, K.J.; et al. Six-month follow-up of gut microbiota richness in patients with COVID-19. Gut 2021. [Google Scholar] [CrossRef]
- Yasui, H.; Kiyoshima, J.; Hori, T.; Shida, K. Protection against influenza virus infection of mice fed Bifidobacterium breve YIT4064. Clin. Diagn. Lab. Immunol. 1999, 6, 186–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Cao, J.; Wang, M.; Lu, M.; Chen, G.; Gao, F.; Liu, Z.; Zhang, D.; Ke, X.; Yi, M. Effects of Lactococcus lactis subsp. lactis JCM5805 on colonization dynamics of gut microbiota and regulation of immunity in early ontogenetic stages of tilapia. Fish Shellfish Immunol. 2019, 86, 53–63. [Google Scholar] [CrossRef]
- Su, M.; Jia, Y.; Li, Y.; Zhou, D.; Jia, J. Probiotics for the prevention of ventilator-associated pneumonia: A meta-analysis of randomized controlled trials. Respir. Care 2020, 65, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 cytokine storm; what we know so far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Britton, G.J.; Chen-Liaw, A.; Cossarini, F.; Livanos, A.E.; Spindler, M.P.; Plitt, T.; Eggers, J.; Mogno, I.; Gonzalez-Reiche, A.; Siu, S.; et al. SARS-CoV-2-specific IgA and limited inflammatory cytokines are present in the stool of select patients with acute COVID-19. MedRxiv 2020. [Google Scholar] [CrossRef]
- Han, S.K.; Shin, Y.J.; Lee, D.Y.; Kim, K.M.; Yang, S.J.; Kim, D.S.; Choi, J.W.; Lee, S.; Kim, D.H. Lactobacillus rhamnosus HDB1258 modulates gut microbiota-mediated immune response in mice with or without lipopolysaccharide-induced systemic inflammation. BMC Microbiol. 2021, 21, 146. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Hober, D.; Hamze, M.; Chihib, N.E.; Drider, D. Antiviral potential of lactic acid bacteria and their bacteriocins. Probiotics Antimicrob. Proteins 2014, 6, 177–185. [Google Scholar] [CrossRef]
- Baindara, P.; Chakraborty, R.; Holliday, Z.M.; Mandal, S.M.; Schrum, A.G. Oral probiotics in coronavirus disease 2019: Connecting the gut-lung axis to viral pathogenesis, inflammation, secondary infection and clinical trials. New Microbes New Infect. 2021, 40, 100837. [Google Scholar] [CrossRef] [PubMed]
- Starosila, D.; Rybalko, S.; Varbanetz, L.; Ivanskaya, N.; Sorokulova, I. Anti-influenza activity of a Bacillus subtilis probiotic strain. Antimicrob. Agents Chemother. 2017, 61, e00539-17. [Google Scholar] [CrossRef] [Green Version]
- Malaczewska, J.; Kaczorek-Lukowska, E.; Wojcik, R.; Siwicki, A.K. Antiviral effects of nisin, lysozyme, lactoferrin and their mixtures against bovine viral diarrhoea virus. BMC Vet. Res. 2019, 15, 318. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Wang, C.T.; Zhang, F.S.; Qi, F.; Wang, S.F.; Ma, S.; Wu, T.J.; Tian, H.; Tian, Z.T.; Zhang, S.L.; et al. Effect of probiotics on the incidence of ventilator-associated pneumonia in critically ill patients: A randomized controlled multicenter trial. Intensive Care Med. 2016, 42, 1018–1028. [Google Scholar] [CrossRef]
- Morrow, L.E.; Kollef, M.H.; Casale, T.B. Probiotic prophylaxis of ventilator-associated pneumonia: A blinded, randomized, controlled trial. Am. J. Respir. Crit. Care Med. 2010, 182, 1058–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, Y.J.; Lee, Y.T.; Ngo, V.L.; Cho, Y.H.; Ko, E.J.; Hong, S.M.; Kim, K.H.; Jang, J.H.; Oh, J.S.; Park, M.K.; et al. Heat-killed Lactobacillus casei confers broad protection against influenza A virus primary infection and develops heterosubtypic immunity against future secondary infection. Sci. Rep. 2017, 7, 17360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidossi, A.; De Grandi, R.; Toscano, M.; Bottagisio, M.; De Vecchi, E.; Gelardi, M.; Drago, L. Probiotics Streptococcus salivarius 24SMB and Streptococcus oralis 89a interfere with biofilm formation of pathogens of the upper respiratory tract. BMC Infect. Dis. 2018, 18, 653. [Google Scholar] [CrossRef] [PubMed]
