
Nanomotion Spectroscopy as a New Approach to Characterize Bacterial Virulence


 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. AFM Cantilever Preparation
2.3. Measurements
2.4. Microscopic Characterization of the Bacteria on the Cantilever
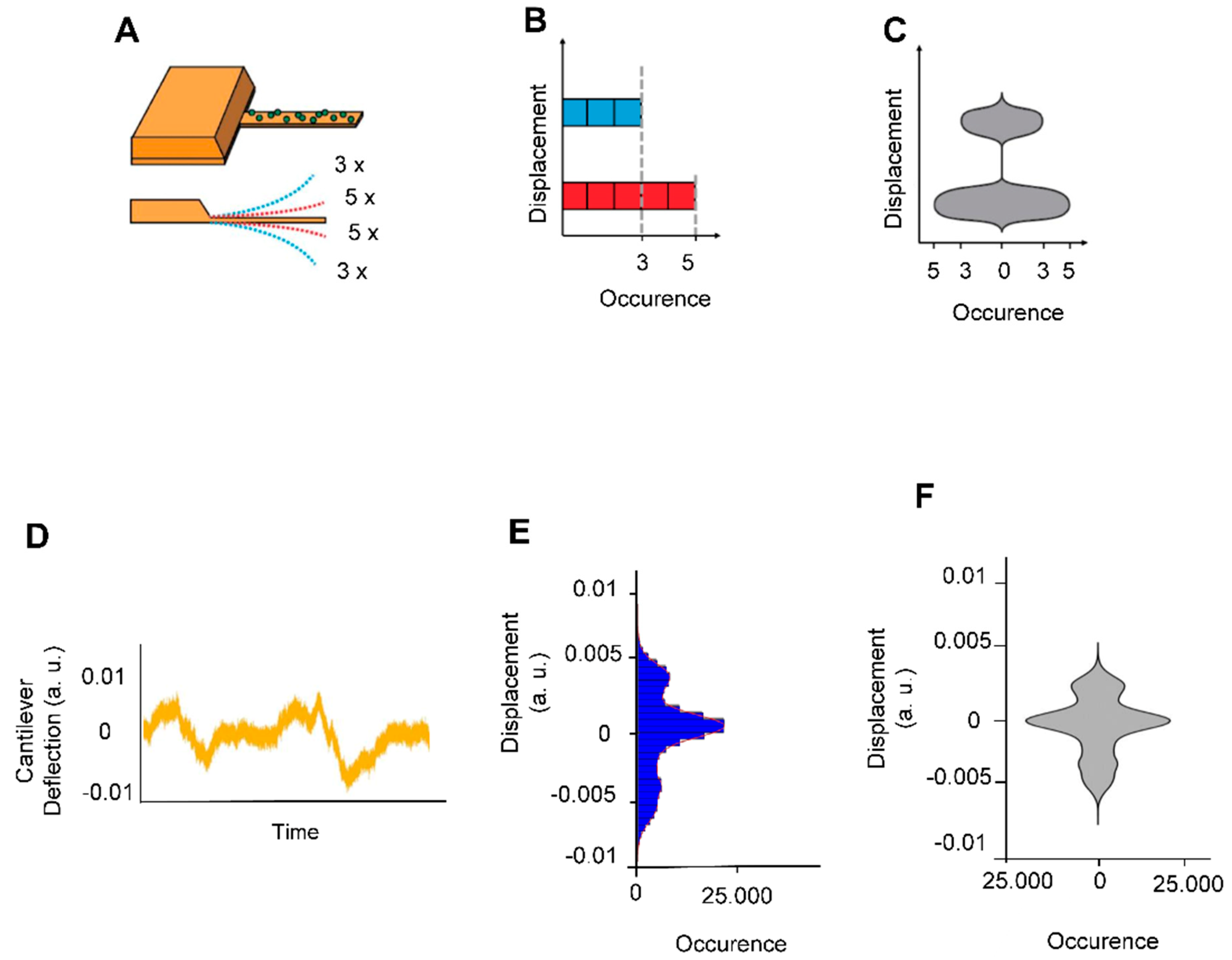
2.5. Data Processing
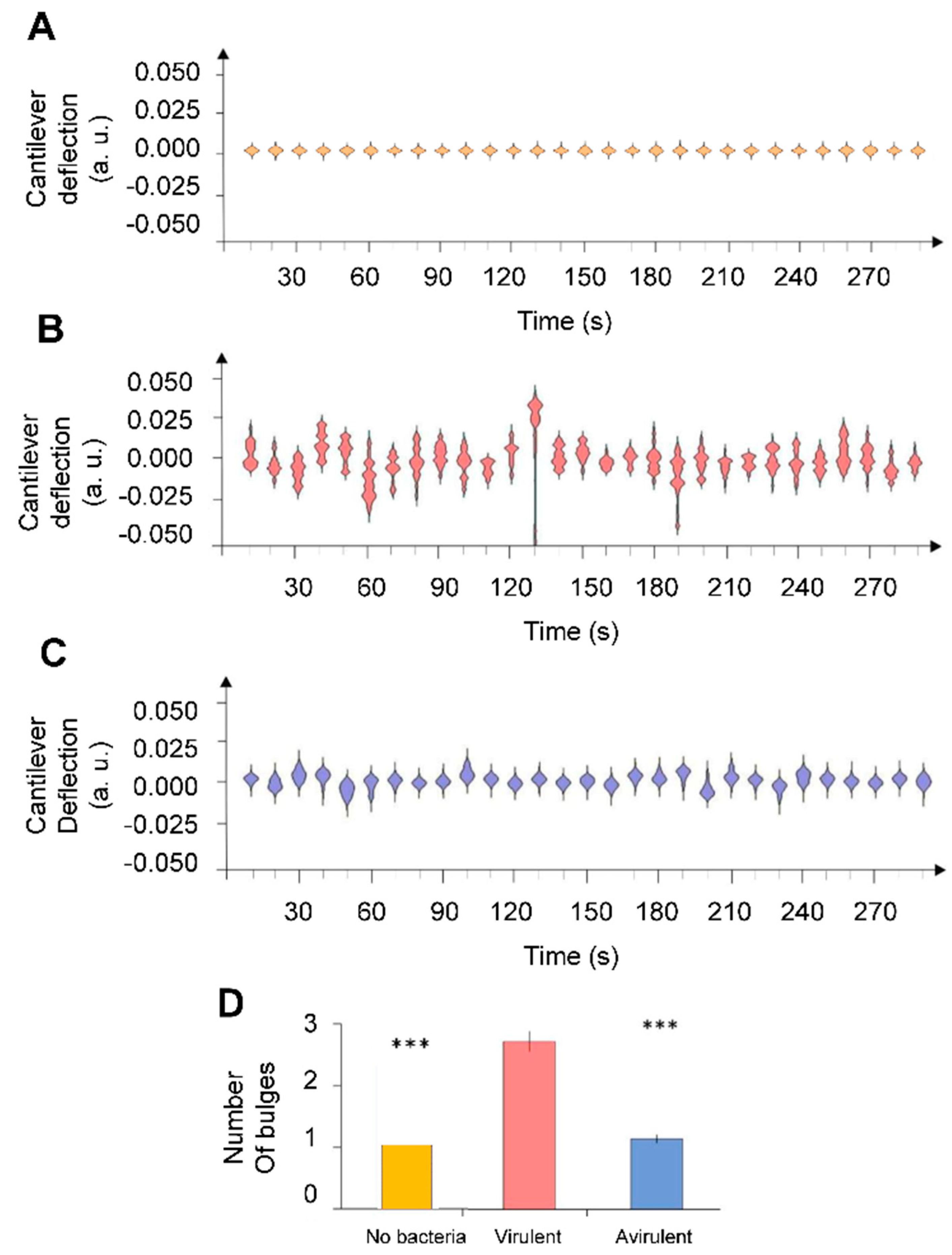
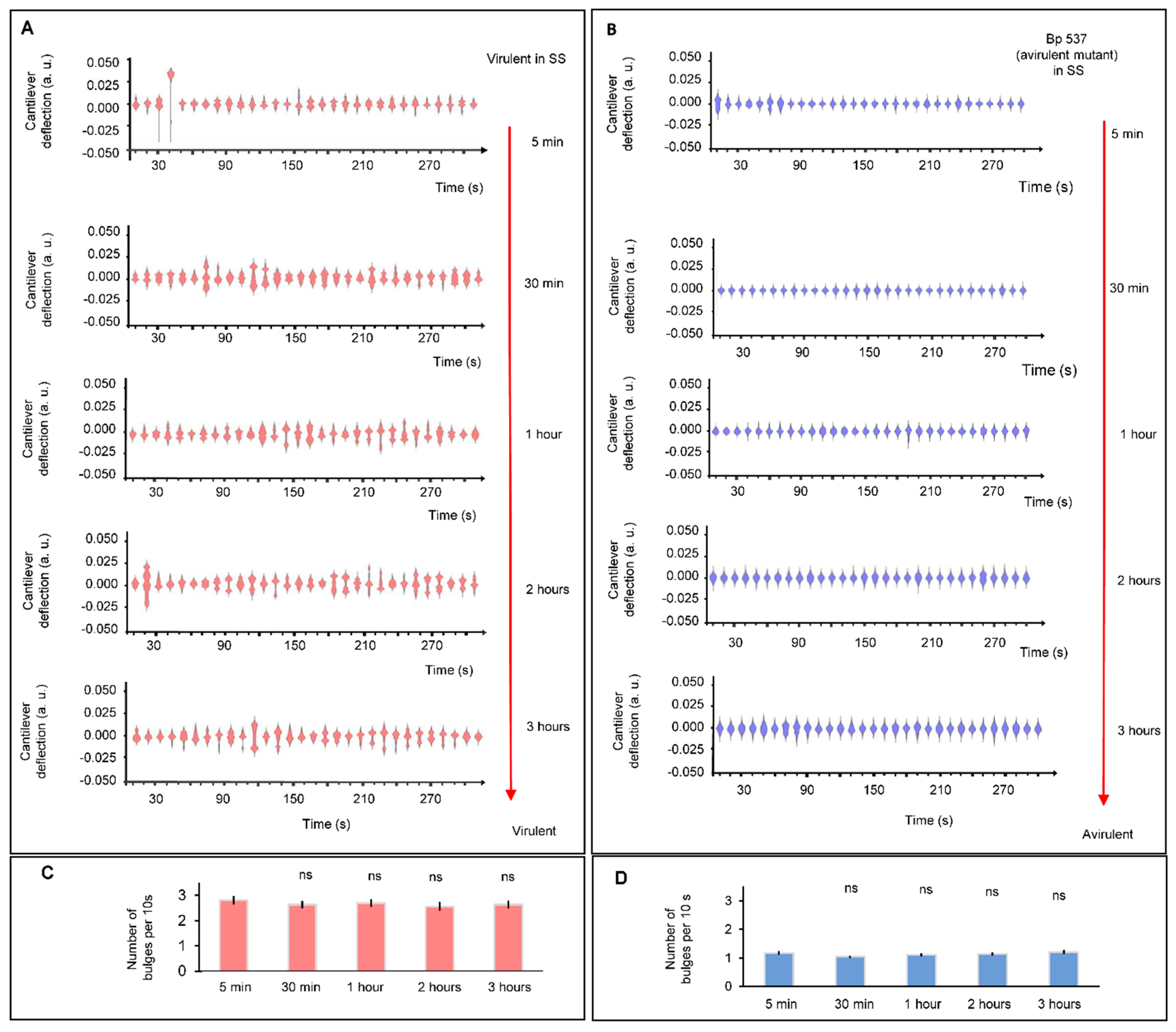
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug. Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [Green Version]
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A Review on Antibiotic Resistance: Alarm Bells Are Ringing. Cureus 2017, 9, e1403. [Google Scholar] [CrossRef] [Green Version]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The Threat of Antimicrobial Resistance in Developing Countries: Causes and Control Strategies. Antimicrob. Resist. Infect. Control. 2017, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Longo, G.; Alonso-Sarduy, L.; Rio, L.M.; Bizzini, A.; Trampuz, A.; Notz, J.; Dietler, G.; Kasas, S. Rapid Detection of Bacterial Resistance to Antibiotics Using AFM Cantilevers as Nanomechanical Sensors. Nat. Nanotechnol. 