
Fluoroquinolone-Transition Metal Complexes: A Strategy to Overcome Bacterial Resistance



Abstract
1. Introduction
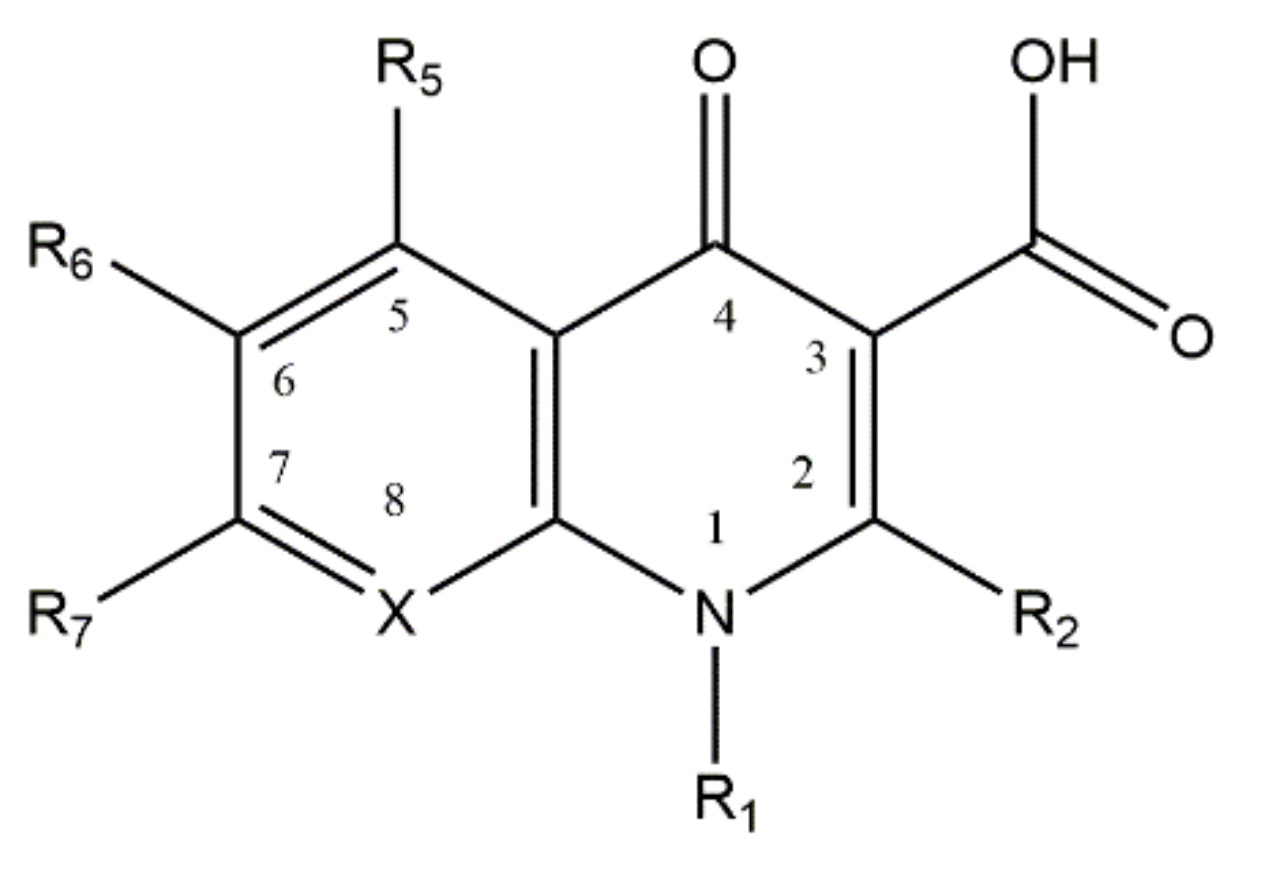
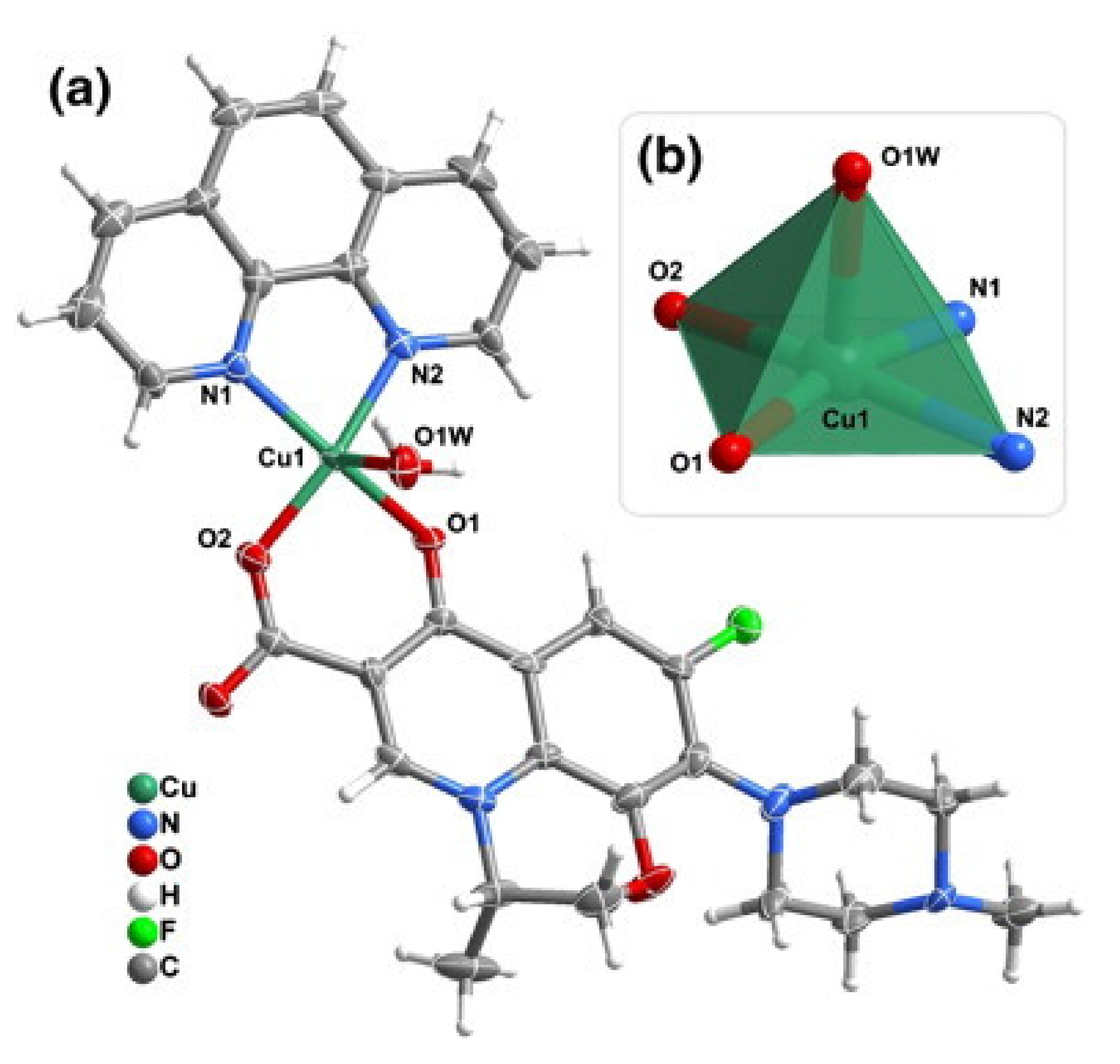
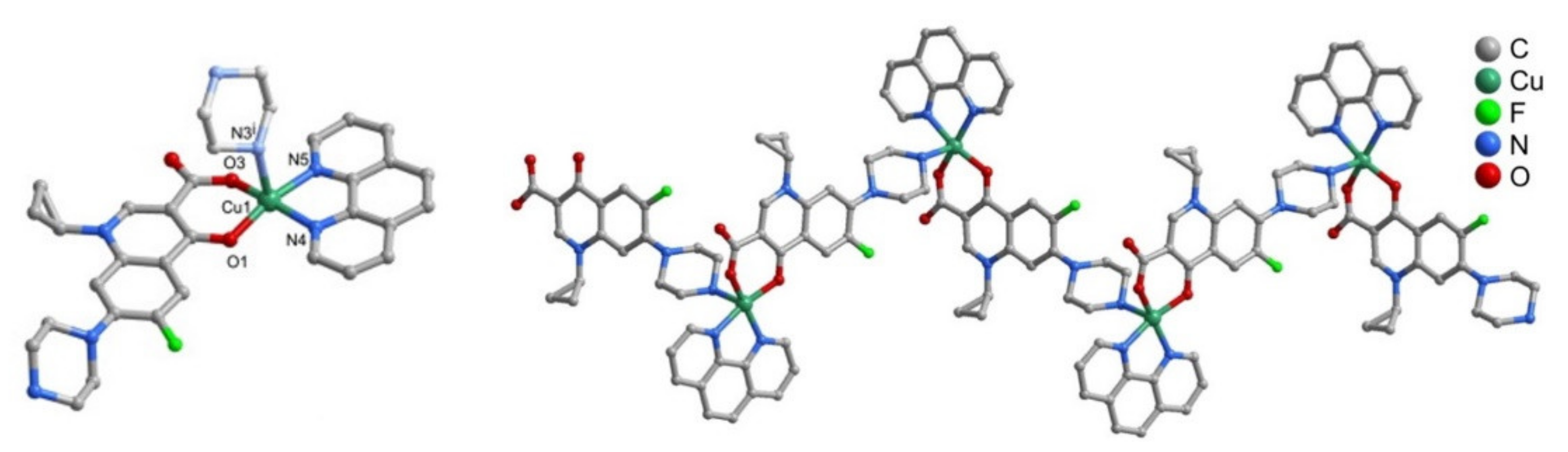
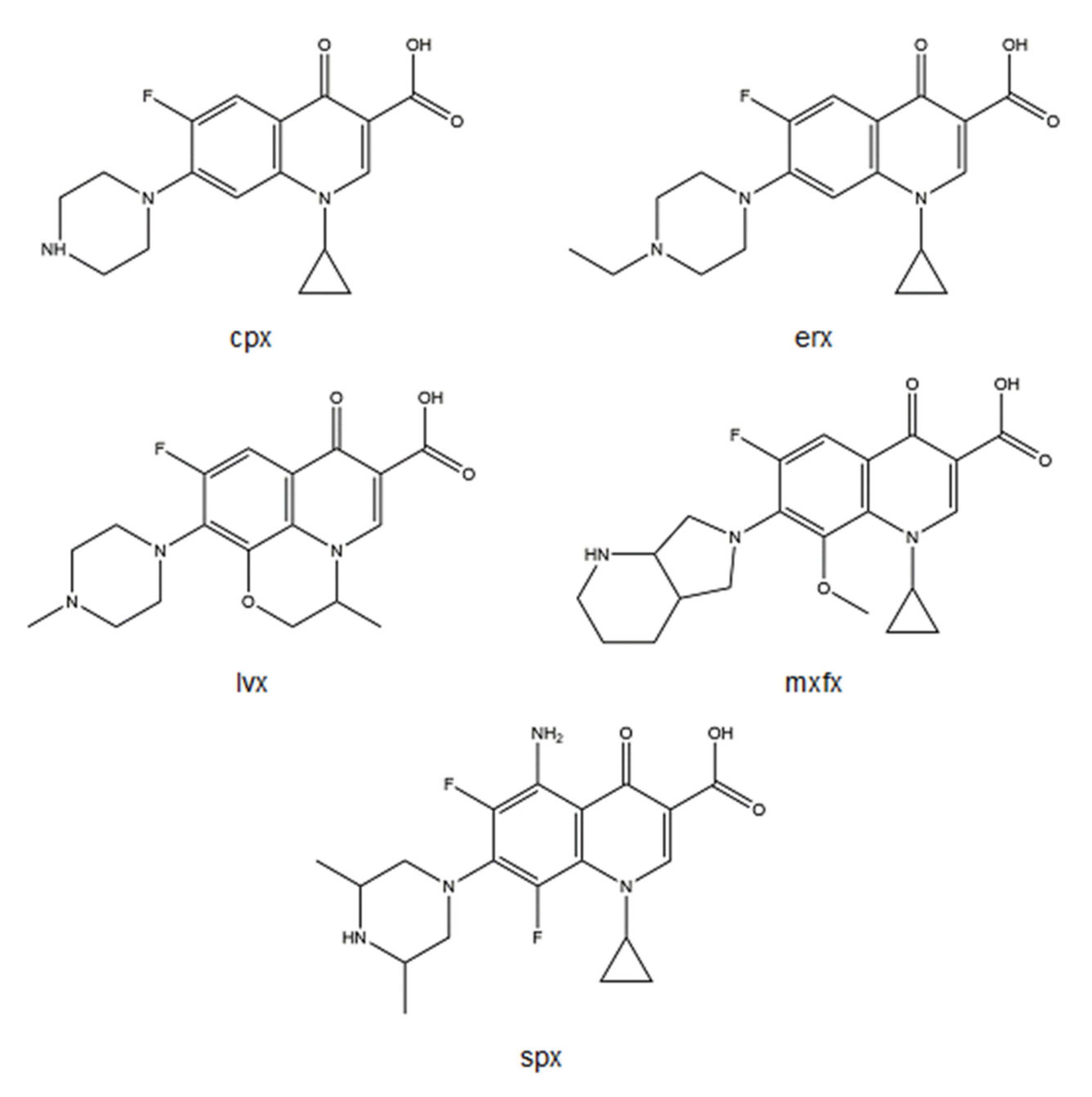
2. Fluoroquinolone Metalloantibiotics
3. Metalloantibiotic-Membrane Interactions
3.1. Partition Constants
3.2. Porins
3.2.1. Microbiological Studies
3.2.2. Biophysical Studies
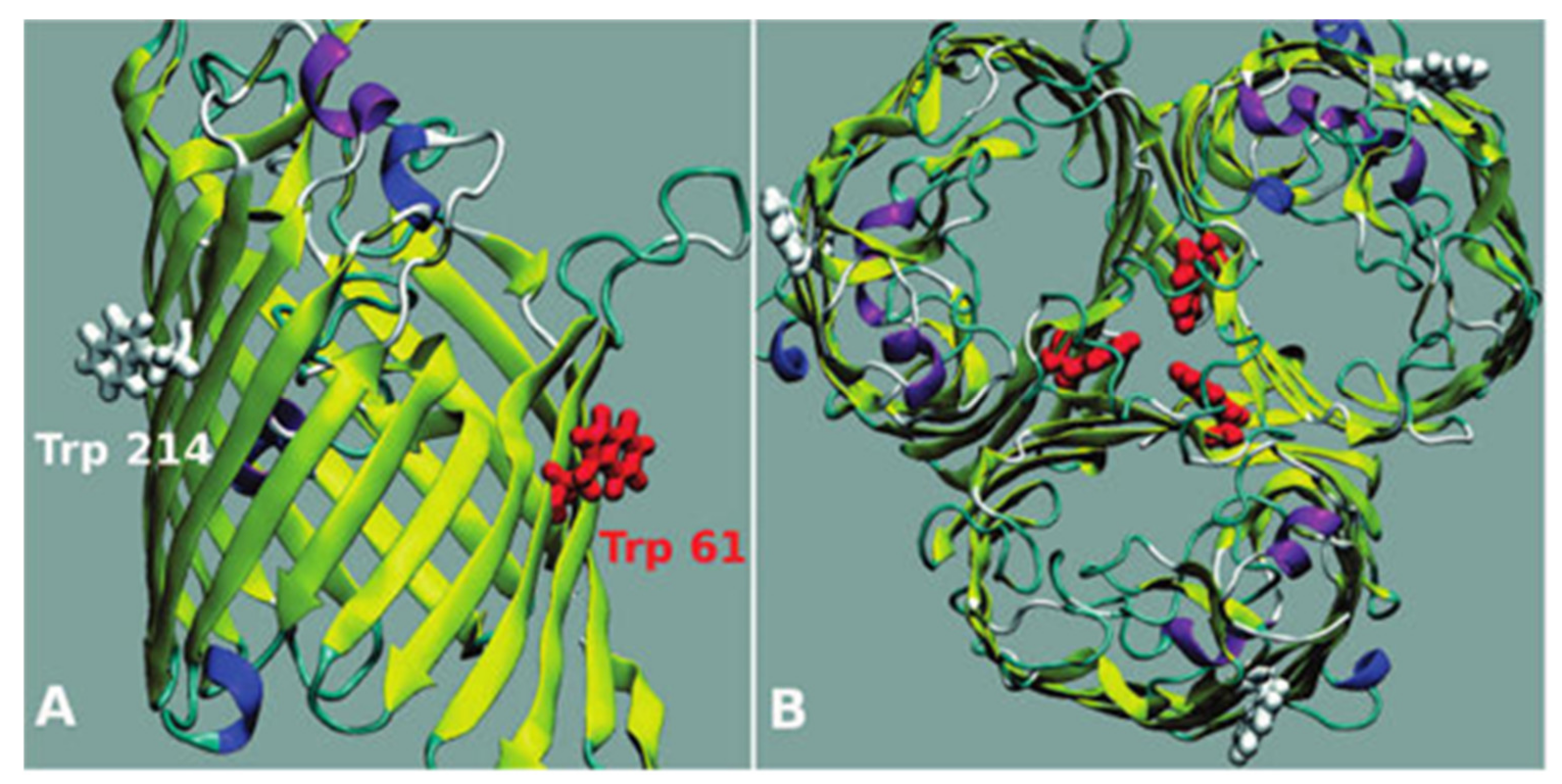
3.2.3. Computational Experiments
4. Antimicrobial Activity, Mechanism of Action, and Safety Profile of Metalloantibiotics
4.1. Antimicrobial Activity
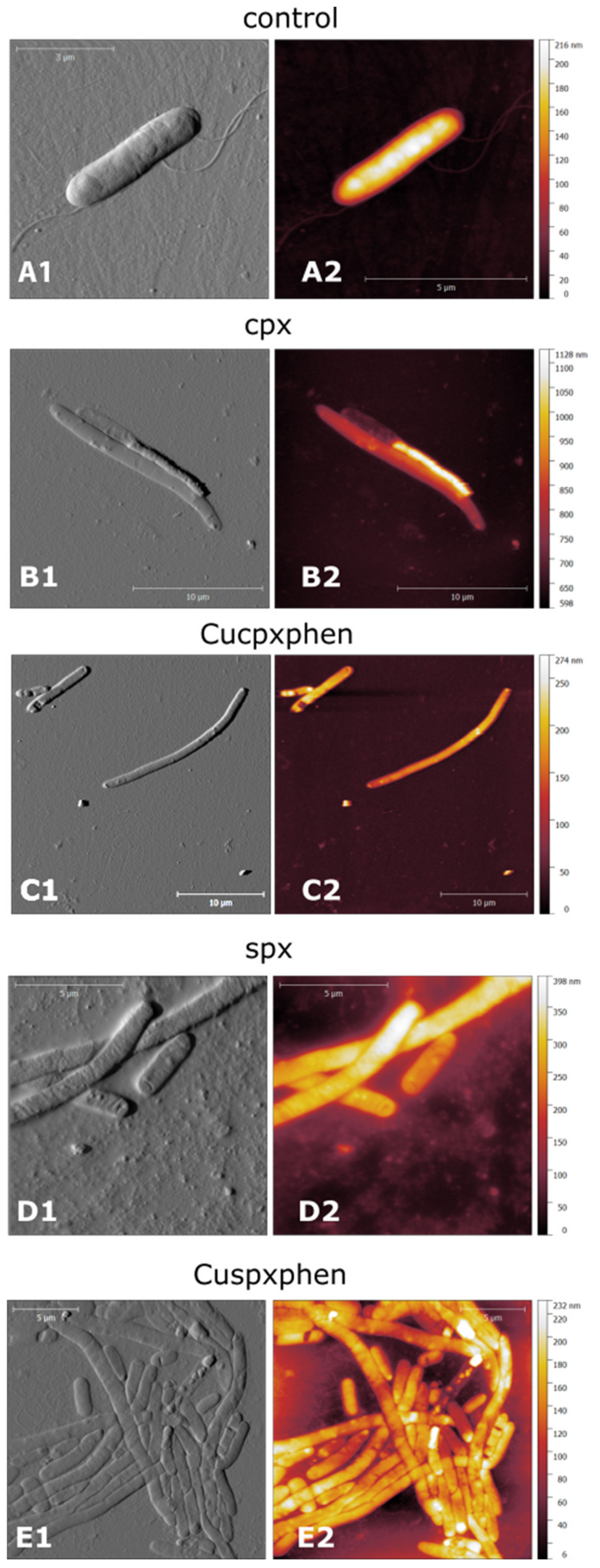
4.2. Mechanism of Action
4.3. Safety Profile
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andersson, M.I.; MacGowan, A.P. Development of the quinolones. J. Antimicrob. Chemother. 