
Effects of Efficient Expression of Vitreoscilla Hemoglobin on Production, Monosaccharide Composition, and Antioxidant Activity of Exopolysaccharides in Ganoderma lucidum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
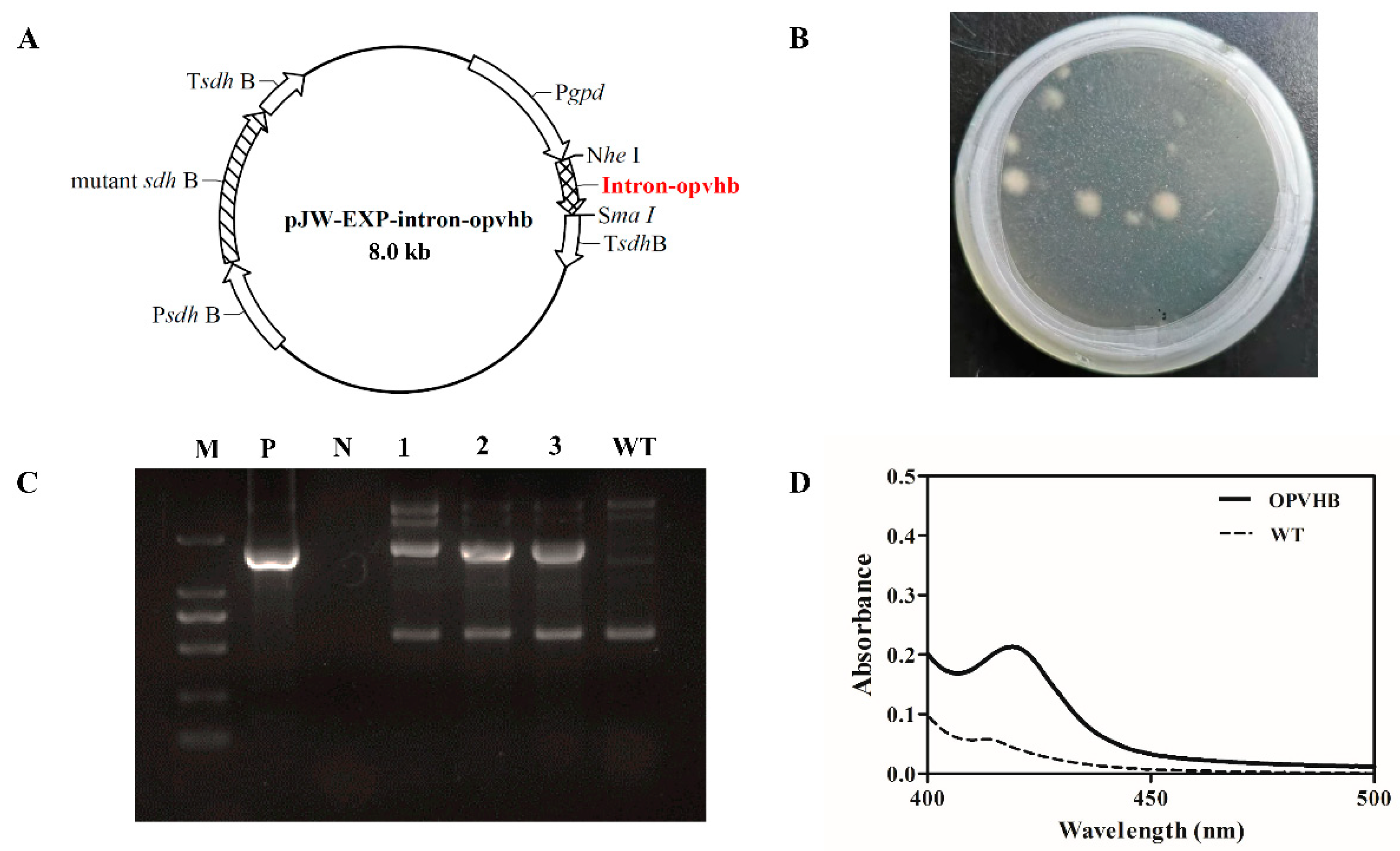
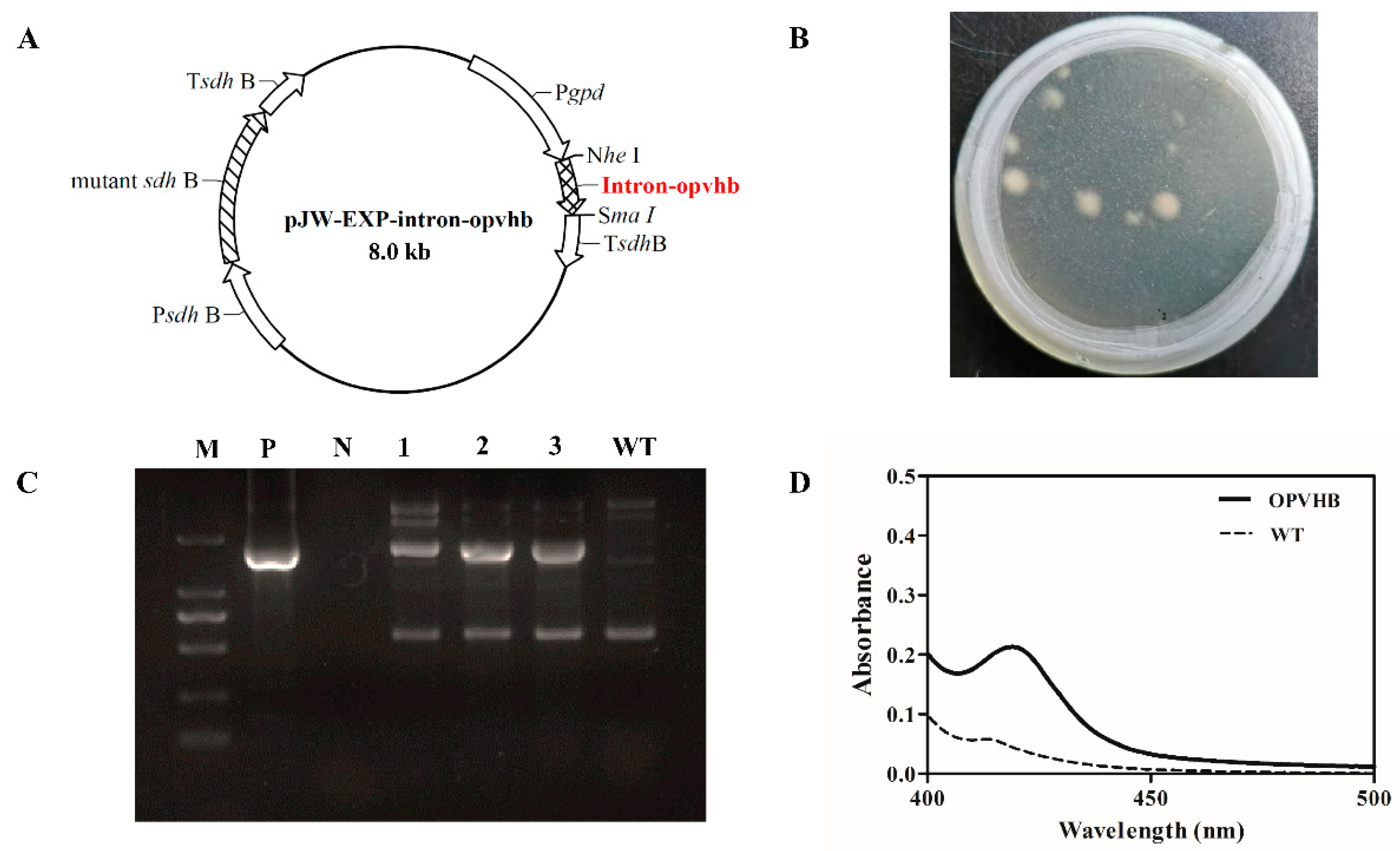
2.2. Construction of Plasmid pJW-EXP-Intron-Opvhb
2.3. Genetic Transformation of G. lucidum Protoplasts and Identification of Transformants
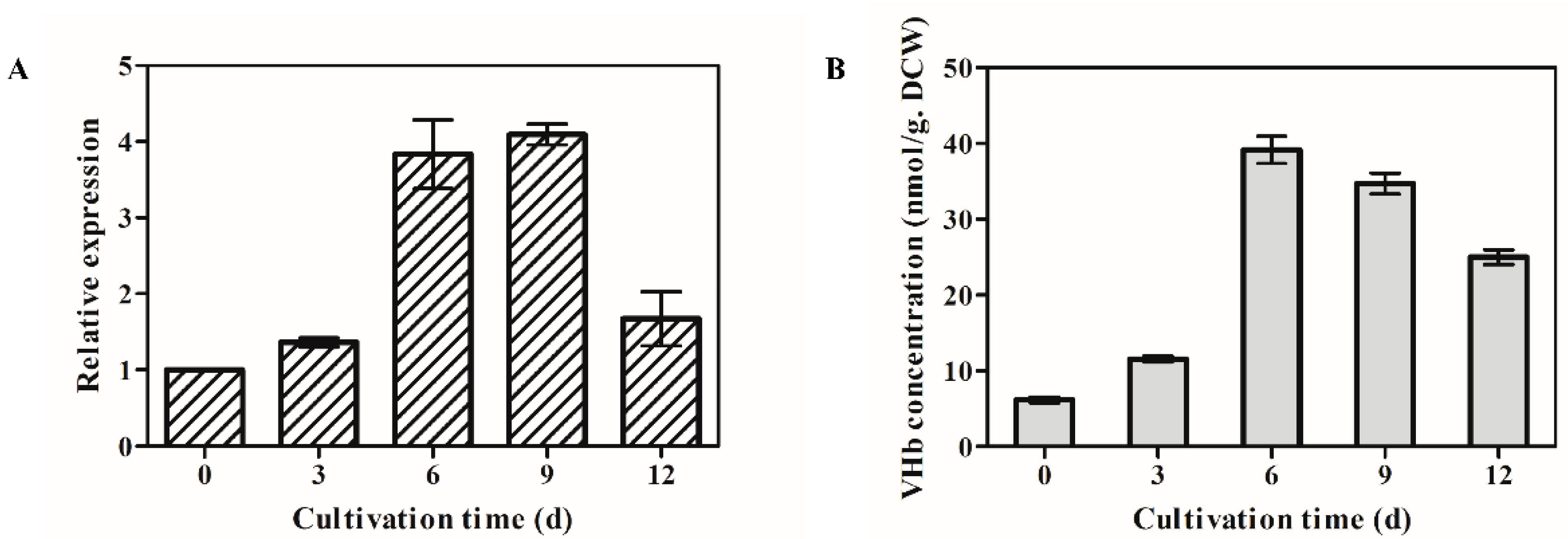
2.4. Transcription Analysis of OPVHb Gene Using Quantitative Real-Time PCR (qRT-PCR)
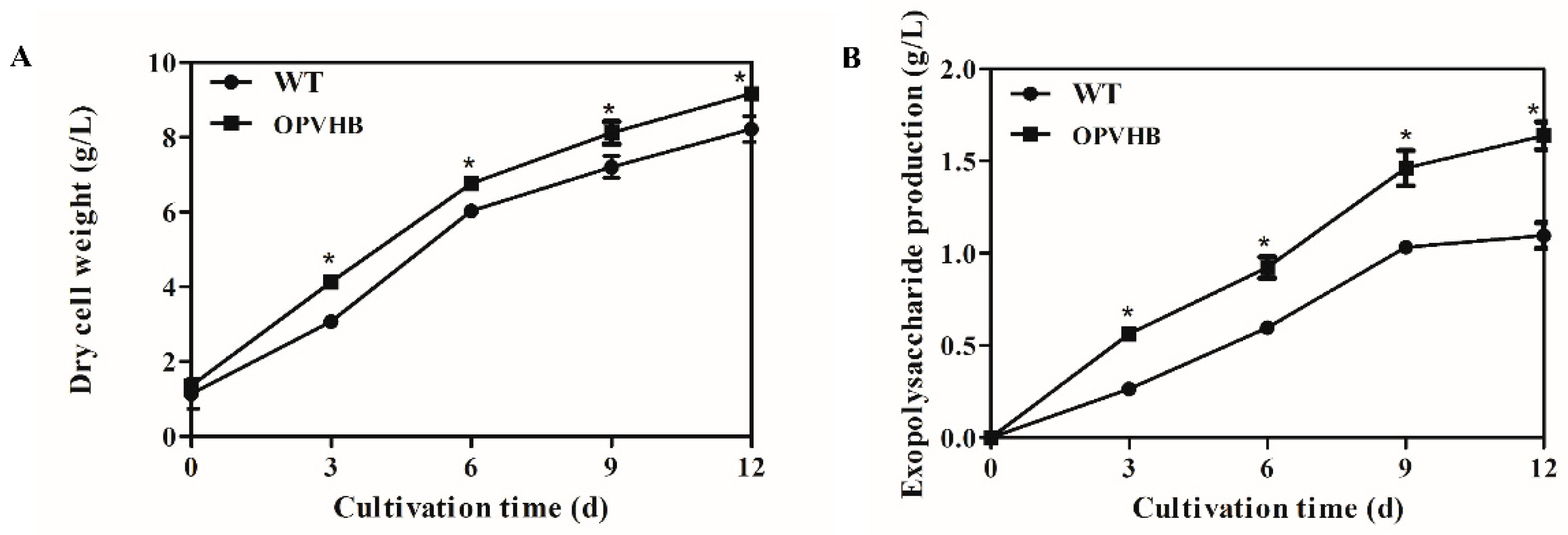
2.5. Production of Biomass and Exopolysaccharides
2.6. Extraction of G. lucidum Exopolysaccharides
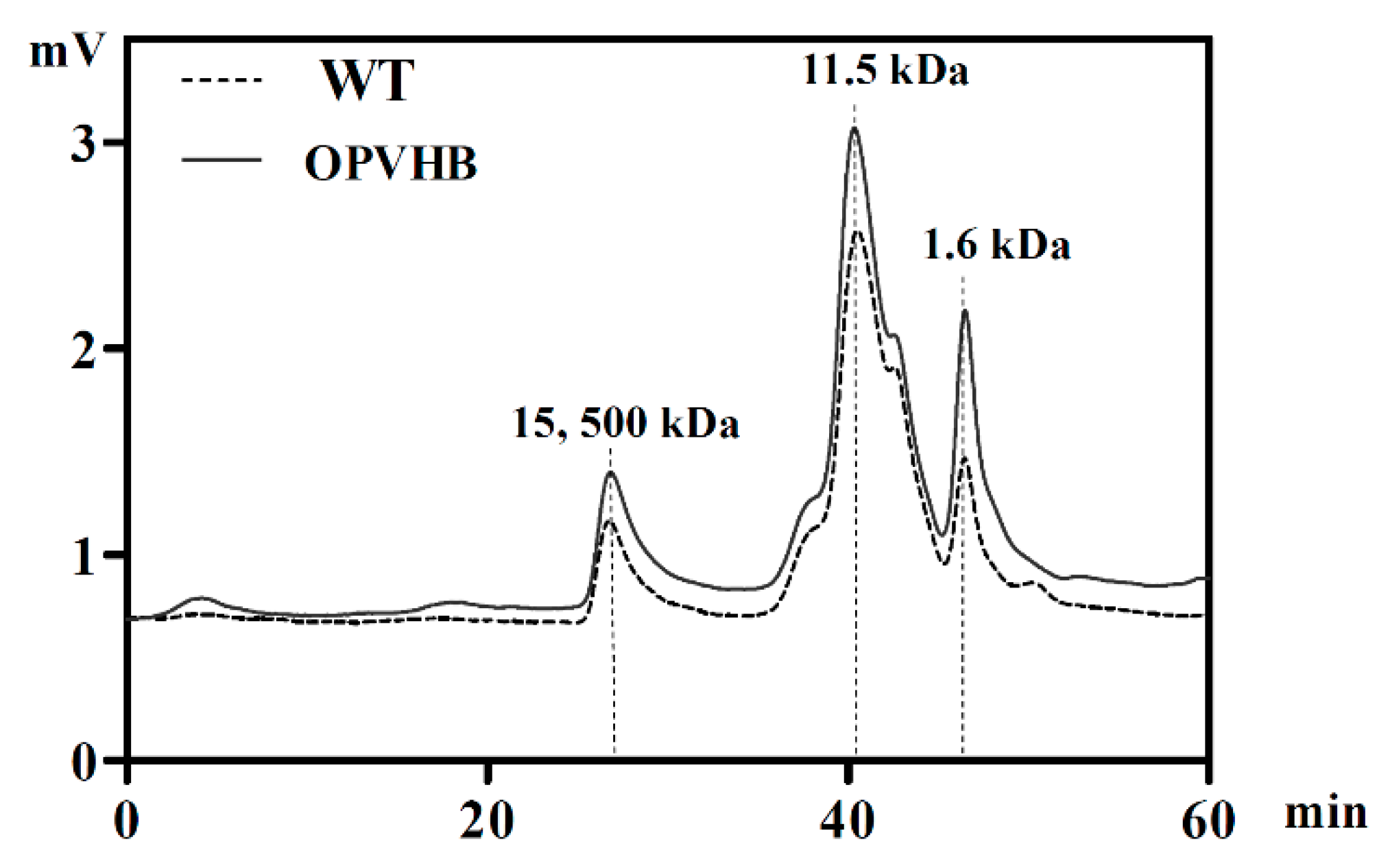
2.7. Determination of Exopolysaccharide Molecular Weight
2.8. Analysis of Monosaccharide Compositions
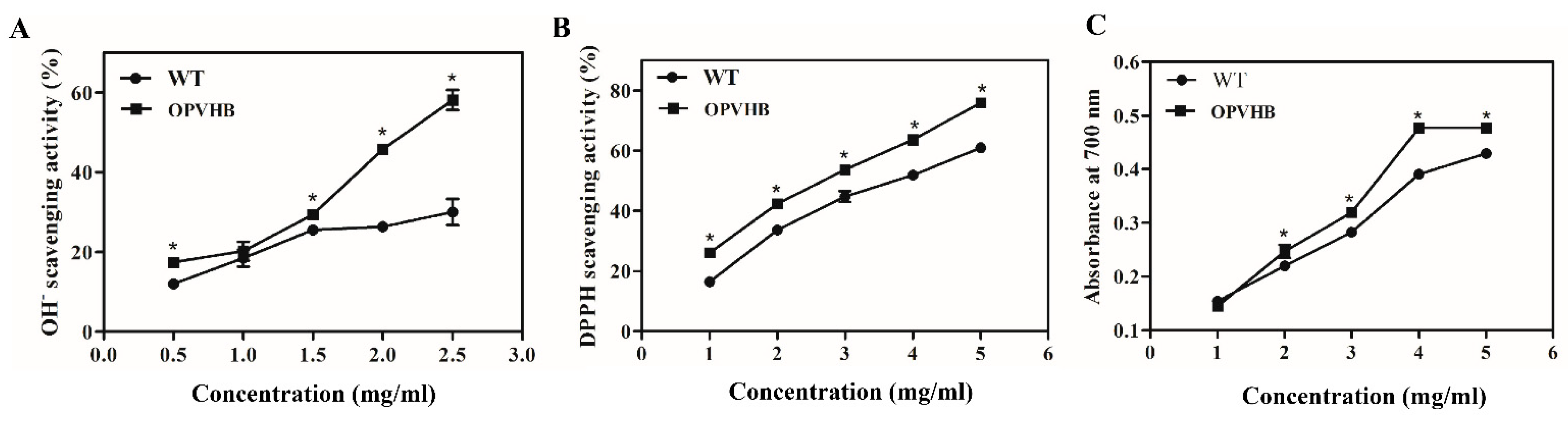
2.9. Measurements of Antioxidant Activity
2.10. Statistical Analysis
3. Results and Discussion
3.1. Expression of the VHb Gene in G. lucidum
3.2. Profiles of VHb Expression in the Transformant OPVHB
3.3. Increased Production of Exopolysaccharides by Expression of Opvhb in G. lucidum
3.4. Effects of Opvhb Expression on Molecular Weight Distribution and Monosaccharide Composition of G. lucidum Exopolysaccharides
3.5. Effect of Efficient Opvhb Expression on Antioxidant Activity of Exopolysaccharides
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hsu, K.D.; Cheng, K.C. From nutraceutical to clinical trial: Frontiers in Ganoderma development. Appl. Microbiol. Biotechnol. 2018, 102, 9037–9051. [Google Scholar] [CrossRef] [PubMed]