- Di Pierro, F. A possible probiotic (S. salivarius K12) approach to improve oral and lung microbiotas and raise defenses against SAR S-CoV-2. Minerva Med. 2020, 111, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Bottari, B.; Castellone, V.; Neviani, E. Probiotics and COVID-19. Int. J. Food Sci. Nutr. 2021, 72, 293–299. [Google Scholar] [CrossRef]
- Kumar, R.; Seo, B.J.; Mun, M.R.; Kim, C.J.; Lee, I.; Kim, H.; Park, Y.H. Putative probiotic Lactobacillus spp. from porcine gastrointestinal tract inhibit transmissible gastroenteritis coronavirus and enteric bacterial pathogens. Trop. Anim. Health Prod. 2010, 42, 1855–1860. [Google Scholar]
- Chai, W.; Burwinkel, M.; Wang, Z.; Palissa, C.; Esch, B.; Twardziok, S.; Rieger, J.; Wrede, P.; Schmidt, M.F. Antiviral effects of a probiotic Enterococcus faecium strain against transmissible gastroenteritis coronavirus. Arch. Virol. 2013, 158, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.S.; Liu, Q.; Jiang, Y.L.; Yang, W.T.; Huang, H.B.; Shi, C.W.; Yang, G.L.; Wang, C.F. Surface-displayed porcine IFN-lambda3 in Lactobacillus plantarum inhibits porcine enteric coronavirus infection of porcine intestinal epithelial cells. J. Microbiol. Biotechnol. 2020, 30, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Mak, J.W.Y.; Chan, F.K.L.; Ng, S.C. Probiotics and COVID-19: One size does not fit all. Lancet Gastroenterol. Hepatol. 2020, 5, 644–645. [Google Scholar] [CrossRef]
- Kurian, S.J.; Unnikrishnan, M.K.; Miraj, S.S.; Bagchi, D.; Banerjee, M.; Reddy, B.S.; Rodrigues, G.S.; Manu, M.K.; Saravu, K.; Mukhopadhyay, C.; et al. Probiotics in prevention and treatment of COVID-19: Current perspective and future prospects. Arch. Med. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zhang, M.; Yao, G.; Kwok, L.Y.; Zhang, W. Probiotics as adjunctive treatment for patients contracted COVID-19: Current understanding and future needs. Front. Nutr. 2021, 8, 669808. [Google Scholar] [CrossRef]
- Sundararaman, A.; Ray, M.; Ravindra, P.V.; Halami, P.M. Role of probiotics to combat viral infections with emphasis on COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 8089–8104. [Google Scholar] [CrossRef] [PubMed]
- Khaled, J.M.A. Probiotics, prebiotics, and COVID-19 infection: A review article. Saudi J. Biol. Sci. 2021, 28, 865–869. [Google Scholar] [CrossRef]
- Esaiassen, E.; Cavanagh, P.; Hjerde, E.; Simonsen, G.S.; Stoen, R.; Klingenberg, C. Bifidobacterium longum subspecies infantis bacteremia in 3 extremely preterm infants receiving probiotics. Emerg. Infect. Dis. 2016, 22, 1664–1666. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, C.; Pillonel, T.; Torregrossa, A.; Prod’hom, G.; Fischer, C.J.; Greub, G.; Giannoni, E. Bifidobacterium longum bacteremia in preterm infants receiving probiotics. Clin. Infect. Dis. 2015, 60, 924–927. [Google Scholar] [CrossRef] [Green Version]
- Alataby, H.; Atemnkeng, F.; Bains, S.S.; Kenne, F.M.; Diaz, K.; Nfonoyim, J. A COVID-19 case complicated by Candida dubliniensis and Klebsiella pneumoniae-carbapenem-resistant Enterobacteriaceae. J. Med. Cases 2020, 11, 403–406. [Google Scholar] [CrossRef]