2003, 51, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, P.C.; Hunter, P.A. The fluoroquinolone antibacterials: Past, present and future perspectives. Int. J. Antimicrob. Agents 2000, 16, 5–15. [Google Scholar] [CrossRef]
- Emmerson, A.M.; Jones, A.M. The quinolones: Decades of development and use. J. Antimicrob. Chemother. 2003, 51, 13–20. [Google Scholar] [CrossRef]
- Naqvi, S.A.R.; Drlica, K. Fluoroquinolones as imaging agents for bacterial infection. Dalton Trans. 2017, 46, 14452–14460. [Google Scholar] [CrossRef]
- Andriole, V.T. The Quinolones: Past, present, and future. Clin. Infect. Dis. 2005, 41, S113–S119. [Google Scholar] [CrossRef] [PubMed]
- Suaifan, G.A.R.Y.; Mohammed, A.A.M. Fluoroquinolones structural and medicinal developments (2013–2018): Where are we now? Bioorg. Med. Chem. 2019, 27, 3005–3060. [Google Scholar] [CrossRef] [PubMed]
- Aldred, K.J.; Kerns, R.J.; Osheroff, N. Mechanism of quinolone action and resistance. Biochemistry 2014, 53, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Redgrave, L.S.; Sutton, S.B.; Webber, M.A.; Piddock, L.J.V. Fluoroquinolone resistance: Mechanisms, impact on bacteria, and role in evolutionary success. Trends Microbiol. 2014, 22, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Mustaev, A.; Malik, M.; Zhao, X.; Kurepina, N.; Luan, G.; Oppegard, L.M.; Hiasa, H.; Marks, K.R.; Kerns, R.J.; Berger, J.M.; et al. Fluoroquinolone-Gyrase-DNA complexes: Two modes of drug binding. J. Biol. Chem. 2014, 289, 12300–12312. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Wright, G.D. Molecular mechanisms of antibiotic resistance. Chem. Commun. 2011, 47, 4055–4061. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4, 2. [Google Scholar] [CrossRef]
- Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017.