- Sliva, D. Ganoderma lucidum in cancer research. Leuk. Res. 2006, 30, 767–768. [Google Scholar] [CrossRef] [PubMed]
- Boh, B.; Verovic, M.; Zhang, J.; Zhi-Bin, L. Ganoderma lucidum and its pharmaceutically active compounds. Biotechnol. Annu. Rev. 2007, 13, 265–301. [Google Scholar]
- Lu, J.H.; He, R.J.; Sun, P.L.; Zhang, F.M.; Linhardt, R.J.; Zhang, A.Q. Molecular mechanisms of bioactive polysaccharides from Ganoderma lucidum (Lingzhi), a review. Int. J. Biol. Macromol. 2020, 150, 765–774. [Google Scholar] [CrossRef]
- Fang, Q.H.; Zhong, J.J. Submerged fermentation of higher fungus Ganoderma lucidum for production of valuable bioactive metabolites-ganoderic acid and polysaccharide. Biochem. Eng. J. 2002, 10, 61–65. [Google Scholar] [CrossRef]
- Habijanic, J.; Berovic, M.; Boh, B.; Wraber, B.; Petravic-Tominac, V. Production of biomass and polysaccharides of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidm (W. Curt.:Fr.) P. Karst. (higher basidiomycetes). Int. J. Med. Mushrooms 2013, 15, 81–90. [Google Scholar] [CrossRef]
- Hsieh, C.; Tseng, M.H.; Liu, C.J. Production of polysaccharides from Ganoderma lucidum (CCRC 36041) under limitations of nutrients. Enzyme Microb. Technol. 2006, 38, 109–117. [Google Scholar] [CrossRef]
- Liu, G.Q.; Zhang, K.C. Enhancement of polysaccharides production in Ganoderma lucidum by the addition of ethyl acetate extracts from Eupolyphaga sinensis and Catharsius molossus. Appl. Microbiol. Biotechnol. 2007, 74, 572–577. [Google Scholar] [CrossRef]
