
Organic Amendments Alter Soil Hydrology and Belowground Microbiome of Tomato (Solanum lycopersicum)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Growth Conditions
2.3. Organic Amendments
2.4. Growing Media
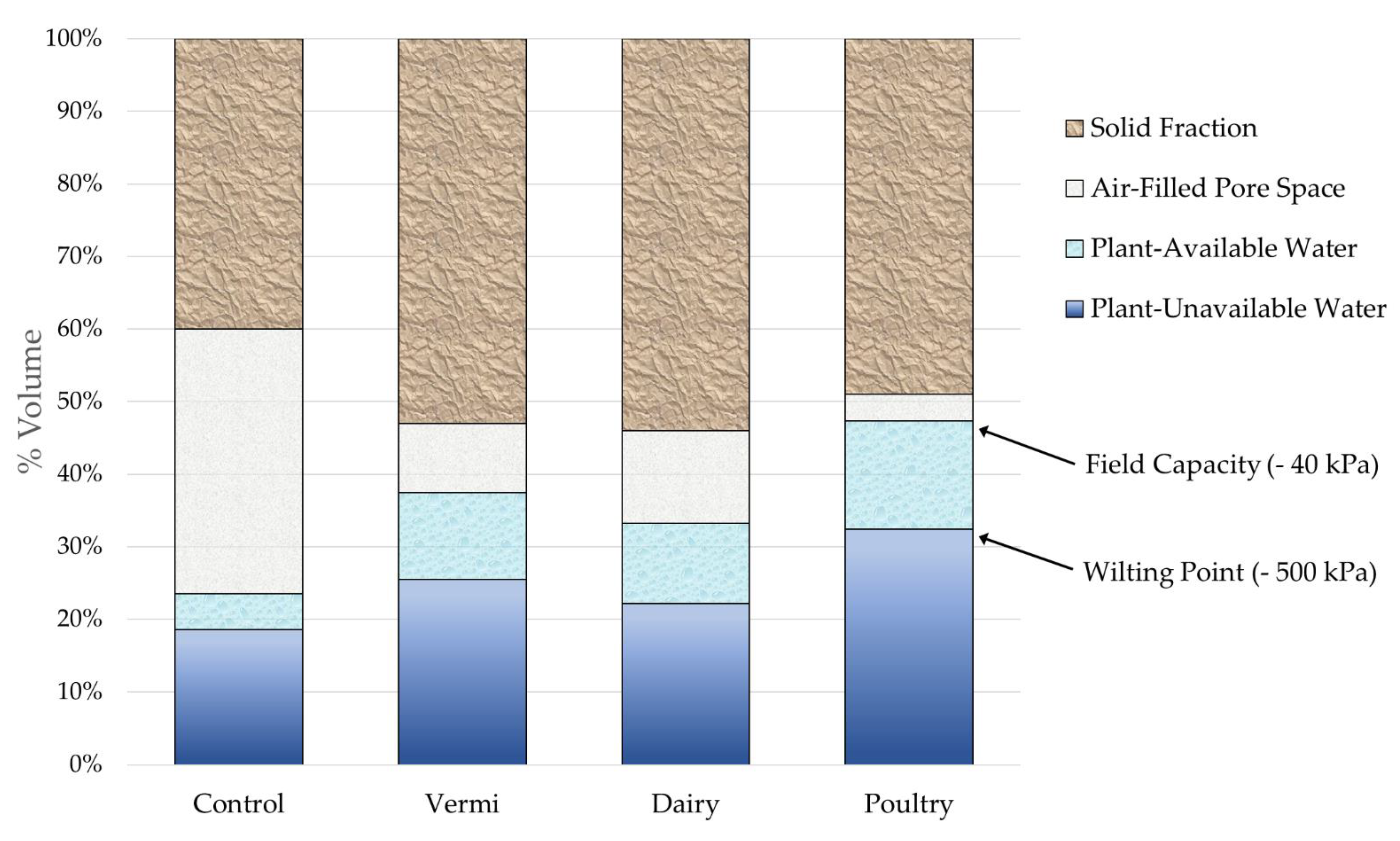
2.5. Soil Physical Properties
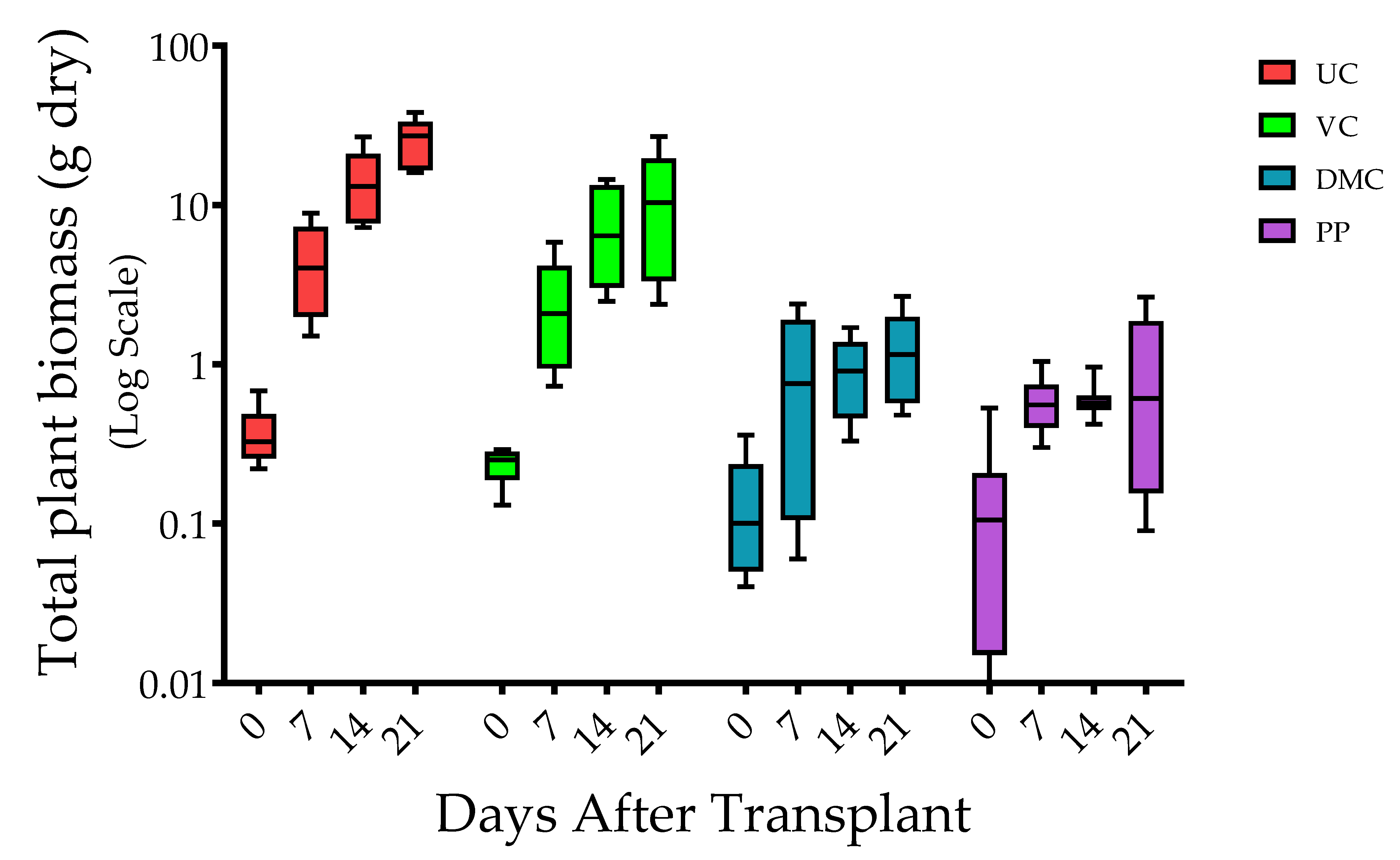
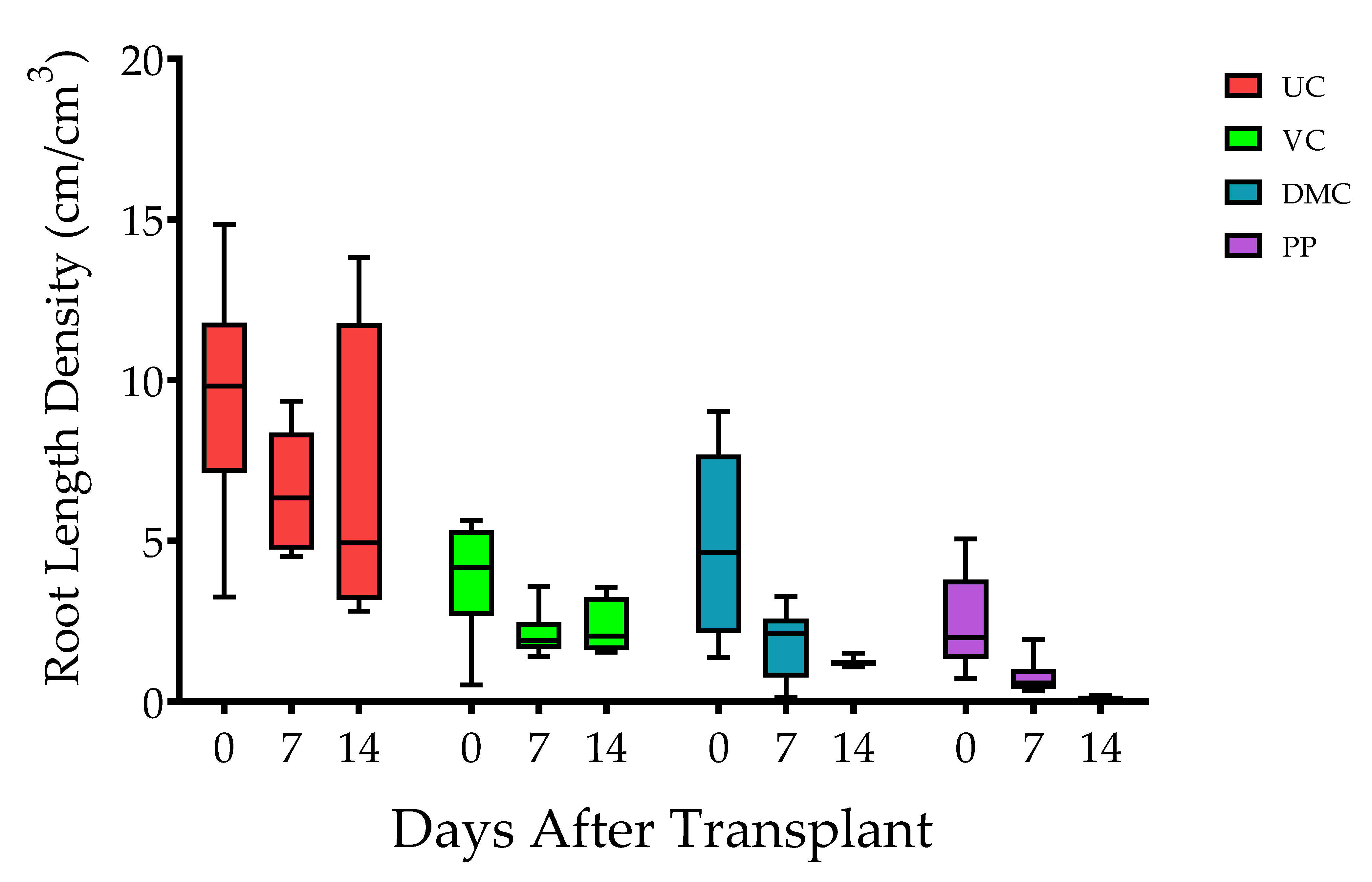
2.6. Plant Growth
2.7. Root Microbiome
2.8. PMA Photolysis and DNA Extraction
2.9. Amplicon Sequencing
2.10. Statistical Analysis
3. Results