- ECDC. Antimicrobial Resistance Surveillance in EUROPE 2015. Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net); European Centre for Disease Prevention and Control: Stockholm, Sweden, 2017. [Google Scholar]
- Fàbrega, A.; Madurga, S.; Giralt, E.; Vila, J. Mechanism of action of and resistance to quinolones. Microb. Biotechnol. 2009, 2, 40–61. [Google Scholar] [CrossRef]
- Kim, E.S.; Hooper, D.C. Clinical importance and epidemiology of quinolone resistance. Infect. Chemother. 2014, 46, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Gould, K.A.; Pan, X.-S.; Kerns, R.J.; Fisher, L.M. Ciprofloxacin Dimers Target Gyrase in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 2108–2115. [Google Scholar] [CrossRef]
- Bansal, S.; Tandon, V. Contribution of mutations in DNA gyrase and topoisomerase IV genes to ciprofloxacin resistance in Escherichia coli clinical isolates. Int. J. Antimicrob. Agents 2011, 37, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. MMBR 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.M.; Heinze, T.M.; Strakosha, R.; Elkins, C.A.; Sutherland, J.B. Acetylation of fluoroquinolone antimicrobial agents by an Escherichia coli strain isolated from a municipal wastewater treatment plant. J. Appl. Microbiol. 2009, 106, 564–571. [Google Scholar] [CrossRef]
- Aeschlimann, J.R. The role of multidrug efflux pumps in the antibiotic resistance of Pseudomonas aeruginosa and other Gram-negative bacteria. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2003, 23, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59–71. [Google Scholar] [CrossRef]
- Prajapati, J.D.; Kleinekathöfer, U.; Winterhalter, M. How to enter a bacterium: Bacterial porins and the permeation of antibiotics. Chem. Rev. 2021, 121, 5158–5192. [Google Scholar] [CrossRef]
- Adabi, M.; Talebi-Taher, M.; Arbabi, L.; Afshar, M.; Fathizadeh, S.; Minaeian, S.; Moghadam-Maragheh, N.; Majidpour, A. Spread of efflux pump overexpressing-mediated fluoroquinolone resistance and multidrug resistance in Pseudomonas aeruginosa by using an efflux pump inhibitor. Infect. Chemother. 2015, 47, 98–104. [Google Scholar] [CrossRef]
- Schulz, G.E. The structure of bacterial outer membrane proteins. Biochim. Biophys. Acta BBA Biomembr. 2002, 1565, 308–317. [Google Scholar] [CrossRef]
- Ziervogel, B.K.; Roux, B. The binding of antibiotics in OmpF porin. Structure 2013, 21, 76–87. [Google Scholar] [CrossRef]
- Saraiva, R.G.; Lopes, J.A.; Machado, J.; Gameiro, P.; Feio, M.J. Discrimination of single-porin Escherichia (E.) coli mutants by ATR and transmission mode FTIR spectroscopy. J. Biophotonics 2014, 7, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Doukyu, N. Contributions of mutations in acrR and marR genes to organic solvent tolerance in Escherichia coli. AMB Express 2012, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Pagès, J.-M. Structure, function and regulation of outer membrane proteins involved in drug transport in Enterobactericeae: The OmpF/C—TolC case. Open Microbiol. J. 2013, 7, 22–33. [Google Scholar] [CrossRef]
- Feio, M.J.; Sousa, I.; Ferreira, M.; Cunha-Silva, L.; Saraiva, R.G.; Queirós, C.; Alexandre, J.G.; Claro, V.; Mendes, A.; Ortiz, R.; et al. Fluoroquinolone–metal complexes: A route to counteract bacterial resistance? J. Inorg. Biochem. 2014, 138, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Turel, I. The interactions of metal ions with quinolone antibacterial agents. Coord. Chem. Rev. 2002, 232, 27–47. [Google Scholar] [CrossRef]
- Hernández-Gil, J.; Perelló, L.; Ortiz, R.; Alzuet, G.; González-Álvarez, M.; Liu-González, M. Synthesis, structure and biological properties of several binary and ternary complexes of copper(II) with ciprofloxacin and 1,10 phenanthroline. Polyhedron 2009, 28, 138–144. [Google Scholar] [CrossRef]
- Saraiva, R.; Lopes, S.; Ferreira, M.; Novais, F.; Pereira, E.; Feio, M.J.; Gameiro, P. Solution and biological behaviour of enrofloxacin metalloantibiotics: A route to counteract bacterial resistance? J. Inorg. Biochem. 2010, 104, 843–850. [Google Scholar] [CrossRef]
- Sousa, I.; Claro, V.; Pereira, J.L.; Amaral, A.L.; Cunha-Silva, L.; de Castro, B.; Feio, M.J.; Pereira, E.; Gameiro, P. Synthesis, characterization and antibacterial studies of a copper(II) levofloxacin ternary complex. J. Inorg. Biochem. 2012, 110, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, P.; Rodrigues, C.; Baptista, T.; Sousa, I.; de Castro, B. Solution studies on binary and ternary complexes of copper(II) with some fluoroquinolones and 1,10-phenanthroline: Antimicrobial activity of ternary metalloantibiotics. Int. J. Pharm. 2007, 334, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Bessa, L.J.; Sousa, C.F.; Eaton, P.; Bongiorno, D.; Stefani, S.; Campanile, F.; Gameiro, P. Fluoroquinolone metalloantibiotics: A promising approach against Methicillin-Resistant Staphylococcus aureus. Int. J. Environ. Res. Public Health 2020, 17, 3127. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Sousa, C.F.; Gameiro, P. Fluoroquinolone metalloantibiotics to bypass antimicrobial resistance mechanisms: Decreased permeation through porins. Membranes 2021, 11, 3. [Google Scholar] [CrossRef]
- Sousa, C.F.; Coimbra, J.T.S.; Ferreira, M.; Pereira-Leite, C.; Reis, S.; Ramos, M.J.; Fernandes, P.A.; Gameiro, P. Passive diffusion of ciprofloxacin and its metalloantibiotic: A computational and experimental study. J. Mol. Biol. 2021, 433, 166911. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.; Coimbra, J.; Gomes, I.; Franco, R.; Fernandes, P.A.; Gameiro, P. The binding of free and copper-complexed fluoroquinolones to OmpF porins: An experimental and molecular docking study. RSC Adv. 2017, 7, 10009–10019. [Google Scholar] [CrossRef]
- Chohan, Z.H.; Supuran, C.T.; Scozzafava, A. Metal binding and antibacterial activity of ciprofloxacin complexes. J. Enzym. Inhib. Med. Chem. 2005, 20, 303–307. [Google Scholar] [CrossRef]
- Vieira, L.M.M.; de Almeida, M.V.; Lourenço, M.C.S.; Bezerra, F.A.F.M.; Fontes, A.P.S. Synthesis and antitubercular activity of palladium and platinum complexes with fluoroquinolones. Eur. J. Med. Chem. 2009, 44, 4107–4111. [Google Scholar] [CrossRef] [PubMed]
- Osredkar, J.; Sustar, N. Copper and zinc, biological role and significance of copper/zinc imbalance. J. Clin. Toxicol. 2011, S3:001. [Google Scholar] [CrossRef]
- Cobine, P.A.; Pierrel, F.; Winge, D.R. Copper trafficking to the mitochondrion and assembly of copper metalloenzymes. Biochim. Et Biophys. Acta BBA Mol. Cell Res. 2006, 1763, 759–772. [Google Scholar] [CrossRef]