- Zhu, L.W.; Tang, Y.J. Significance of protein elicitor isolated from Tuber melanosporum on the production of ganoderic acid and Ganoderma polysaccharides during the fermentation of Ganoderma lucidum. Bioprocess Biosyst. Eng. 2010, 33, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.J.; Zhang, W.; Zhong, J.J. Performance analyses of a pH-shift and DOT-shift integrated fed-batch fermentation process for the production of ganoderic acid and Ganoderma polysaccharides by medicinal mushroom Ganoderma lucidum. Bioresour. Technol. 2009, 100, 1852–1859. [Google Scholar] [CrossRef]
- Zhu, L.W.; Zhong, J.J.; Tang, Y.J. Multi-fed batch culture integrated with three-stage light irradiation and multiple additions of copper ions for thehyperproduction of ganoderic acid and Ganoderma polysaccharides by the medicinal mushroom Ganoderma lucidum. Process Biochem. 2010, 45, 1904–1911. [Google Scholar] [CrossRef]
- Ji, S.L.; Liu, R.; Ren, M.F.; Li, H.J.; Xu, J.W. Enhanced production of polysaccharide through overexpression of homologous uridine diphosphateglucose pyrophosphorylase gene in a submerged culture of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes). Int. J. Med. Mushrooms 2015, 17, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.W.; Ji, S.L.; Li, H.J.; Zhou, J.S.; Duan, Y.Q.; Dang, L.Z.; Mo, M.H. Increased polysaccharide production and biosynthetic gene expressions in a submergedculture of Ganoderma lucidum by the overexpression of the homologous-phosphoglucomutase gene. Bioprocess Biosyst. Eng. 2015, 38, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Stark, B.C.; Pagilla, K.R.; Dikshit, K.L. Recent applications of Vitreoscilla hemoglobin technology in bioproduct synthesis and bioremediation. Appl. Microbiol. Biotechnol. 2015, 99, 1627–1636. [Google Scholar] [CrossRef]