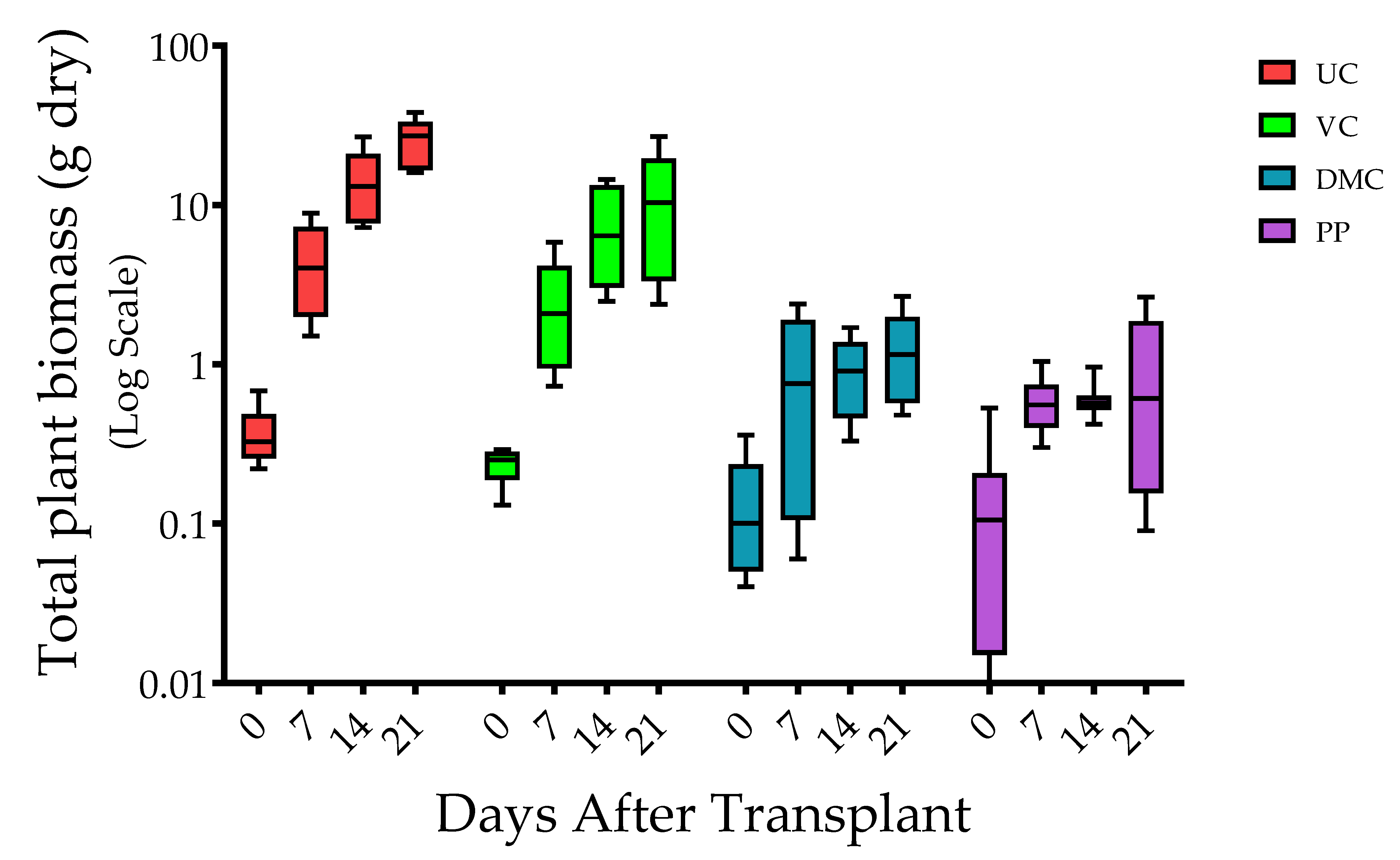
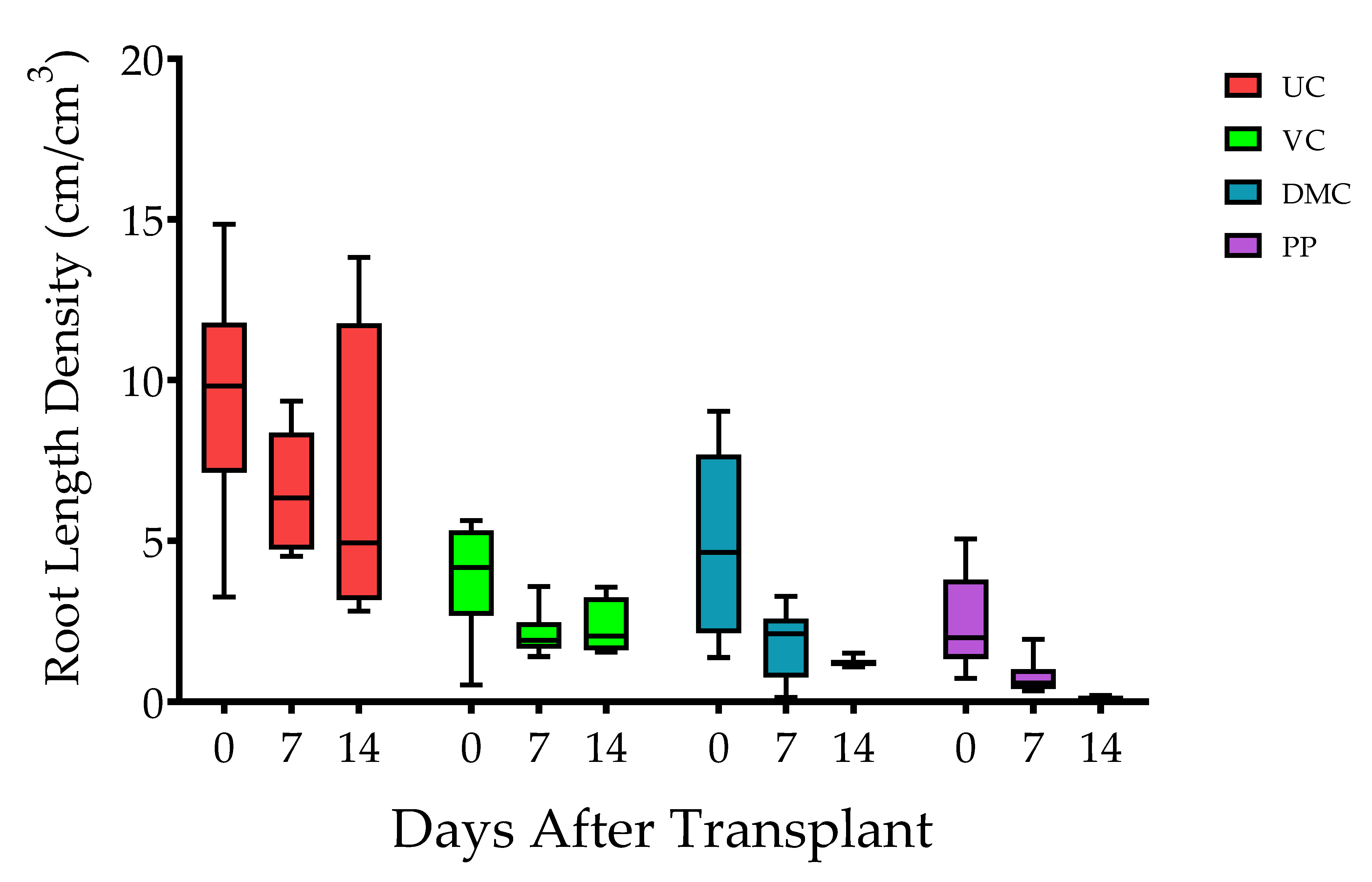
3.1. Plant Growth
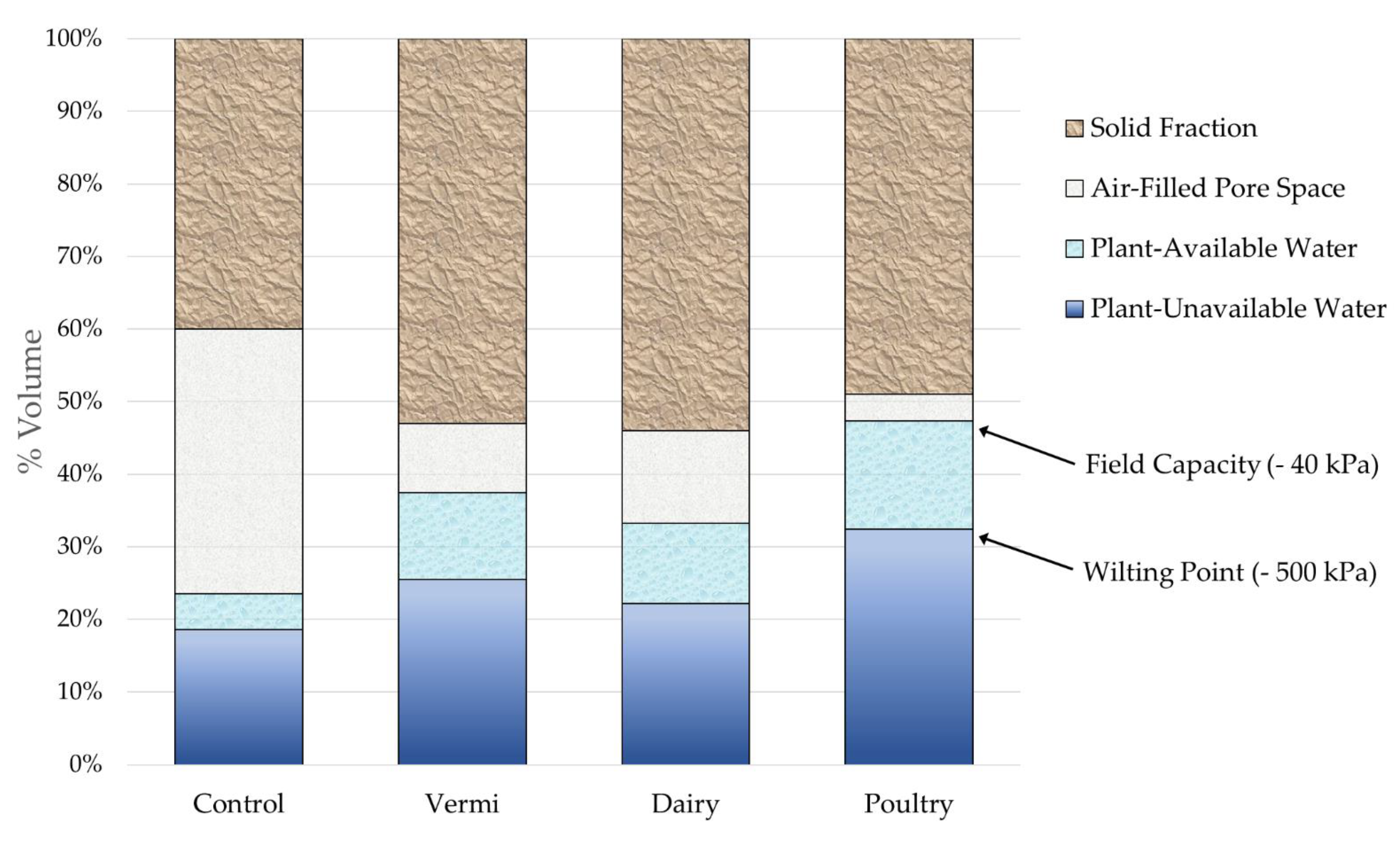
3.2. Soil Physical and Hydrological Properties
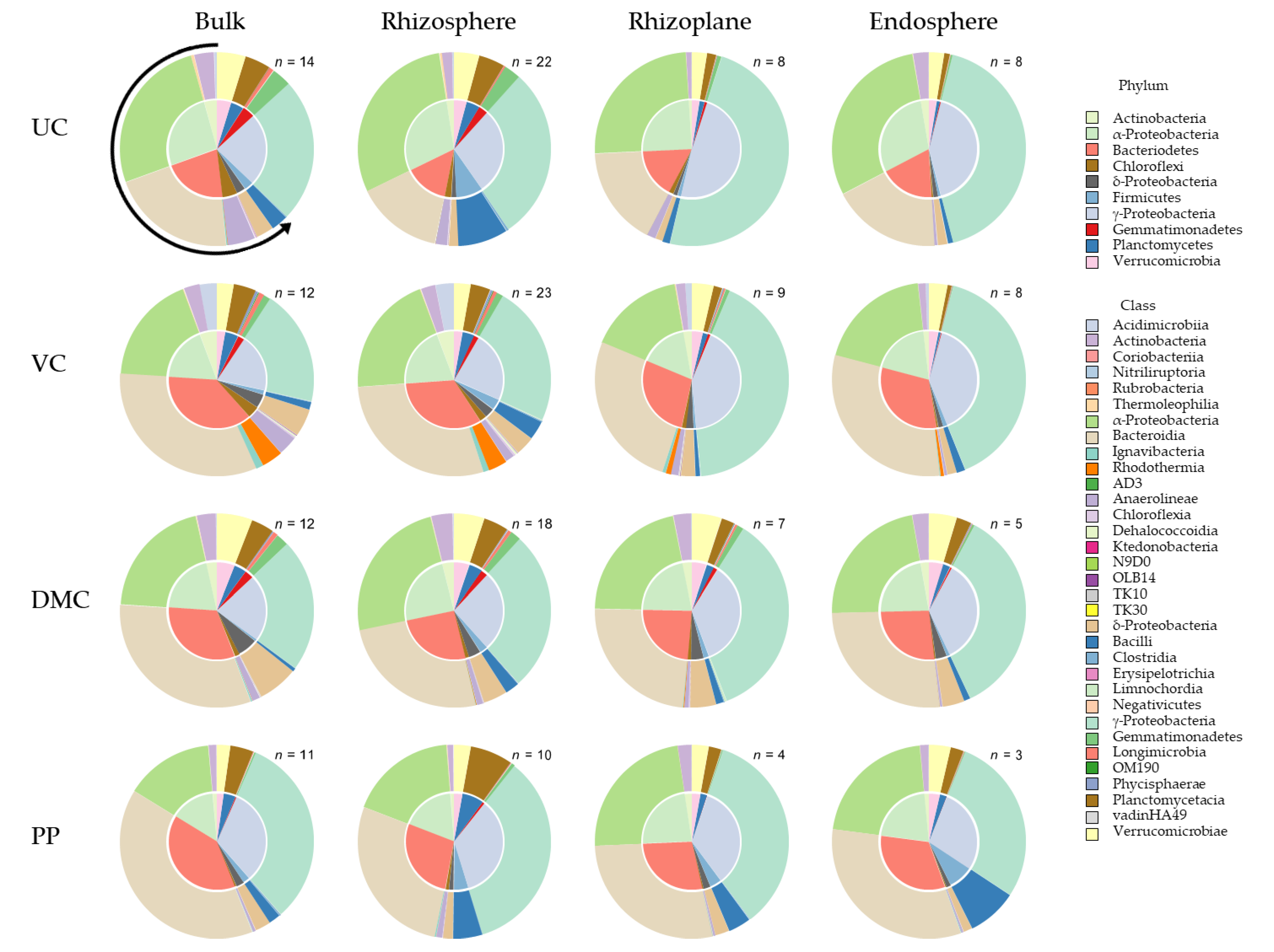
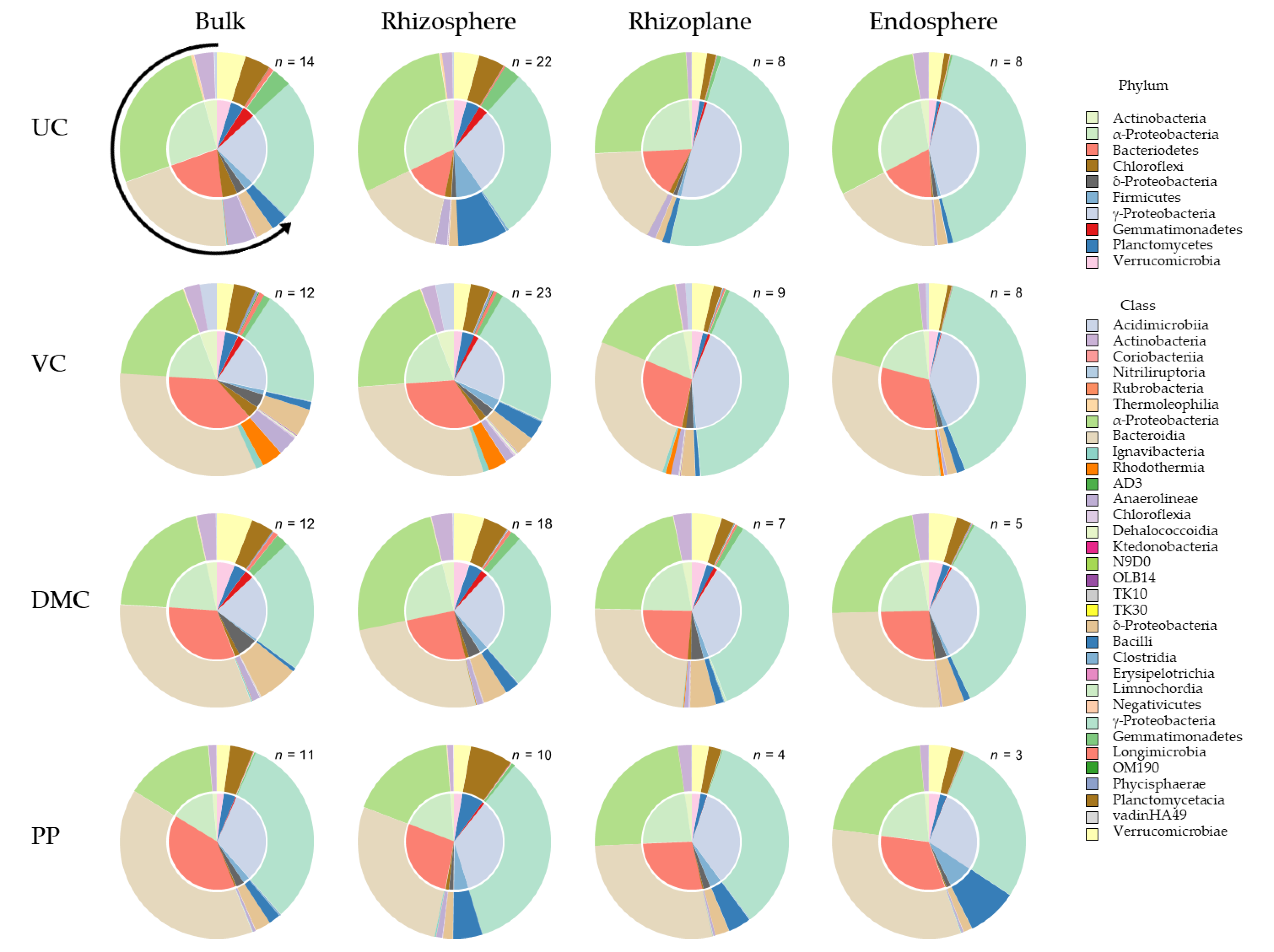
3.3. 16S Community Composition
3.3.1. Plant Phenology
3.3.2. Root Microhabitat
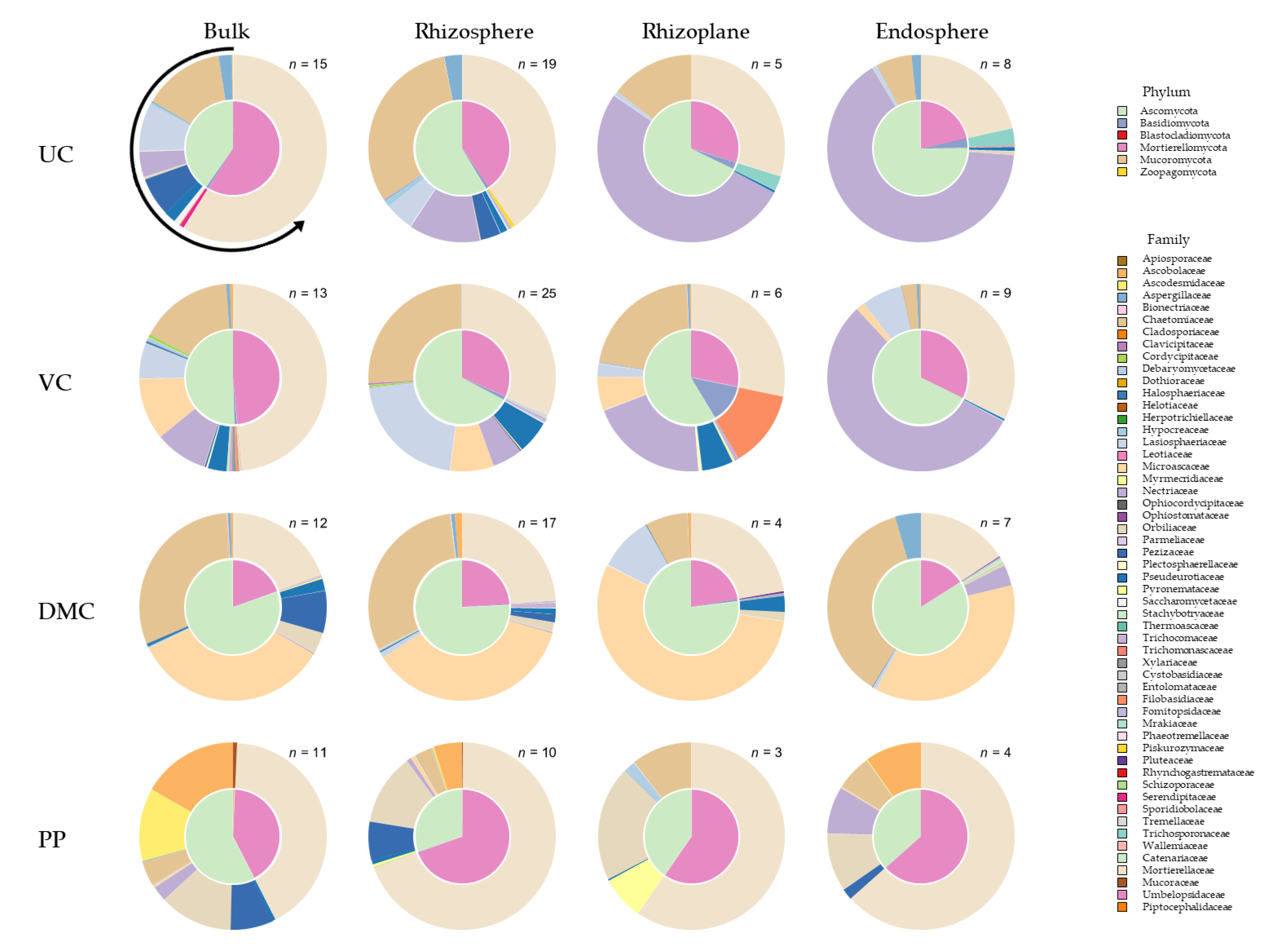
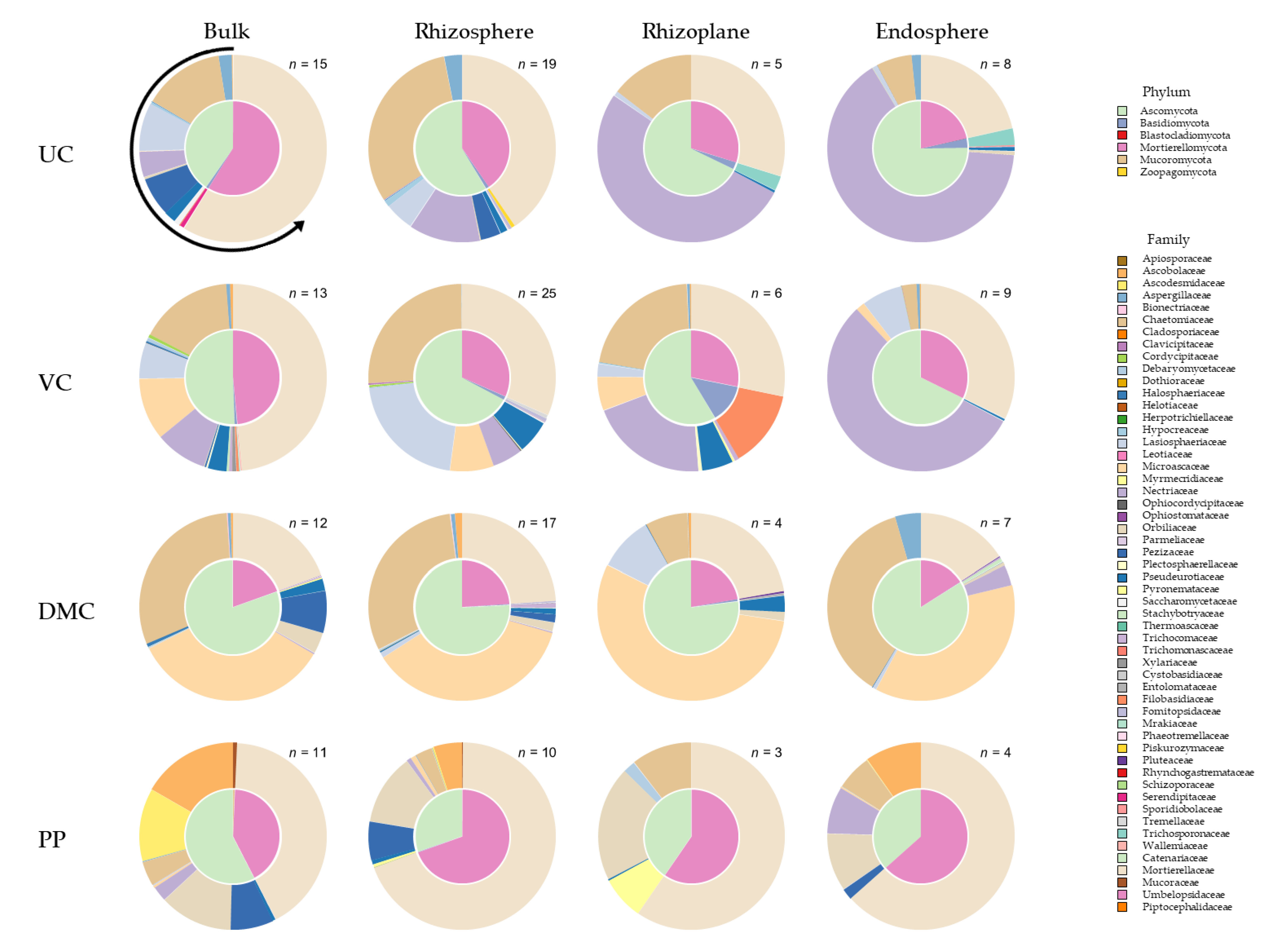
3.4. Fungal Community Composition
4. Discussion
4.1. Soil Properties and Plant Growth