- Samanovic, M.I.; Ding, C.; Thiele, D.J.; Darwin, K.H. Copper in microbial pathogenesis: Meddling with the metal. Cell Host Microbe 2012, 11, 106–115. [Google Scholar] [CrossRef]
- Palmer, L.D.; Skaar, E.P. Transition metals and virulence in bacteria. Annu. Rev. Genet. 2016, 50, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; McDermott, P.; Walker, R. Pharmacology of the fluoroquinolones: A perspective for the use in domestic animals. Vet. J. 2006, 172, 10–28. [Google Scholar] [CrossRef]
- Vázquez, J.L.; Berlanga, M.; Merino, S.; Domènech, Ò.; Viñas, M.; Montero, M.T.; Hernández-Borrell, J. Determination by fluorimetric titration of the ionization constants of ciprofloxacin in solution and in the presence of liposomes. Photochem. Photobiol. 2001, 73, 14–19. [Google Scholar] [CrossRef]
- Sun, J.; Sakai, S.; Tauchi, Y.; Deguchi, Y.; Chen, J.; Zhang, R.; Morimoto, K. Determination of lipophilicity of two quinolone antibacterials, ciprofloxacin and grepafloxacin, in the protonation equilibrium. Eur. J. Pharm. Biopharm. 2002, 54, 51–58. [Google Scholar] [CrossRef]
- Nikaido, H.; Thanassi, D.G. Penetration of lipophilic agents with multiple protonation sites into bacterial cells: Tetracyclines and fluoroquinolones as examples. Antimicrob. Agents Chemother. 1993, 37, 1393–1399. [Google Scholar] [CrossRef] [PubMed]
- Cama, J.; Schaich, M.; Al Nahas, K.; Hernandez-Ainsa, S.; Pagliara, S.; Keyser, U.F. Direct optofluidic measurement of the lipid permeability of fluoroquinolones. Sci. Rep. 2016, 6, 32824. [Google Scholar] [CrossRef]
- Fernandes, P.; Sousa, I.; Cunha-Silva, L.; Ferreira, M.; de Castro, B.; Pereira, E.F.; Feio, M.J.; Gameiro, P. Synthesis, characterization and antibacterial studies of a copper(II) lomefloxacin ternary complex. J. Inorg. Biochem. 2014, 131, 21–29. [Google Scholar] [CrossRef]
- Wallis, S.C.; Gahan, L.R.; Charles, B.G.; Hambley, T.W.; Duckworth, P.A. Copper(II) complexes of the fluoroquinolone antimicrobial ciprofloxacin. Synthesis, X-ray structural characterization, and potentiometric study. J. Inorg. Biochem. 1996, 62, 1–16. [Google Scholar] [CrossRef]
- Macias, B.; Villa, M.V.; Sastre, M.; Castiñeiras, A.; Borras, J. Complexes of Co(II) and Zn(II) with ofloxacin. Crystal structure of [Co(oflo)2(MeOH)2]·4MeOH. J. Pharm. Sci. 2002, 91, 2416–2423. [Google Scholar] [CrossRef]
- López-Gresa, M.P.; Ortiz, R.; Perelló, L.; Latorre, J.; Liu-González, M.; García-Granda, S.; Pérez-Priede, M.; Cantón, E. Interactions of metal ions with two quinolone antimicrobial agents (cinoxacin and ciprofloxacin): Spectroscopic and X-ray structural characterization. Antibacterial studies. J. Inorg. Biochem. 2002, 92, 65–74. [Google Scholar] [CrossRef]
- Wu, G.; Wang, G.; Fu, X.; Zhu, L. Synthesis, crystal structure, stacking effect and antibacterial studies of a novel quaternary copper (II) complex with quinolone. Mol. A J. Synth. Chem. Nat. Prod. Chem. 2003, 8, 287–296. [Google Scholar] [CrossRef]
- Macías, B.; Villa, M.a.V.; Rubio, I.; Castiñeiras, A.; Borrás, J.n. Complexes of Ni(II) and Cu(II) with ofloxacin: Crystal structure of a new Cu(II) ofloxacin complex. J. Inorg. Biochem. 2001, 84, 163–170. [Google Scholar] [CrossRef]
- Sigman, D.S.; Graham, D.R.; D’Aurora, V.; Stern, A.M. Oxygen-dependent cleavage of DNA by the 1,10-phenanthroline. cuprous complex. Inhibition of Escherichia coli DNA polymerase I. J. Biol. Chem. 1979, 254, 12269–12272. [Google Scholar] [CrossRef]
- Chen, C.-h.B.; Milne, L.; Landgraf, R.; Perrin, D.M.; Sigman, D.S. Artificial nucleases. ChemBioChem 2001, 2, 735–740. [Google Scholar] [CrossRef]
- Chen, C.H.; Sigman, D.S. Nuclease activity of 1,10-phenanthroline-copper: Sequence-specific targeting. Proc. Natl. Acad. Sci. USA 1986, 83, 7147–7151. [Google Scholar] [CrossRef]
- Hirohama, T.; Kuranuki, Y.; Ebina, E.; Sugizaki, T.; Arii, H.; Chikira, M.; Tamil Selvi, P.; Palaniandavar, M. Copper(II) complexes of 1,10-phenanthroline-derived ligands: Studies on DNA binding properties and nuclease activity. J. Inorg. Biochem. 2005, 99, 1205–1219. [Google Scholar] [CrossRef]
- Lu, L.-P.; Zhu, M.-L.; Yang, P. Crystal structure and nuclease activity of mono(1,10-phenanthroline) copper complex. J. Inorg. Biochem. 2003, 95, 31–36. [Google Scholar] [CrossRef]
- Fernández-Botello, A.; Holý, A.n.; Moreno, V.; Sigel, H. Intramolecular stacking interactions in ternary copper(II) complexes formed by a heteroaromatic amine and 9-[2-(2-phosphonoethoxy)ethyl]adenine, a relative of the antiviral nucleotide analogue 9-[2-(phosphonomethoxy)ethyl]adenine. J. Inorg. Biochem. 2004, 98, 2114–2124. [Google Scholar] [CrossRef] [PubMed]
- Lüth, M.S.; Kapinos, L.E.; Song, B.; Lippert, B.; Sigel, H. Extent of intramolecular stacking interactions in the mixed-ligand complexes formed in aqueous solution by copper(II), 2,2′-bipyridine or 1,10-phenanthroline and 2′-deoxyguanosine 5′-monophosphate. J. Chem. Soc. Dalton Trans. 1999, 3, 357–366. [Google Scholar] [CrossRef]
- Sigel, H. Ternary complexes in solution—XIX Relation between ligand basicity and complex stability. A method for the estimation of stability constants. J. Inorg. Nucl. Chem. 1975, 37, 507–509. [Google Scholar] [CrossRef]
- Ruíz, P.; Ortiz, R.; Perelló, L.; Alzuet, G.; González-Álvarez, M.; Liu-González, M.; Sanz-Ruíz, F. Synthesis, structure, and nuclease properties of several binary and ternary complexes of copper(II) with norfloxacin and 1,10 phenantroline. J. Inorg. Biochem. 2007, 101, 831–840. [Google Scholar] [CrossRef]
- Efthimiadou, E.K.; Sanakis, Y.; Katsarou, M.; Raptopoulou, C.P.; Karaliota, A.; Katsaros, N.; Psomas, G. Neutral and cationic mononuclear copper(II) complexes with enrofloxacin: Structure and biological activity. J. Inorg. Biochem. 2006, 100, 1378–1388. [Google Scholar] [CrossRef]
- Matos, C.; Moutinho, C.; Lobão, P. Liposomes as a model for the biological membrane: Studies on daunorubicin bilayer interaction. J. Membr. Biol. 2012, 245, 69–75. [Google Scholar] [CrossRef]
- Esteves, F.; Moutinho, C.; Matos, C. Correlation between octanol/water and liposome/water distribution coefficients and drug absorption of a set of pharmacologically active compounds. J. Liposome Res. 2013, 23, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Gameiro, P.; Prieto, M.; de Castro, B. Interaction of rifampicin and isoniazid with large unilamellar liposomes: Spectroscopic location studies. Biochim. Biophys. Acta BBA Gen. Subj. 2003, 1620, 151–159. [Google Scholar] [CrossRef]
- Omran, A.A.; Kitamura, K.; Takegami, S.; Kume, M.; Yoshida, M.; El-Sayed, A.-A.Y.; Mohamed, M.H.; Abdel-Mottaleb, M. 19F NMR spectrometric determination of the partition coefficients of some fluorinated psychotropic drugs between phosphatidylcholine bilayer vesicles and water. J. Pharm. Biomed. Anal. 2002, 30, 1087–1092. [Google Scholar] [CrossRef]