- Chien, L.J.; Lee, C.K. Enhanced hyaluronic acid production in Bacillus subtilis by coexpressing bacterial hemoglobin. Biotechnol. Prog. 2007, 23, 1017–1022. [Google Scholar] [CrossRef]
- Wu, X.C.; Chen, Y.M.; Li, Y.D.; Li, O.; Zhu, L.; Qian, C.D.; Tao, X.L.; Teng, Y. Constitutive expression of Vitreoscilla haemoglobin in Sphingomonas elodea to improve gellan gum production. J. Appl. Microbiol. 2010, 110, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Zhu, P.; Jiang, R.F.; Wu, L.T.; Feng, X.H.; Li, S.; Xu, H. Enhancement of welan gum production in Sphingomonas sp. HT-1 via heterologous expression of Vitreoscilla hemoglobin gene. Carbohydr. Polym. 2017, 156, 135–142. [Google Scholar] [CrossRef]
- Xue, S.J.; Jiang, H.; Chen, L.; Ge, N.; Liu, G.L.; Hu, Z.; Chi, Z.M.; Chi, Z. Over-expression of Vitreoscilla hemoglobin (VHb) and flavohemoglobin (FHb) genes greatly enhances pullulan production. Int. J. Biol. Macromol. 2019, 132, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Sun, S.; Zhang, S. Enhanced production of total flavones and exopolysaccharides via Vitreoscilla hemoglobin biosynthesis in Phellinus igniarius. Bioresour. Technol. 2011, 102, 1747–1751. [Google Scholar] [CrossRef]
- Li, H.J.; Zhang, D.H.; Yue, T.H.; Jiang, L.X.; Yu, X.; Zhao, P.; Li, T.; Xu, J.W. Improved polysaccharide production in a submerged culture of Ganoderma lucidum by the heterologous expression of Vitreoscilla hemoglobin gene. J. Biotechnol. 2016, 217, 132–137. [Google Scholar] [CrossRef]