- Miao, Q.; Ma, Y.; Ling, Y.; Jin, W.; Su, Y.; Wang, Q.; Pan, J.; Zhang, Y.; Chen, H.; Yuan, J.; et al. Evaluation of superinfection, antimicrobial usage, and airway microbiome with metagenomic sequencing in COVID-19 patients: A cohort study in Shanghai. J. Microbiol. Immunol. Infect. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, G.; Borrazzo, C.; Pinacchio, C.; Santinelli, L.; Innocenti, G.P.; Cavallari, E.N.; Celani, L.; Marazzato, M.; Alessandri, F.; Ruberto, F.; et al. Oral bacteriotherapy in patients with COVID-19: A retrospective cohort study. Front. Nutr. 2020, 7, 613928. [Google Scholar] [CrossRef]
- d’Ettorre, G.; Ceccarelli, G.; Marazzato, M.; Campagna, G.; Pinacchio, C.; Alessandri, F.; Ruberto, F.; Rossi, G.; Celani, L.; Scagnolari, C.; et al. Challenges in the management of SARS-CoV2 Infection: The role of oral bacteriotherapy as complementary therapeutic strategy to avoid the progression of COVID-19. Front. Med. 2020, 7, 389. [Google Scholar] [CrossRef] [PubMed]
- Clinicaltrials.gov. Available online: https://clinicaltrials.gov/ct2/home (accessed on 24 July 2021).
- Gautier, T.; Gall, S.D.L.; Sweidan, A.; Tamanai-Shacoori, Z.; Jolivet-Gougeon, A.; Loréal, O.; Bousarghin, L. Next-generation probiotics and their metabolites in COVID-19. Microorganisms 2021, 9, 941. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| ClinicalTrials.gov Identifier | Study Title | First Posted | Study Design | Probiotic Strain | Location | Outcome Measures | Status |
|---|---|---|---|---|---|---|---|
| NCT04366180 | Evaluation of probiotic Lactobacillus coryniformis K8 on COVID-19 prevention in healthcare workers | 28 April 2020 | Randomized | L. coryniformis K8 | Granada, Spain | Incidence of COVID-19 infection in healthcare workers | Recruiting |
| NCT04390477 | Study to evaluate the effect of a probiotic in COVID-19 | 15 May 2020 | Randomized | Not revealed | Alicante, Spain | ICU admission rate | Recruiting |
| NCT04399252 | Effect of Lactobacillus on the microbiome of household contacts exposed to COVID-19 | 22 May 2020 | Randomized | L. rhamnosus GG | North Carolina, United States | Incidence of symptoms of COVID-19 | Active, not recruiting |
| NCT04420676 | Synbiotic therapy of gastrointestinal symptoms during COVID-19 infection (SynCov) | 9 June 2020 | Randomized | Omni-Biotic® 10 AAD (chiefly Lactobacillus and Bifidobacterium) | Graz, Austria | Stool calprotectin | Recruiting |
| NCT04462627 | Reduction of COVID 19 transmission to health care professionals | 8 July 2020 | Non-randomized | Metagenics Probactiol plus (chiefly Lactobacillus and Bifidobacterium) | Brussels, Belgium | Antibody concentration | Recruiting |
| NCT04507867 | Effect of a NSS to reduce complications in patients with COVID-19 and comorbidities in stage III | 11 August 2020 | Randomized | Saccharomyces bourllardii with nutritional support system (NSS) | Mexico | Oxygen saturation | Not yet recruiting |
| NCT04517422 | Efficacy of L. plantarum and P. acidilactici in adults with SARS-CoV-2 and COVID-19 | 18 August 2020 | RCT | L. plantarum and P. acidilactici | Mexico City, Mexico | Severity progression of COVID-19 | Completed |
| NCT04621071 | Efficacy of probiotics in reducing duration and symptoms of COVID-19 (PROVID-19) | 9 November 2020 | RCT | Not revealed | Canada, Quebec | Duration of symptoms of the COVID-19 | Recruiting |