4.2. Plant Phenology and Root Microhabitat
4.3. Treatment-Specific Microbial Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, S. Greenhouse gas accounting for landfill diversion of food scraps and yard waste. Compost Sci. Util. 2016, 24, 11–19. [Google Scholar] [CrossRef]
- Astill, G.; Minor, T.; Thornsbury, S. Changes in U.S. produce grower food safety practices from 1999 to 2016. Food Control 2019, 104, 326–332. [Google Scholar] [CrossRef]
- Celestina, C.; Hunt, J.R.; Sale, P.W.G.; Franks, A.E. Attribution of crop yield responses to application of organic amendments: A critical review. Soil Tillage Res. 2019, 186, 135–145. [Google Scholar] [CrossRef]
- Cesarano, G.; De Filippis, F.; La Storia, A.; Scala, F.; Bonanomi, G. Organic amendment type and application frequency affect crop yields, soil fertility and microbiome composition. Appl. Soil Ecol. 2017, 120, 254–264. [Google Scholar] [CrossRef]
- Bonanomi, G.; Lorito, M.; Vinale, F.; Woo, S.L. Organic amendments, beneficial microbes, and soil microbiota: Toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 2018, 56, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Aggelides, S.M.; Londra, P.A. Effects of compost produced from town wastes and sewage sludge on the physical properties of a loamy and a clay soil. Bioresour. Technol. 2000, 71, 253–259. [Google Scholar] [CrossRef]
- US Composting Council. Compost Definition. Available online: https://www.compostingcouncil.org/page/CompostDefinition (accessed on 4 June 2021).