- Liu, K.; Sun, B.; You, H.; Tu, J.L.; Yu, X.; Zhao, P.; Xu, J.W. Dual sgRNA-directed gene deletion in basidiomycete Ganoderma lucidum using the CRISPR/Cas9 system. Microb. Biotechnol. 2020, 13, 386–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, T.T.; Chang, C.A.; Chiu, K.H.; Tsay, P.K.; Jen, J.F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.Y.; Pang, X.B.; Yao, W.B.; Gao, X.D. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 46, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef]
- Soltani, M.; Kamyab, H.; El-Enshasy, H.A. Molecular weight (Mw) and monosaccharide composition (MC): Two major factors affecting the therapeutic action of polysaccharides extracted from Cordyceps sinensis-mini review. J. Pure Appl. Microbiol. 2013, 7, 1601–1613. [Google Scholar]
- Fraga, I.; Coutinho, J.; Bezerra, R.M.; Dias, A.A.; Marques, G.; Nunes, F.M. Influence of culture medium growth variables on Ganoderma lucidum exopolysaccharides structural features. Carbohydr. Polym. 2014, 111, 936–946. [Google Scholar] [CrossRef]
- Peng, L.; Qiao, S.K.; Xu, Z.H.; Guan, F.; Ding, Z.Y.; Gu, Z.H.; Zhang, L.; Shi, G.Y. Effects of culture conditions on monosaccharide composition of Ganoderma lucidum exopolysaccharide and on activities of related enzymes. Carbohydr. Polym. 2015, 133, 104–109. [Google Scholar] [CrossRef]
- Peng, L.; Li, J.; Liu, Y.; Xu, Z.H.; Wu, J.Y.; Ding, Z.Y.; Gu, Z.H.; Zhang, L.; Shi, G.Y. Effects of mixed carbon sources on galactose and mannose content of exopolysaccharides and related enzyme activities in Ganoderma lucidum. RSC Adv. 2016, 6, 39284–39291. [Google Scholar] [CrossRef]
- Sun, B.; You, H.; Xu, J.W. Enhancement of ganoderic acid production by promoting sporulation in a liquid static culture of Ganoderma species. J. Biotechnol. 2021, 328, 72–77. [Google Scholar] [CrossRef]