- Sousa, C.F.; Ferreira, M.; Abreu, B.; Medforth, C.J.; Gameiro, P. Interactions of a non-fluorescent fluoroquinolone with biological membrane models: A multi-technique approach. Int. J. Pharm. 2015, 495, 761–770. [Google Scholar] [CrossRef]
- Rodrigues, C.; Gameiro, P.; Reis, S.; Lima, J.L.F.C.; de Castro, B. Derivative spectrophotometry as a tool for the determination of drug partition coefficients in water/dimyristoyl-l-α-phosphatidylglycerol (DMPG) liposomes. Biophys. Chem. 2001, 94, 97–106. [Google Scholar] [CrossRef]
- Rodrigues, C.; Gameiro, P.; Reis, S.; Lima, J.L.F.C.; de Castro, B. Spectrophotometric determination of drug partition coefficients in dimyristoyl-l-α-phosphatidylcholine/water: A comparative study using phase separation and liposome suspensions. Anal. Chim. Acta 2001, 428, 103–109. [Google Scholar] [CrossRef]
- Rodrigues, C.; Gameiro, P.; Reis, S.; Lima, J.L.F.C.; de Castro, B. Interaction of grepafloxacin with large unilamellar liposomes: Partition and fluorescence studies reveal the importance of charge interactions. Langmuir 2002, 18, 10231–10236. [Google Scholar] [CrossRef]
- Ferreira, M.; Gameiro, P. Ciprofloxacin metalloantibiotic: An effective antibiotic with an influx route strongly dependent on lipid interaction? J. Membr. Biol. 2015, 248, 125–136. [Google Scholar] [CrossRef] [PubMed]
- De Castro, B.; Gameiro, P.; Lima, J.L.F.C.; Matos, C.; Reis, S. Location and partition coefficients of anti-inflammatory drugs in EPC liposomes. A fluorescence quenching study using n-(9-anthroyloxy)-stearic probes. Colloids Surf. A Physicochem. Eng. Asp. 2001, 190, 205–212. [Google Scholar] [CrossRef]
- Lopes, S.C.; Ribeiro, C.; Gameiro, P. A new approach to counteract bacteria resistance: A comparative study between moxifloxacin and a new moxifloxacin derivative in different model systems of bacterial membrane. Chem. Biol. Drug Des. 2013, 81, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.C.; Prieto, M.; Castanho, M.A.R.B. Quantifying molecular partition into model systems of biomembranes: An emphasis on optical spectroscopic methods. Biochim. Biophys. Acta BBA Biomembr. 2003, 1612, 123–135. [Google Scholar] [CrossRef]
- Coimbra, J.T.S.; Fernandes, P.A.; Ramos, M.J. Revisiting partition in hydrated bilayer systems. J. Chem. Theory Comput. 2017, 13, 2290–2299. [Google Scholar] [CrossRef] [PubMed]
- Orsi, M.; Essex, J.W. Permeability of drugs and hormones through a lipid bilayer: Insights from dual-resolution molecular dynamics. Soft Matter 2010, 6, 3797–3808. [Google Scholar] [CrossRef]
- Atkovska, K.; Klingler, J.; Oberwinkler, J.; Keller, S.; Hub, J.S. Rationalizing steroid interactions with lipid membranes: Conformations, partitioning, and kinetics. ACS Cent. Sci. 2018, 4, 1155–1165. [Google Scholar] [CrossRef]
- López-Lara, I.M.; Geiger, O. Novel pathway for phosphatidylcholine biosynthesis in bacteria associated with eukaryotes. J. Biotechnol. 2001, 91, 211–221. [Google Scholar] [CrossRef]
- Strahl, H.; Errington, J. Bacterial membranes: Structure, domains, and Ffunction. Annu. Rev. Microbiol. 2017, 71, 519–538. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Epand, R.F. Lipid domains in bacterial membranes and the action of antimicrobial agents. Biochim. Biophys. Acta BBA Biomembr. 2009, 1788, 289–294. [Google Scholar] [CrossRef]
- Lopes, S.; Neves, C.S.; Eaton, P.; Gameiro, P. Cardiolipin, a key component to mimic the E. coli bacterial membrane in model systems revealed by dynamic light scattering and steady-state fluorescence anisotropy. Anal. Bioanal. Chem. 2010, 398, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.S.; Mohamed, Z.J.; Artim, C.M.; Thornlow, D.N.; Hassler, J.F.; Rigoglioso, V.P.; Daniel, S.; Alabi, C.A. Antibacterial isoamphipathic oligomers highlight the importance of multimeric lipid aggregation for antibacterial potency. Commun. Biol. 2018, 1, 220. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.; Lopes, S.C.; Gameiro, P. New insights into the translocation route of enrofloxacin and its metalloantibiotics. J. Membr. Biol. 2011, 241, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Neves, P.; Berkane, E.; Gameiro, P.; Winterhalter, M.; de Castro, B. Interaction between quinolones antibiotics and bacterial outer membrane porin OmpF. Biophys. Chem. 2005, 113, 123–128. [Google Scholar] [CrossRef]
- Choi, U.; Lee, C.-R. Distinct roles of uuter membrane porins in antibiotic resistance and membrane integrity in Escherichia coli. Front. Microbiol. 2019, 10, 953. [Google Scholar] [CrossRef]
- Mach, T.; Neves, P.; Spiga, E.; Weingart, H.; Winterhalter, M.; Ruggerone, P.; Ceccarelli, M.; Gameiro, P. Facilitated permeation of antibiotics across membrane channels—Interaction of the quinolone moxifloxacin with the OmpF Channel. J. Am. Chem. Soc. 2008, 130, 13301–13309. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta BBA Biomembr. 2016, 1858, 980–987. [Google Scholar] [CrossRef]
- Graef, F.; Vukosavljevic, B.; Michel, J.-P.; Wirth, M.; Ries, O.; De Rossi, C.; Windbergs, M.; Rosilio, V.; Ducho, C.; Gordon, S.; et al. The bacterial cell envelope as delimiter of anti-infective bioavailability—An in vitro permeation model of the Gram-negative bacterial inner membrane. J. Control. Release 2016, 243, 214–224. [Google Scholar] [CrossRef]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta 2009, 1794, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. [Google Scholar] [CrossRef]
- Rollauer, S.E.; Sooreshjani, M.A.; Noinaj, N.; Buchanan, S.K. Outer membrane protein biogenesis in Gram-negative bacteria. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20150023. [Google Scholar] [CrossRef]
- Ferreira, A.R.; Teixeira, C.; Sousa, C.F.; Bessa, L.J.; Gomes, P.; Gameiro, P. How insertion of a single tryptophan in the N-terminus of a cecropin A-melittin hybrid peptide changes its antimicrobial and biophysical profile. Membranes 2021, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.T.; Lee, M.H.; Weng, C.F.; Leong, M.K. In silico prediction of PAMPA effective permeability using a two-QSAR approach. Int. J. Mol. Sci. 2019, 20, 3170. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Hossain, K.R.; Chen, R.; Ho, K.K.K.; Kuppusamy, R.; Clarke, R.J.; Kumar, N.; Willcox, M.D.P. Mechanism of action of surface immobilized antimicrobial peptides against Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 3053. [Google Scholar] [CrossRef]
- Prochnow, H.; Fetz, V.; Hotop, S.K.; Garcia-Rivera, M.A.; Heumann, A.; Bronstrup, M. Subcellular quantification of uptake in Gram-negative bacteria. Anal. Chem. 2019, 91, 1863–1872. [Google Scholar] [CrossRef]