2013, 8, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Aghayee, S.; Benadiba, C.; Notz, J.; Kasas, S.; Dietler, G.; Longo, G. Combination of Fluorescence Microscopy and Nanomotion Detection to Characterize Bacteria. J. Mol. Recognit. 2013, 26, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Kasas, S.; Ruggeri, F.S.; Benadiba, C.; Maillard, C.; Stupar, P.; Tournu, H.; Dietler, G.; Longo, G. Detecting Nanoscale Vibrations as Signature of Life. Proc. Natl. Acad. Sci. USA 2015, 112, 378–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupar, P.; Opota, O.; Longo, G.; Prod’hom, G.; Dietler, G.; Greub, G.; Kasas, S. Nanomechanical Sensor Applied to Blood Culture Pellets: A Fast Approach to Determine the Antibiotic Susceptibility against Agents of Bloodstream Infections. Clin. Microbiol. Infect. 2017, 23, 400–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustazzolu, A.; Venturelli, L.; Dinarelli, S.; Brown, K.; Floto, R.A.; Dietler, G.; Fattorini, L.; Kasas, S.; Girasole, M.; Longo, G. A Rapid Unraveling of the Activity and Antibiotic Susceptibility of Mycobacteria. Antimicrob. Agents. Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Finger, H.; von Koenig, C.H.W. Bordetella. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Melton, A.R.; Weiss, A.A. Environmental Regulation of Expression of Virulence Determinants in Bordetella Pertussis. J. Bacteriol. 1989, 171, 6206–6212. [Google Scholar] [CrossRef] [Green Version]
- Martinez de Tejada, G.; Cotter, P.A.; Heininger, U.; Camilli, A.; Akerley, B.J.; Mekalanos, J.J.; Miller, J.F. Neither the Bvg- Phase nor the Vrg6 Locus of Bordetella Pertussis Is Required for Respiratory Infection in Mice. Infect. Immun. 1998, 66, 2762–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, K.; Bonocora, R.P.; Kim, D.D.; Chen, Q.; Wade, J.T.; Stibitz, S.; Hinton, D.M. The BvgAS Regulon of Bordetella Pertussis. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Gouw, D.; Hermans, P.W.M.; Bootsma, H.J.; Zomer, A.; Heuvelman, K.; Diavatopoulos, D.A.; Mooi, F.R. Differentially Expressed Genes in Bordetella Pertussis Strains Belonging to a Lineage Which Recently Spread Globally. PLoS ONE 2014, 9, e84523. [Google Scholar] [CrossRef]
- Hot, D.; Antoine, R.; Renauld-Mongénie, G.; Caro, V.; Hennuy, B.; Levillain, E.; Huot, L.; Wittmann, G.; Poncet, D.; Jacob-Dubuisson, F.; et al. Differential Modulation of Bordetella Pertussis Virulence Genes as Evidenced by DNA Microarray Analysis. Mol. Genet. Genomics 2003, 269, 475–486. [Google Scholar] [CrossRef]
- Relman, D.; Tuomanen, E.; Falkow, S.; Golenbock, D.T.; Saukkonen, K.; Wright, S.D. Recognition of a Bacterial Adhesion by an Integrin: Macrophage CR3 (Alpha M Beta 2, CD11b/CD18) Binds Filamentous Hemagglutinin of Bordetella Pertussis. Cell 1990, 61, 1375–1382. [Google Scholar] [CrossRef]
- Stainer, D.W.; Scholte, M.J. A Simple Chemically Defined Medium for the Production of Phase I Bordetella Pertussis. J. Gen. Microbiol. 1970, 63, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Venturelli, L.; Kohler, A.-C.; Stupar, P.; Villalba, M.I.; Kalauzi, A.; Radotic, K.; Bertacchi, M.; Dinarelli, S.; Girasole, M.; Pešić, M.; et al. Perspective View on the Nanomotion Detection of Living Organisms and Its Features. J. Mol. Recognit. 2020, 33, e2849. [Google Scholar] [CrossRef]
- Hoffmann, Holger. Violin Plot, MATLAB Central File Exchange. 2021. Available online: Www.Mathworks.Com/Matlabcentral/Fileexchange/45134-Violin-Plot (accessed on 5 March 2020).