- Xu, J.W.; Xu, Y.N.; Zhong, J.J. Production of individual ganoderic acids and expression of biosynthetic genes in liquid static and shaking cultures of Ganoderma lucidum. Appl. Microbiol. Biotechnol. 2010, 85, 941–948. [Google Scholar] [CrossRef]
- Xu, J.W.; Xu, Y.N.; Zhong, J.J. Enhancement of ganoderic acid accumulation by overexpression of an N-terminally truncated 3-hydroxy-3-methylglutaryl coenzyme A reductase gene in the basidiomycete Ganoderma lucidum. Appl. Environ. Microbiol. 2012, 78, 7968–7976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Ji, S.L.; He, Y.L.; Ren, M.F.; Xu, J.W. Development of an expression plasmid and its use in genetic manipulation of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes). Int. J. Med. Mushrooms 2014, 16, 161–168. [Google Scholar] [CrossRef]
- Zhang, D.H.; Jiang, L.X.; Li, N.; Yu, X.Y.; Zhao, P.; Li, T.; Xu, J.W. Overexpression of the squalene epoxidase gene alone and in combination with the 3-hydroxy-3-methylglutaryl coenzyme A gene increases ganoderic acid production in Ganoderma lingzhi. J. Agric. Food Chem. 2017, 65, 4683–4690. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.M.; Wang, S.Y.; Fu, W.C. Lower temperature cultures enlarge the effects of Vitreoscilla hemoglobin expression on recombinant Pichia pastoris. Int. J. Mol. Sci. 2012, 13, 13212–13226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 24, 425–436. [Google Scholar] [CrossRef]