| NCT04666116 | Changes in viral load in COVID-19 after probiotics | 14 December 2020 | Randomized, single blind | GASTEEL PLUS (mixture of Bifidobacteria and Lactobacillus) | Valencia, Spain | Viral load in nasopharyngeal smear | Recruiting |
| NCT04734886 | The effect of probiotic supplementation on SARS-CoV-2 antibody response after COVID-19 | 2 February 2021 | Randomized | L. reuteri DSM 17938 + vitamin D | Örebro Län, Sweden | SARS-CoV-2 specific antibodies | Recruiting |
| NCT04756466 | Effect of the consumption of a Lactobacillus strain on the incidence of COVID-19 in the elderly | 16 February 2021 | RCT | Lactobacillus strain | A Coruña, Spain | Incidence of SARS CoV-2 infection | Active, not recruiting |
| NCT04798677 | Efficacy and tolerability of ABBC1 in volunteers receiving the influenza or COVID-19 Vaccine | 15 March 2021 | Non-randomized | S. cerevisiae, rich in selenium and zinc | Barcelona, Spain | Change in acute immune response to influenza vaccine after supplementation | Recruiting |
| NCT04813718 | Post COVID-19 syndrome and the gut-lung axis | 24 March 2021 | Randomized | Omni-Biotic Pro Vi 5 (chiefly Lactobacillus) | Graz, Austria | Microbiome composition | Recruiting |
| NCT04847349 | Live microbials to boost anti-severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) immunity clinical trial | 19 April 2021 | RCT | OL-1 (Content not revealed) | New Jersey, United States | Change in serum titer of anti-SARS-CoV-2 IgG | Recruiting |
| NCT04854941 | Efficacy of probiotics in the treatment of hospitalized patients with novel coronavirus infection | 22 April 2021 | Randomized | L. rhamnosus, B. bifidum, B. longum subsp. infantis and B. longum | Moscow, Russian | Mortality | Completed |
| NCT04877704 | Symprove (Probiotic) as an add-on to COVID-19 management | 7 May 2021 | Randomized | Symprove ( L. rhamnosus, E. faecium, L. acidophilus and L. plantarum) | London, United Kingdom | Length of hospital stay | Not yet recruiting |
| NCT04884776 | Modulation of gut microbiota to enhance health and immunity | 13 May 2021 | RCT | Probiotics blend (3 Bifidobacteria) | Hong Kong | Restoration of gut dysbiosis | Not yet recruiting |
| NCT04907877 | Bifidobacteria and Lactobacillus in symptomatic adult COVID-19 outpatients (ProCOVID) | 1 June 2021 | Randomized | NordBiotic ImmunoVir (mixture of Bifidobacteria and Lactobacillus) | Not revealed | Global symptom score | Not yet recruiting |
| NCT04922918 | Ligilactobacillus salivarius MP101 for elderly in a nursing home (PROBELDERLY) | 11 June 2021 | Single group | Ligilactobacillus salivarius MP101 | Madrid, Spain | Barthel index, functional status score | Recruiting |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, Y.-P.; Lee, C.-C.; Lee, J.-C.; Tsai, P.-J.; Ko, W.-C. Gut Dysbiosis during COVID-19 and Potential Effect of Probiotics. Microorganisms 2021, 9, 1605. https://doi.org/10.3390/microorganisms9081605
Hung Y-P, Lee C-C, Lee J-C, Tsai P-J, Ko W-C. Gut Dysbiosis during COVID-19 and Potential Effect of Probiotics. Microorganisms. 2021; 9(8):1605. https://doi.org/10.3390/microorganisms9081605
Chicago/Turabian StyleHung, Yuan-Pin, Ching-Chi Lee, Jen-Chieh Lee, Pei-Jane Tsai, and Wen-Chien Ko. 2021. "Gut Dysbiosis during COVID-19 and Potential Effect of Probiotics" Microorganisms 9, no. 8: 1605. https://doi.org/10.3390/microorganisms9081605
APA StyleHung, Y.-P., Lee, C.-C., Lee, J.-C., Tsai, P.-J., & Ko, W.-C. (2021). Gut Dysbiosis during COVID-19 and Potential Effect of Probiotics. Microorganisms, 9(8), 1605. https://doi.org/10.3390/microorganisms9081605