- Cama, J.; Voliotis, M.; Metz, J.; Smith, A.; Iannucci, J.; Keyser, U.F.; Tsaneva-Atanasova, K.; Pagliara, S. Single-cell microfluidics facilitates the rapid quantification of antibiotic accumulation in Gram-negative bacteria. Lab A Chip 2020, 20, 2765–2775. [Google Scholar] [CrossRef]
- Vergalli, J.; Dumont, E.; Cinquin, B.; Maigre, L.; Pajovic, J.; Bacqué, E.; Mourez, M.; Réfrégiers, M.; Pagès, J.-M. Fluoroquinolone structure and translocation flux across bacterial membrane. Sci. Rep. 2017, 7, 9821. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.J.; Foulds, J. Escherichia coli K-12 tolF mutants: Alterations in protein composition of the outer membrane. J. Bacteriol. 1977, 130, 781–786. [Google Scholar] [CrossRef]
- Foulds, J.; Chai, T.J. New major outer membrane proteins found in an Escherichia coli tolF mutant resistant to bacteriophage TuIb. J. Bacteriol. 1978, 133, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, K.R.; Hajjar, E.; Mach, T.; Lovelle, M.; Kumar, A.; Sousa, I.; Spiga, E.; Weingart, H.; Gameiro, P.; Winterhalter, M.; et al. Molecular basis of enrofloxacin translocation through OmpF, an outer membrane channel of Escherichia coli—When binding does not imply translocation. J. Phys. Chem. B 2010, 114, 5170–5179. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, F.; Neves, P.; Gameiro, P.; Loura, L.M.S.; Prieto, M. Ciprofloxacin interactions with bacterial protein OmpF: Modelling of FRET from a multi-tryptophan protein trimer. Biochim. Biophys. Acta BBA Biomembr. 2007, 1768, 2822–2830. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Neves, P.; Sousa, I.; Winterhalter, M.; Gameiro, P. Fluorescence quenching as a tool to investigate quinolone antibiotic interactions with bacterial protein OmpF. J. Membr. Biol. 2009, 227, 133–140. [Google Scholar] [CrossRef]
- Talan, D.A.; Takhar, S.S.; Krishnadasan, A.; Abrahamian, F.M.; Mower, W.R.; Moran, G.J.; Group, E.M.I.N.S. Fluoroquinolone-resistant and Extended-Spectrum β-Lactamase–Producing Escherichia coli infections in patients with pyelonephritis, United States. Emerg. Infect. Dis. 2016, 22, 1594–1603. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Jeon, S.-M.; Kim, H.; Lim, N.; Park, M.-S.; Kim, S.-H. Resistance to fluoroquinolone by a combination of efflux and target site mutations in Enteroaggregative Escherichia coli isolated in Korea. Osong Public Health Res. Perspect. 2012, 3, 239–244. [Google Scholar] [CrossRef]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant Pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef]
- Yayan, J.; Ghebremedhin, B.; Rasche, K. Antibiotic resistance of Pseudomonas aeruginosa in pneumonia at a single university hospital center in Germany over a 10-year period. PLoS ONE 2015, 10, e0139836. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Campanile, F.; Bongiorno, D.; Perez, M.; Mongelli, G.; Sessa, L.; Benvenuto, S.; Gona, F.; Varaldo, P.E.; Stefani, S. Epidemiology of Staphylococcus aureus in Italy: First nationwide survey, 2012. J. Glob. Antimicrob. Resist. 2015, 3, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Nishino, K.; Roberts, M.C.; Tolmasky, M.; Aminov, R.I.; Zhang, L. Mechanisms of antibiotic resistance. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Sivagurunathan, N.; Krishnan, S.; Rao, J.V.; Nagappa, A.N.; Subrahmanyam, V.M.; Vanathi, B.M. Synergy of gatifloxacin with cefoperazone and cefoperazone–sulbactam against resistant strains of Pseudomonas aeruginosa. J. Med. Microbiol. 2008, 57, 1514–1517. [Google Scholar] [CrossRef]
- Fish, D.N.; Choi, M.K.; Jung, R. Synergic activity of cephalosporins plus fluoroquinolones against Pseudomonas aeruginosa with resistance to one or both drugs. J. Antimicrob. Chemother. 2002, 50, 1045–1049. [Google Scholar] [CrossRef]
- Drago, L.; Nicola, L.; Rodighiero, V.; Larosa, M.; Mattina, R.; De Vecchi, E. Comparative evaluation of synergy of combinations of β-lactams with fluoroquinolones or a macrolide in Streptococcus pneumoniae. J. Antimicrob. Chemother. 2011, 66, 845–849. [Google Scholar] [CrossRef]
- Louie, A.; Liu, W.; VanGuilder, M.; Neely, M.N.; Schumitzky, A.; Jelliffe, R.; Fikes, S.; Kurhanewicz, S.; Robbins, N.; Brown, D.; et al. Combination treatment with meropenem plus levofloxacin is synergistic against Pseudomonas aeruginosa infection in a murine model of pneumonia. J. Infect. Dis. 2015, 211, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulou, T.; Meletiadis, J.; Sein, T.; Papaioannidou, P.; Tsiouris, I.; Roilides, E.; Walsh, T.J. Comparative pharmacodynamic interaction analysis between ciprofloxacin, moxifloxacin and levofloxacin and antifungal agents against Candida albicans and Aspergillus fumigatus. J. Antimicrob. Chemother. 2009, 63, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.H.; Brown, G.H.; Peterson, M.L.; Rotschafer, J.C. Application of fluoroquinolone pharmacodynamics. J. Antimicrob. Chemother. 2000, 46, 669–683. [Google Scholar] [CrossRef] [PubMed]
- Akinremi, C.A.; Obaleye, J.A.; Amolegbe, S.A.; Adediji, J.F.; Bamigboye, M.O. Biological activities of some fluoroquinolones-metal complexes. Int. J. Med. Biomed. Res. 2012, 1, 24–34. [Google Scholar] [CrossRef]
- Diver, J.M.; Wise, R. Morphological and biochemical changes in Escherichia coli after exposure to ciprofloxacin. J. Antimicrob. Chemother. 1986, 18 (Suppl. D), 31–41. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Lourenço, O.; Queiroz, J.A.; Domingues, F.C. Bacteriostatic versus bactericidal activity of ciprofloxacin in Escherichia coli assessed by flow cytometry using a novel far-red dye. J. Antibiot. 2011, 64, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; O’Driscoll, N.H.; Lamb, A.J. Morphological and ultrastructural changes in bacterial cells as an indicator of antibacterial mechanism of action. Cell. Mol. Life Sci. 2016, 73, 4471–4492. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Tsurumaki, Y.; Takei, M.; Hosaka, M.; Oomori, Y. In vitro method for prediction of the phototoxic potentials of fluoroquinolones. Toxicol. Vitr. 2001, 15, 721–727. [Google Scholar] [CrossRef]
- Hayashi, N.; Nakata, Y.; Yazaki, A. New findings on the structure-phototoxicity relationship and photostability of fluoroquinolones with various substituents at position 1. Antimicrob. Agents Chemother. 2004, 48, 799–803. [Google Scholar] [CrossRef]
- Stahlmann, R. Clinical toxicological aspects of fluoroquinolones. Toxicol. Lett. 2002, 127, 269–277. [Google Scholar] [CrossRef]