- Metz, B.; Hoonakker, M.; Uittenbogaard, J.P.; Weyts, M.; Mommen, G.P.M.; Meiring, H.D.; Tilstra, W.; Pennings, J.L.A.; van der Pol, L.A.; Kuipers, B.; et al. Proteome Analysis Is a Valuable Tool to Monitor Antigen Expression during Upstream Processing of Whole-Cell Pertussis Vaccines. J. Proteome. Res. 2017, 16, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.A.; Falkow, S. Genetic Analysis of Phase Change in Bordetella Pertussis. Infect. Immun. 1984, 43, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Gonyar, L.A.; Gelbach, P.E.; McDuffie, D.G.; Koeppel, A.F.; Chen, Q.; Lee, G.; Temple, L.M.; Stibitz, S.; Hewlett, E.L.; Papin, J.A.; et al. In Vivo Gene Essentiality and Metabolism in Bordetella Pertussis. mSphere 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Karataev, G.I.; Sinyashina, L.N.; Medkova, A.Y.; Semin, E.G.; Shevtsova, Z.V.; Matua, A.Z.; Kondzariya, I.G.; Amichba, A.A.; Kubrava, D.T.; Mikvabia, Z.Y. Insertional Inactivation of Virulence Operon in Population of Persistent Bordetella pertussis Bacteria. Genetika 2016, 52, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Veal-Carr, W.L.; Stibitz, S. Demonstration of Differential Virulence Gene Promoter Activation in Vivo in Bordetella Pertussis Using RIVET. Mol. Microbiol. 2005, 55, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Sarduy, L.; De Los Rios, P.; Benedetti, F.; Vobornik, D.; Dietler, G.; Kasas, S.; Longo, G. Real-time monitoring of protein conformational changes using a nano-mechanical sensor. PLoS ONE 2014, 9, e103674. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalba, M.I.; Venturelli, L.; Willaert, R.; Vela, M.E.; Yantorno, O.; Dietler, G.; Longo, G.; Kasas, S. Nanomotion Spectroscopy as a New Approach to Characterize Bacterial Virulence. Microorganisms 2021, 9, 1545. https://doi.org/10.3390/microorganisms9081545
Villalba MI, Venturelli L, Willaert R, Vela ME, Yantorno O, Dietler G, Longo G, Kasas S. Nanomotion Spectroscopy as a New Approach to Characterize Bacterial Virulence. Microorganisms. 2021; 9(8):1545. https://doi.org/10.3390/microorganisms9081545
Chicago/Turabian StyleVillalba, Maria I., Leonardo Venturelli, Ronnie Willaert, Maria E. Vela, Osvaldo Yantorno, Giovanni Dietler, Giovanni Longo, and Sandor Kasas. 2021. "Nanomotion Spectroscopy as a New Approach to Characterize Bacterial Virulence" Microorganisms 9, no. 8: 1545. https://doi.org/10.3390/microorganisms9081545
APA StyleVillalba, M. I., Venturelli, L., Willaert, R., Vela, M. E., Yantorno, O., Dietler, G., Longo, G., & Kasas, S. (2021). Nanomotion Spectroscopy as a New Approach to Characterize Bacterial Virulence. Microorganisms, 9(8), 1545. https://doi.org/10.3390/microorganisms9081545