- Neher, D.A.; Weicht, T.R.; Bates, S.T.; Leff, J.W.; Fierer, N. Changes in bacterial and fungal communities across compost recipes, preparation methods, and composting times. PLoS ONE 2013, 8, e79512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittenhouse, T. Tipsheet: Compost. Available online: https://attra.ncat.org/product/tipsheet-compost/ (accessed on 4 June 2021).
- Dominguez, J.; Edwards, C.A.; Subler, S. A comparison of vermicomposting and composting. BioCycle 1997, 38, 57–59. [Google Scholar]
- US Department of Agriculture. Processed Animal Manures in Organic Crop Production; NOP 5006; USDA: Washington, DC, USA, 2011.
- Kyakuwaire, M.; Olupot, G.; Amoding, A.; Nkedi-Kizza, P.; Basamba, T.A. How safe is chicken litter for land application as an organic fertilizer? A review. Int. J. Environ. Res. Public Health 2019, 16, 3521. [Google Scholar] [CrossRef] [Green Version]
- Obi, M.E.; Ebo, P.O. The effects of organic and inorganic amendments on soil physical properties and maize production in a severely degraded sandy soil in southern Nigeria. Bioresour. Technol. 1995, 51, 117–123. [Google Scholar] [CrossRef]
- Linn, D.M.; Doran, J.W. Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and nontilled soils. Soil Sci. Soc. Am. J. 1984, 48, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Pagliai, M.; Vittori Antisari, L. Influence of waste organic matter on soil micro-and macrostructure. Bioresour. Technol. 1993, 43, 205–213. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Vo, D.; Graham, K.J.; Scow, K.M. Soil water content and organic carbon availability are major determinants of soil microbial community composition. Microb. Ecol. 2004, 48, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Saison, C.; Degrange, V.; Oliver, R.; Millard, P.; Commeaux, C.; Montange, D.; Le Roux, X. Alteration and resilience of the soil microbial community following compost amendment: Effects of compost level and compost-borne microbial community. Environ. Microbiol. 2006, 8, 247–257. [Google Scholar] [CrossRef]
- Inbar, E.; Green, S.J.; Hadar, Y.; Minz, D. Competing factors of compost concentration and proximity to root affect the distribution of Streptomycetes. Microb. Ecol. 2005, 50, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Van Wees, S.C.M.; Van der Ent, S.; Pieterse, C.M.J. Plant immune responses triggered by beneficial microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2008, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.E.; Barea, J.-M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Glick, B. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbon, E.H.; Liberman, L.M. Beneficial microbes affect endogenous mechanisms controlling root development. Trends Plant Sci. 2016, 21, 218–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marasco, R.; Rolli, E.; Ettoumi, B.; Vigani, G.; Mapelli, F.; Borin, S.; Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A. A drought resistance-promoting microbiome is selected by root system under desert farming. PLoS ONE 2012, 7, e48479. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Pangesti, N.; Pineda, A.; Pieterse, C.M.; Dicke, M.; van Loon, J.J. Two-way plant mediated interactions between root-associated microbes and insects: From ecology to mechanisms. Front. Plant Sci. 2013, 4, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhold-Hurek, B.; Bunger, W.; Burbano, C.S.; Sabale, M.; Hurek, T. Roots shaping their microbiome: Global hotspots for microbial activity. Annu. Rev. Phytopathol. 2015, 53, 403–424. [Google Scholar] [CrossRef]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Allard, S.M.; Walsh, C.S.; Wallis, A.E.; Ottesen, A.R.; Brown, E.W.; Micallef, S.A. Solanum lycopersicum (tomato) hosts robust phyllosphere and rhizosphere bacterial communities when grown in soil amended with various organic and synthetic fertilizers. Sci. Total Environ. 2016, 573, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, A.; Satpute, A.; Albrecht, U.; Strauss, S.L. Impact of soil microbial amendments on tomato rhizosphere microbiome and plant growth in field soil. Microb. Ecol. 2020, 80, 398–409. [Google Scholar] [CrossRef]
- Neher, D.A.; Cutler, A.J.; Weicht, T.R.; Sharma, M.; Millner, P.D. Composts of poultry litter or dairy manure differentially affect survival of enteric bacteria in fields with spinach. J. Appl. Microbiol. 2019, 126, 1910–1922. [Google Scholar] [CrossRef]
- Neher, D.A.; Limoges, M.A.; Weicht, T.R.; Sharma, M.; Millner, P.D.; Donnelly, C. Bacterial community dynamics distinguish poultry compost from dairy compost and non-amended soils planted with spinach. Microorganisms 2020, 8, 1601. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; Alioto, D.; Minutolo, M.; Marra, R.; Cesarano, G.; Vinale, F. Organic amendments modulate soil microbiota and reduce virus disease incidence in the TSWV-tomato pathosystem. Pathogens 2020, 9, 379. [Google Scholar] [CrossRef] [PubMed]
- Jack, A.L.H.; Rangarajan, A.; Culman, S.W.; Sooksa-Nguan, T.; Thies, J.E. Choice of organic amendments in tomato transplants has lasting effects on bacterial rhizosphere communities and crop performance in the field. Appl. Soil Ecol. 2011, 48, 94–101. [Google Scholar] [CrossRef]
- Neher, D.A.; Fang, L.; Weicht, T.R. Ecoenzymes as indicators of compost to suppress Rhizoctonia solani. Compost Sci. Util. 2017, 25, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Blouin, M.; Barrere, J.; Meyer, N.; Lartigue, S.; Barot, S.; Mathieu, J. Vermicompost significantly affects plant growth: A meta-analysis. Agron. Sustain. Dev. 2019, 39, 34. [Google Scholar] [CrossRef]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2016, 2, 16242. [Google Scholar] [CrossRef]
- Fittipaldi, M.; Nocker, A.; Codony, F. Progress in understanding preferential detection of live cells using viability dyes in combination with DNA amplification. J. Microbiol. Methods 2012, 91, 276–289. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazcano, C.; Gomez-Brandon, M.; Dominguez, J. Comparison of the effectiveness of composting and vermicomposting for the biological stabilization of cattle manure. Chemosphere 2008, 72, 1013–1019. [Google Scholar] [CrossRef]
- Lim, S.L.; Wu, T.Y.; Lim, P.N.; Shak, K.P. The use of vermicompost in organic farming: Overview, effects on soil and economics. J. Sci. Food Agric. 2015, 95, 1143–1156. [Google Scholar] [CrossRef]
- Wichuk, K.M.; McCartney, D. Compost stability and maturity evaluation—A literature review. Can. J. Civil Eng. 2010, 37, 1505–1523. [Google Scholar] [CrossRef]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Yu, X.; Liang, X.; Liu, Y.; Borriss, R.; Liu, Y. Addition of plant-growth-promoting Bacillus subtilis PTS-394 on tomato rhizosphere has no durable impact on composition of root microbiome. BMC Microbiol. 2017, 17, 131. [Google Scholar] [CrossRef]