- Baik, S.; Lau, J.; Huser, V.; McDonald, C.J. Association between tendon ruptures and use of fluoroquinolone, and other oral antibiotics: A 10-year retrospective study of 1 million US senior Medicare beneficiaries. BMJ Open 2020, 10, e034844. [Google Scholar] [CrossRef]
- Chinen, T.; Sasabuchi, Y.; Matsui, H.; Yasunaga, H. Association between third-generation fluoroquinolones and Achilles tendon rupture: A self-controlled case series analysis. Ann. Fam. Med. 2021, 19, 212–216. [Google Scholar] [CrossRef]
- Stephenson, A.L.; Wu, W.; Cortes, D.; Rochon, P.A. Tendon injury and fluoroquinolone use: A systematic review. Drug Saf. 2013, 36, 709–721. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FQ | MHL Log β1100 | ML Log β11–10 | M(HL)2 Log β1200 | ML2 Log β12–20 | M(HL)A Log β1101 | MLA Log β11–11 |
|---|---|---|---|---|---|---|
| cpx [30] | 5.48 ± 0.03 | −2.49 ± 0.05 | − | −6.13 ± 0.04 | 17.96 ± 0.05 | 8.42 ± 0.04 |
| erx [33] | 6.39 ± 0.03 | −0.42 ± 0.04 | 10.97 ± 0.04 | −3.32 ± 0.06 | 16.54 ± 0.05 | 9.21 ± 0.04 |
| lvx [34] | 6.22 ± 0.04 | −1.02 ± 0.08 | 10.92 ± 0.05 | −4.52 ± 0.07 | 16.53 ± 0.04 | 9.30 ± 0.09 |
| mxfx [30] | 6.01 ± 0.04 | −1.32 ± 0.08 | 12.24 ± 0.07 | −4.89 ± 0.04 | 15.95 ± 0.04 | 7.84 ± 0.09 |
| spx [30] | 6.09 ± 0.08 | − | 12.58 ± 0.08 | − | 16.46 ± 0.09 | 8.93 ± 0.09 |
| ofx [35] | 6.24 ± 0.04 | − | 11.20 ± 0.04 | − | 16.69 ± 0.03 | 9.21 ± 0.04 |
| nfx [35] | 6.95 ± 0.03 | − | 12.70 ± 0.04 | − | 17.56 ± 0.05 | 9.55 ± 0.04 |
| lmx [51] | 6.04 ± 0.01 | −0.98 ± 0.03 | 10.51 ± 0.15 | −5.48 ± 0.15 | 17.35 ± 0.07 | 8.45 ± 0.05 |
| Metalloantibiotic and its Speciation | Membrane Mimetic System | Log 𝑲𝒑 | Technique |
|---|---|---|---|
| Cucpxphen 97% di-cationic; 3% mono-cationic | DMPC | 4.15 ± 0.11 [75] | Fluorescence ST |
| POPC | 3.2 ± 0.2 [38] | Fluorescence ST | |
| POPG | 3.38 ± 0.09 [38] | Fluorescence ST | |
| POPG | 3.69 ± 0.05 [38] | UV-vis | |
| POPC | −0.25–1.18 [38] | MD simulations | |
| POPG | 1.99–2.47 [38] | MD simulations | |
| POPE:POPG (75:25) | 4.94 ± 0.02 [75] | Fluorescence ST | |
| POPE:POPG (75:25) | 4.45 ± 0.02 [75] | Fluorescence TR | |
| POPE:POPG:CL (67:23:10) | 4.50 ± 0.03 [75] | Fluorescence ST | |
| POPE:POPG:CL (67:23:10) | 4.15 ± 0.01 [75] | Fluorescence TR | |
| E. coli total lipid extract | 5.32 ± 0.09 [75] | Fluorescence ST | |
| E. coli total lipid extract | 4.65 ± 0.03 [75] | Fluorescence TR | |
| Cuerxphen 42.7% di-cationic; 50.1% mono-cationic | DMPC | 3.40 ± 0.03 a | Fluorescence ST |
| POPE:POPG (75:25) | 4.49 ± 0.02 [87] | Fluorescence ST | |
| POPE:POPG:CL (67:23:10) | 4.67 ± 0.01 [87] | Fluorescence ST | |
| E. coli polar lipid extract | 5.18 ± 0.01 [87] | Fluorescence ST | |
| E. coli total lipid extract | 4.85 ± 0.02 [87] | Fluorescence ST | |
| Culvxphen 38.5% di-cationic; 57% mono-cationic | DMPC | 4.36 ± 0.03 a | Fluorescence ST |
| POPE:POPG:CL (67:23:10) | 4.33 ± 0.10 a | Fluorescence ST | |
| E. coli total lipid extract | 4.26 ± 0.04 a | Fluorescence ST | |
| Cumxfxphen 82.7% di-cationic; 16.1% mono-cationic | DMPC | N.d. [77] | Fluorescence ST |
| POPE:POPG (75:25) | 4.22 ± 0.10 [77] | Fluorescence ST | |
| POPE:POPG:CL (67:23:10) | 4.38 ± 0.01 [77] | Fluorescence ST | |
| E. coli polar lipid extract | 4.39 ± 0.02 [77] | Fluorescence ST | |
| E. coli total lipid extract | 4.75 ± 0.01 [77] | Fluorescence ST | |
| Cuspxphen 52.6% di-cationic; 38.1% mono-cationic | POPE:POPG (75:25) | 5.05 ± 0.08 [71] | Fluorescence ST |
| POPE:POPG (75:25) | 5.04 ± 0.04 [71] | Fluorescence TR | |
| E. coli total lipid extract | 4.87 ± 0.08 [71] | Fluorescence ST | |
| E. coli total lipid extract | 5.24 ± 0.01 [71] | Fluorescence TR |
| Metalloantibiotic | ΔΔG binding/Kcal Mol−1 | |||
|---|---|---|---|---|
| Rigid Docking Protocol | Flexible Docking Protocol | |||
| 4KRA | 2OMF | 4KRA | 2OMF | |
| Cucpxphen | 1.2 | −0.01 | 2.6 | 2.6 |
| Cuspxphen | 1.0 | 0.3 | 2.9 | 3.1 |
| Metalloantibiotic | MIC Value/μmol dm−3 (μg mL−1) | |||
|---|---|---|---|---|
| E. coli ATCC 25922 | P. aeruginosa ATCC 27853 [36] | S. aureus ATCC 25923 [36] | S. aureus ATCC 29213 [36] | |
| Cucpxphen | 0.011 [36]–0.026 [30] (0.008–0.016) | 0.17–0.35 (0.12–0.25) | 0.35–0.71 (0.25–0.5) | 1.41 (1) |
| Cuerxphen | 0.022 [36]–0.027 [33] (0.015) | 2.97 (2) | 0.37–0.74 (0.25–0.5) | 0.18–0.37 (0.12–0.25) |
| Culvxphen | 0.021 [36]–0.07 [34] (0.015–0.030) | 1.39 (1) | 0.35–0.69 (0.25–0.5) | 0.35 (0.25) |
| Cumxfxphen | 0.018 [30]–0.038 [36] (0.015–0.030) | 2.54 (2) | 0.08–0.15 (0.06–0.12) | 0.08–0.15 (0.06–0.12) |
| Cuspxphen | 0.005 [36]–0.020 [30] (0.004–0.016) | 0.65 (0.5) | 0.32 (0.25) | 0.08–0.16 (0.06–0.12) |
| Cuofxphen | 0.050 [35]–0.083 [30] (0.03) | − | − | − |
| Cunfxphen | 0.107 [35]–0.185 [30] (0.06) | − | − | − |
| Compound | MIC Value/μmol dm−3 (μg mL−1) | |||||
|---|---|---|---|---|---|---|
| E. coli HSJ Ec002 | E. coli HSJ Ec003 | MRSA Sa1-SA3 | MRSA Sa3-SA3 | MRSA 19/35 | MRSA 26/01 | |
| cpx | 386.3 (128) | 193.2 (64) | 386.3 (128) | 386.3–772.6 (128–256) | ≥3090.5 (≥1024) | 1545.2 (512) |
| Cucpxphen | 93.0 (64) | 45.1–90.3 (32–64) | 90.3 (64) | 90.3 (64) | 180.5 (128) | 180.5 (128) |
| erx | 178.1 (64) | 178.1 (64) | 22.3 (8) | 44.5–89.0 (16–32) | 178.1 (64) | 712.3 (256) |
| Cuerxphen | 95.0 (64) | 95.0 (64) | 95.0 (64) | 95.0 (64) | 95.0 (64) | 95.0 (64) |
| lvx | 22.1 (8) | 88.6 (32) | 708.4 (256) | 44.3 (16) | 177.1 (64) | 1416.8 (512) |
| Culvxphen | 44.4 (32) | 44.4–88.7 (32–64) | 88.7 (64) | 88.7–177.5 (64–128) | 88.7 (64) | 88.7 (64) |
| mxfx | 18.3 (8) | 18.3 (8) | 18.3 (8) | 292.3 (128) | 18.3 (8) | 73.1 (32) |
| Cumxfxphen | 20.3 (16) | 40.6 (32) | 10.2 (8) | 10.2 (8) | 20.3 (16) | 40.6 (32) |
| spx | 40.8 (16) | 81.6 (32) | 652.4 (256) | 20.4 (8) | 163.1 (64) | 652.4 (256) |
| Cuspxphen | 41.6 (32) | 83.1 (64) | 83.1–166.2 (64–128) | 41.6 (32) | 41.6 (32) | 83.1 (64) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, M.; Gameiro, P. Fluoroquinolone-Transition Metal Complexes: A Strategy to Overcome Bacterial Resistance. Microorganisms 2021, 9, 1506. https://doi.org/10.3390/microorganisms9071506
Ferreira M, Gameiro P. Fluoroquinolone-Transition Metal Complexes: A Strategy to Overcome Bacterial Resistance. Microorganisms. 2021; 9(7):1506. https://doi.org/10.3390/microorganisms9071506
Chicago/Turabian StyleFerreira, Mariana, and Paula Gameiro. 2021. "Fluoroquinolone-Transition Metal Complexes: A Strategy to Overcome Bacterial Resistance" Microorganisms 9, no. 7: 1506. https://doi.org/10.3390/microorganisms9071506
APA StyleFerreira, M., & Gameiro, P. (2021). Fluoroquinolone-Transition Metal Complexes: A Strategy to Overcome Bacterial Resistance. Microorganisms, 9(7), 1506. https://doi.org/10.3390/microorganisms9071506