- Dai, J.; Wu, Y.; Chen, S.W.; Zhu, S.; Ying, H.P.; Wang, M.; Tang, J. Sugar compositional determination of polysaccharides from Dunaliella salina by modified RP-HPLC method of precolumn derivatization with 1-phenyl-3-methyl-5-pyrazolone. Carbohydr. Polym. 2010, 82, 629–635. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, M.; Zhai, G.Y.; Ma, Z.; Wang, L.Q.; Jia, L. Antioxidant and anti-ageing activities of mycelia zinc polysaccharide from Pholiota nameko SW-03. J. Sci. Food Agric. 2015, 95, 3117–3126. [Google Scholar] [CrossRef]
- Zhang, H.; Zou, P.; Zhao, H.T.; Qiu, J.Q.; Regenstein, J.M.; Yang, X. Isolation, purification, structure and antioxidant activity of polysaccharide from pinecones of Pinus koraiensis. Carbohydr. Polym. 2021, 251, 117078. [Google Scholar] [CrossRef] [PubMed]
- Mora-Lugo, R.; Madrigal, M.; Yelemane, V.; Fernande-Lahore, M. Improved biomass and protein production in solid-state cultures of an Aspergillus sojae strain harboring the Vitreoscilla hemoglobin. Appl. Microbiol. Biotechnol. 2015, 99, 9699–9708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Dong, S.; Li, S.; Xu, X.; Xu, H. Improvement of welan gum biosynthesisand transcriptional analysis of the genes responding to enhanced oxygen transfer by oxygen vectors in Sphingomonas sp. Biochem. Eng. J. 2014, 90, 264–271. [Google Scholar] [CrossRef]
- Barahona, T.; Chandia, N.P.; Encinas, M.V.; Matsuhiro, B.; Zuniga, E.A. Antioxidant capacity of sulfated polysaccharides from seaweeds. A kinetic approach. Food Hydrocoll. 2011, 25, 529–535. [Google Scholar] [CrossRef]
- Ji, X.L.; Hou, C.Y.; Yan, Y.Z.; Shi, M.M.; Liu, Y.Q. Comparison of structural characterization and antioxidant activity of polysaccharides from jujube (Ziziphus jujuba Mill.) fruit. Int. J. Biol. Macromol. 2020, 149, 1008–1018. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exopolysaccharides | Monosaccharide Compostion (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Glc | Gal | Man | Fuc | Rha | Xyl | Ara | GlcUA | |