- Kwak, M.J.; Kong, H.G.; Choi, K.; Kwon, S.K.; Song, J.Y.; Lee, J.; Lee, P.A.; Choi, S.Y.; Seo, M.; Lee, H.J.; et al. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat. Biotechnol. 2018, 36, 1117. [Google Scholar] [CrossRef]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Lee, S.A.; Kim, Y.; Kim, J.M.; Chu, B.; Joa, J.H.; Sang, M.K.; Song, J.; Weon, H.Y. A preliminary examination of bacterial, archaeal, and fungal communities inhabiting different rhizocompartments of tomato plants under real-world environments. Sci. Rep. 2019, 9, 9300. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Lei, S.; Li, Y.; Huang, W.; Ma, R.; Xiong, J.; Zhang, T.; Jin, L.; Haq, H.U.; Xu, X.; et al. Revealing the variation and stability of bacterial communities in tomato rhizosphere microbiota. Microorganisms 2020, 8, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryckeboer, J.; Mergaert, J.; Vaes, K.; Klammer, S.; De Clercq, D.; Coosemans, J.; Insam, H.; Swings, J. A survey of bacteria and fungi occurring during composting and self-heating processes. Ann. Microbiol. 2003, 53, 349–410. [Google Scholar]
- Drew, M.C. Oxygen deficiency and root metabolism: Injury and acclimation under hypoxia and anoxia. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 223–250. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.S.; Taylor, B.R. Effects of pulp mill solids and three composts on early growth of tomatoes. Bioresour. Technol. 2003, 89, 297–305. [Google Scholar] [CrossRef]
- Cabrera, M.; Chiang, S.; Merka, W.; Pancorbo, O.; Thompson, S. Nitrous oxide and carbon dioxide emissions from pelletized and nonpelletized poultry litter incorporated into soil. Plant Soil 1994, 163, 189–195. [Google Scholar] [CrossRef]
- Nishizawa, T.; Quan, A.; Kai, A.; Tago, K.; Ishii, S.; Shen, W.; Isobe, K.; Otsuka, S.; Senoo, K. Inoculation with N2-generating denitrifier strains mitigates N2O emission from agricultural soil fertilized with poultry manure. Biol. Fertil. Soils 2014, 50, 1001–1007. [Google Scholar] [CrossRef]
- Marshall, S.B.; Wood, C.W.; Braun, L.C.; Cabrera, M.L.; Mullen, M.D.; Guertal, E.A. Ammonia volatilization from tall fescue pastures fertilized with broiler litter. J. Environ. Qual. 1998, 1125–1129. [Google Scholar] [CrossRef]
- Pan, W.L.; Madsen, I.J.; Bolton, R.P.; Graves, L.; Sistrunk, T. Ammonia/ammonium toxicity root symptoms induced by inorganic and organic fertilizers and placement. Agron. J. 2016, 108, 2485–2492. [Google Scholar] [CrossRef] [Green Version]
- Barret, M.; Briand, M.; Bonneau, S.; Preveaux, A.; Valiere, S.; Bouchez, O.; Hunault, G.; Simoneau, P.; Jacquesa, M.A. Emergence shapes the structure of the seed microbiota. Appl. Environ. Microbiol. 2015, 81, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Zhou, X.; Zhang, J.; Cai, Z. Highly connected taxa located in the microbial network are prevalent in the rhizosphere soil of healthy plant. Biol. Fertil. Soils 2019, 55, 299–312. [Google Scholar] [CrossRef]
- Haas, D.; Defago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef]
- Passera, A.; Compant, S.; Casati, P.; Maturo, M.G.; Battelli, G.; Quaglino, G.; Antonielli, L.; Salerno, D.; Brasca, M.; Toffolatti, S.L. Not just a pathogen? Description of a plant-beneficial Pseudomonas syringae strain. Front. Microbiol. 2019, 10, 1409. [Google Scholar] [CrossRef] [Green Version]
- Rivas, R.; Velázquez, E.; Willems, A.; Vizcaíno, N.; Subba-Rao, N.S.; Mateos, P.F.; Gillis, M.; Dazzo, F.B.; Martínez-Molina, E. A new species of Devosia that forms a unique nitrogen-fixing root-nodule symbiosis with the aquatic legume Neptunia natans (L.f.) Druce. Appl. Environ. Microbiol. 2002, 68, 5217–5222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, C.; Penton, C.R.; Xiong, W.; Liu, C.; Wang, R.; Liu, Z.; Xu, X.; Li, R.; Shen, Q. Reshaping the rhizosphere microbiome by bio-organic amendment to enhance crop yield in a maize-cabbage rotation system. Appl. Soil Ecol. 2019, 142, 136–146. [Google Scholar] [CrossRef]
- Ofek, M.; Hadar, Y.; Minz, D. Ecology of root colonizing Massilia (Oxalobacteraceae). PLoS ONE 2012, 7, e40117. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Ravnskov, S.; Larsen, J.; Nicolaisen, M. Linking fungal communities in roots, rhizosphere, and soil to the health status of Pisum sativum. FEMS Microbiol. Ecol. 2012, 82, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, R.; Dunlap, C.; Barnett, S.; Franco, C.M.M. Decoding wheat endosphere-rhizosphere microbiomes in Rhizoctonia solani-infested soils challenged by Streptomyces biocontrol agents. Front. Plant Sci. 2019, 10, 1038. [Google Scholar] [CrossRef] [Green Version]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.; Zhao, Q.; Wu, H.; Jousset, A.; Shen, Q. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Biochem. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Fuchs, J.G.; Moënne-Loccoz, Y.; Défago, G. Nonpathogenic Fusarium oxysporum strain Fo47 induces resistance to Fusarium wilt in tomato. Plant Dis. 1997, 81, 492. [Google Scholar] [CrossRef] [Green Version]
- Amatulli, M.T.; Spadaro, D.; Gullino, M.L.; Garibaldi, A. Molecular identification of Fusarium spp. associated with bakanae disease of rice in Italy and assessment of their pathogenicity. Plant Pathol. 2010, 59, 839–844. [Google Scholar] [CrossRef]
- Niehaus, E.M.; Munsterkotter, M.; Proctor, R.H.; Brown, D.W.; Sharon, A.; Idan, Y.; Oren-Young, L.; Sieber, C.M.; Novak, O.; Pencik, A.; et al. Comparative “omics” of the Fusarium fujikuroi species complex highlights differences in genetic potential and metabolite synthesis. Genome Biol. Evol. 2016, 8, 3574–3599. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, J.; Aira, M.; Kolbe, A.R.; Gomez-Brandon, M.; Perez-Losada, M. Changes in the composition and function of bacterial communities during vermicomposting may explain beneficial properties of vermicompost. Sci. Rep. 2019, 9, 9657. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Brandón, M.; Domínguez, J. Recycling of solid organic wastes through vermicomposting: Microbial community changes throughout the process and use of vermicompost as a soil amendment. Crit. Rev. Environ. Sci. Technol. 2014, 44, 1289–1312. [Google Scholar] [CrossRef]
- Jack, A.L.H.; Thies, J.E. Compost and vermicompost as amendments promoting soil health. In Biological Approaches to Sustainable Soil Systems; Uphoff, N., Ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 453–466. [Google Scholar]
- Milkereit, J.; Geisseler, D.; Lazicki, P.; Settles, M.L.; Durbin-Johnson, B.P.; Hodson, A. Interactions between nitrogen availability, bacterial communities, and nematode indicators of soil food web function in response to organic amendments. Appl. Soil Ecol. 2021, 157. [Google Scholar] [CrossRef]
- Visioli, G.; Sanangelantoni, A.M.; Vamerali, T.; Dal Cortivo, C.; Blandino, M. 16S rDNA profiling to reveal the influence of seed-applied biostimulants on the rhizosphere of young maize plants. Molecules 2018, 23, 1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barajas, H.R.; Martinez-Sanchez, S.; Romero, M.F.; Alvarez, C.H.; Servin-Gonzalez, L.; Peimbert, M.; Cruz-Ortega, R.; Garcia-Oliva, F.; Alcaraz, L.D. Testing the two-step model of plant root microbiome acquisition under multiple plant species and soil sources. Front. Microbiol. 2020, 11, 542742. [Google Scholar] [CrossRef]
- Ou, Y.; Penton, C.R.; Geisen, S.; Shen, Z.; Sun, Y.; Lv, N.; Wang, B.; Ruan, Y.; Xiong, W.; Li, R.; et al. Deciphering underlying drivers of disease suppressiveness against pathogenic Fusarium oxysporum. Front. Microbiol. 2019, 10, 2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathma, J.; Sakthivel, N. Molecular and functional characterization of bacteria isolated from straw and goat manure based vermicompost. Appl. Soil Ecol. 2013, 70, 33–47. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Leeman, M.; Van Oorschot, M.; Van der Sluis, I.; Schippers, B.; Bakker, P.A. Dose-response relationships in biological control of Fusarium wilt of radish by Pseudomonas spp. Phytopathology 1995, 85, 1075–1080. [Google Scholar] [CrossRef]
- Mello, B.L.; Alessi, A.M.; McQueen-Mason, S.; Bruce, N.C.; Polikarpov, I. Nutrient availability shapes the microbial community structure in sugarcane bagasse compost-derived consortia. Sci. Rep. 2016, 6, 38781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gannes, V.; Eudoxie, G.; Hickey, W.J. Insights into fungal communities in composts revealed by 454-pyrosequencing: Implications for human health and safety. Front. Microbiol. 2013, 4, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.Z.; Ma, S.C.; Wang, S.P.; Wang, T.T.; Sun, Z.Y.; Tang, Y.Q.; Deng, Y.; Kida, K. A comparative study of composting the solid fraction of dairy manure with or without bulking material: Performance and microbial community dynamics. Bioresour. Technol. 2018, 247, 443–452. [Google Scholar] [CrossRef]
- Chang, Q.; Wang, W.; Regev-Yochay, G.; Lipsitch, M.; Hanage, W.P. Antibiotics in agriculture and the risk to human health: How worried should we be? Evol. Appl. 2015, 8, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Hester, E.; Harpenslager, S.; van Diggelen, J.; Lamers, L.; Jetten, M.; Luke, C.; Lucker, S.; Welte, C. Linking nitrogen load to the structure and function of wetland soil and rhizosphere microbial communities. mSystems 2018, 3, e00214–e00217. [Google Scholar] [CrossRef] [Green Version]
- Akinde, S.B.; Obire, O. Aerobic heterotrophic bacteria and petroleum-utilizing bacteria from cow dung and poultry manure. World J. Microbiol. Biotechnol. 2008, 24, 1999–2002. [Google Scholar] [CrossRef]
- Kay, E.; Bertolla, F.; Vogel, T.M.; Simonet, P. Opportunistic colonization of Ralstonia solanacearum-infected plants by Acinetobacter sp. and its natural competence development. Microb. Ecol. 2002, 43, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Su, J.F.; Zheng, S.C.; Huang, T.l.; Ma, F.; Shao, S.C.; Yang, S.F.; Zhang, L.N. Characterization of the anaerobic denitrification bacterium Acinetobacter sp. SZ28 and its application for groundwater treatment. Bioresour. Technol. 2015, 192, 654–659. [Google Scholar] [CrossRef]
- Willett, W.; Rockstrom, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT-Lancet Commission on healthy diets from sustainable food systems. Lancet 2019, 393, 447–492. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | % Volume Fort Vee | % Volume Organic Amendment | Adjusted N (g L−1) |
|---|---|---|---|
| Control | 100 | 0 | 1.3 |
| Vermicompost | 80 | 20 | 7.5 |
| Dairy manure compost | 80 | 20 | 7.5 |
| Poultry pellets | 80 | 10 * | 5.5 |
| Treatment | % Volume Field Soil | % Volume Amendment | % Volume Vermiculite | Bulk Density (g/cm3) |
|---|---|---|---|---|
| Control | 90 | 0 | 10 | 0.99 |
| Vermicompost | 70 | 20 | 10 | 0.88 |
| Dairy manure compost | 70 | 20 | 10 | 0.83 |
| Poultry pellets | 70 | 10 | 20 * | 0.85 |
| ESV ID | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| ESV_5 | Bacteroidetes | Bacteroidia | Cytophagales | Microscillaceae | Chryseolinea |
| ESV_6 | Proteobacteria | γ-Proteobacteria | Steroidobacterales | Steroidobacteraceae | Steroidobacter |
| ESV_4 | Proteobacteria | γ-Proteobacteria | β-Proteobacteriales | Burkholderiaceae | Massilia |
| ESV_3 | Proteobacteria | γ-Proteobacteria | β-Proteobacteriales | Burkholderiaceae | Massilia |
| ESV_9 | Proteobacteria | α-Proteobacteria | Rhizobiales | Devosiaceae | Devosia |
| ESV_7 | Proteobacteria | γ-Proteobacteria | Alteromonadales | Alteromonadaceae | Rheinheimera |
| ESV_13 | Bacteroidetes | Bacteroidia | Cytophagales | Microscillaceae | Chryseolinea |
| ESV_11 | Proteobacteria | γ-Proteobacteria | Xanthomonadales | Xanthomonadaceae | Luteimonas |
| ESV_12 | Bacteroidetes | Bacteroidia | Chitinophagales | Chitinophagaceae | Parafilimonas |
| ESV_8 | Proteobacteria | γ-Proteobacteria | Pseudomonadales | Moraxellaceae | Acinetobacter |
| ESV_10 | Proteobacteria | γ-Proteobacteria | Pseudomonadales | Pseudomonadaceae | Pseudomonas |
| ESV ID | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| ESV_2 | Mortierellomycota | Mortierellomycetes | Mortierellales | Mortierellaceae | Mortierella |
| ESV_4 | Ascomycota | Sordariomycetes | Hypocreales | Nectriaceae | Fusarium |
| ESV_3 | Mortierellomycota | Mortierellomycetes | Mortierellales | Mortierellaceae | Mortierella |
| ESV_5 | Ascomycota | Sordariomycetes | Sordariales | Chaetomiaceae | Zopfiella |
| ESV_11 | Ascomycota | Sordariomycetes | Sordariales | Chaetomiaceae | Zopfiella |
| ESV_9 | Ascomycota | Sordariomycetes | Microascales | Microascaceae | Scedosporium |
| ESV_13 | Mortierellomycota | Mortierellomycetes | Mortierellales | Mortierellaceae | Mortierella |
| ESV_8 | Basidiomycota | Unknown | Unknown | Unknown | Unknown |
| ESV_12 | Ascomycota | Eurotiomycetes | Onygenales | Incertae sedis | Chrysosporium |
| ESV_6 | Mortierellomycota | Mortierellomycetes | Mortierellales | Mortierellaceae | Mortierella |
| ESV_2 | Mortierellomycota | Mortierellomycetes | Mortierellales | Mortierellaceae | Mortierella |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Readyhough, T.; Neher, D.A.; Andrews, T. Organic Amendments Alter Soil Hydrology and Belowground Microbiome of Tomato (Solanum lycopersicum). Microorganisms 2021, 9, 1561. https://doi.org/10.3390/microorganisms9081561
Readyhough T, Neher DA, Andrews T. Organic Amendments Alter Soil Hydrology and Belowground Microbiome of Tomato (Solanum lycopersicum). Microorganisms. 2021; 9(8):1561. https://doi.org/10.3390/microorganisms9081561
Chicago/Turabian StyleReadyhough, Taylor, Deborah A. Neher, and Tucker Andrews. 2021. "Organic Amendments Alter Soil Hydrology and Belowground Microbiome of Tomato (Solanum lycopersicum)" Microorganisms 9, no. 8: 1561. https://doi.org/10.3390/microorganisms9081561
APA StyleReadyhough, T., Neher, D. A., & Andrews, T. (2021). Organic Amendments Alter Soil Hydrology and Belowground Microbiome of Tomato (Solanum lycopersicum). Microorganisms, 9(8), 1561. https://doi.org/10.3390/microorganisms9081561