| WT | 36.3 ± 0.5 | 19.7 ± 0.4 | 24.9 ± 0.6 | 4.3 ± 0.5 | 2.6 ± 0.1 | 2.8 ± 0.1 | 2.9 ± 0.1 | 6.5 ± 0.3 |
| OPVHB | 35.4 ± 0.4 * | 23.1 ± 0.2 * | 31.5 ± 0.5 * | 3.2 ± 0.2 * | 1.4 ± 0.1 * | 0.9 ± 0 * | 1.1 ± 0.1 * | 3.4 ± 0.4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-X.; Li, N.; Xu, J.-W. Effects of Efficient Expression of Vitreoscilla Hemoglobin on Production, Monosaccharide Composition, and Antioxidant Activity of Exopolysaccharides in Ganoderma lucidum. Microorganisms 2021, 9, 1551. https://doi.org/10.3390/microorganisms9081551
Wang Z-X, Li N, Xu J-W. Effects of Efficient Expression of Vitreoscilla Hemoglobin on Production, Monosaccharide Composition, and Antioxidant Activity of Exopolysaccharides in Ganoderma lucidum. Microorganisms. 2021; 9(8):1551. https://doi.org/10.3390/microorganisms9081551
Chicago/Turabian StyleWang, Zi-Xu, Na Li, and Jun-Wei Xu. 2021. "Effects of Efficient Expression of Vitreoscilla Hemoglobin on Production, Monosaccharide Composition, and Antioxidant Activity of Exopolysaccharides in Ganoderma lucidum" Microorganisms 9, no. 8: 1551. https://doi.org/10.3390/microorganisms9081551
APA StyleWang, Z.-X., Li, N., & Xu, J.-W. (2021). Effects of Efficient Expression of Vitreoscilla Hemoglobin on Production, Monosaccharide Composition, and Antioxidant Activity of Exopolysaccharides in Ganoderma lucidum. Microorganisms, 9(8), 1551. https://doi.org/10.3390/microorganisms9081551