
Ticks and Tick-Borne Pathogens Associated with Dromedary Camels (Camelus dromedarius) in Northern Kenya

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
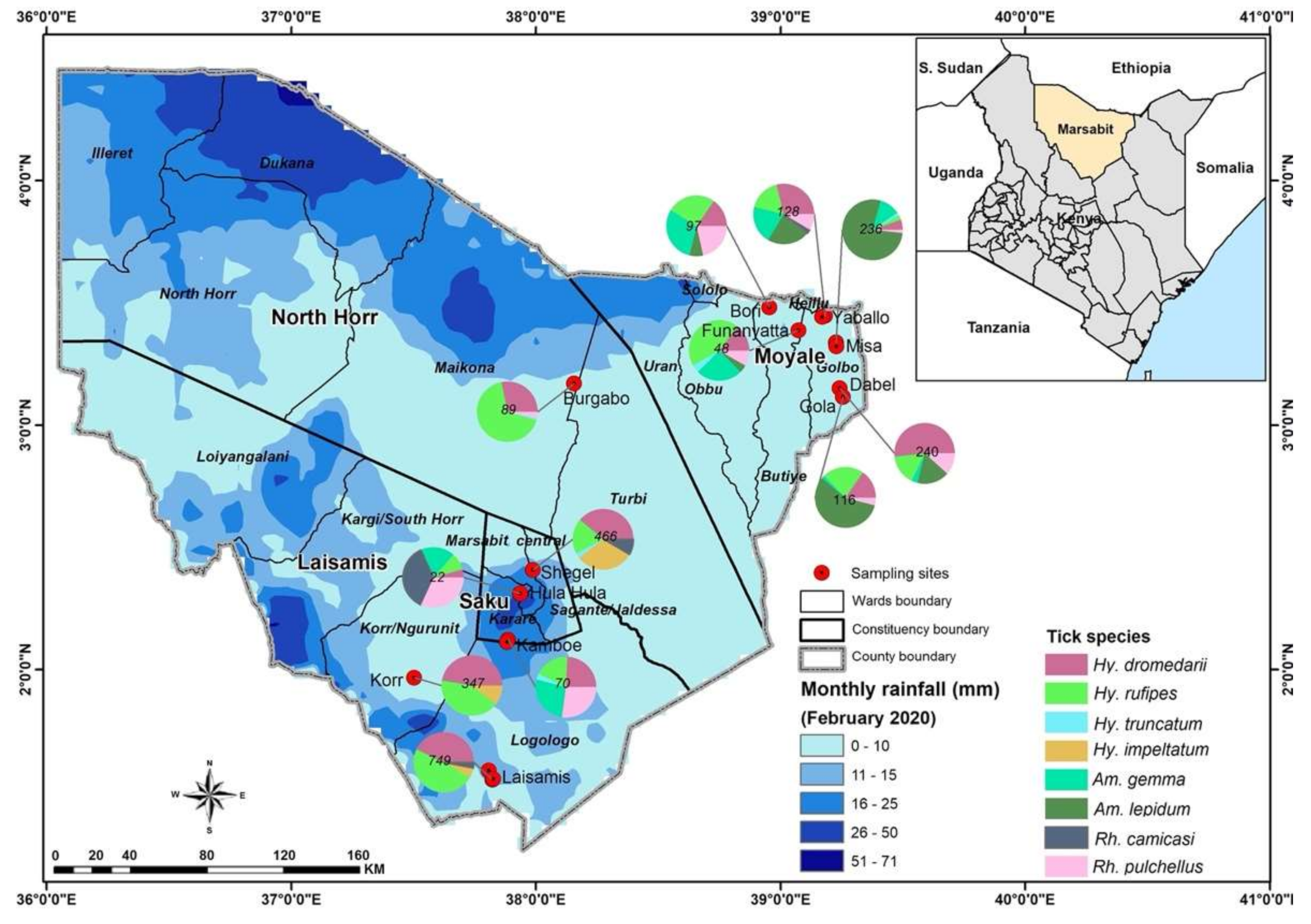
2.1. Study Area
2.2. Ethical Approval
2.3. Collection of Blood Samples and Ticks from Camels and Co-Herded Sheep
2.4. Morphological Identification of Ticks
2.5. Extraction of DNA from Whole Ticks, Tick Leg Tissues and Blood
2.6. Molecular Identification of Ticks
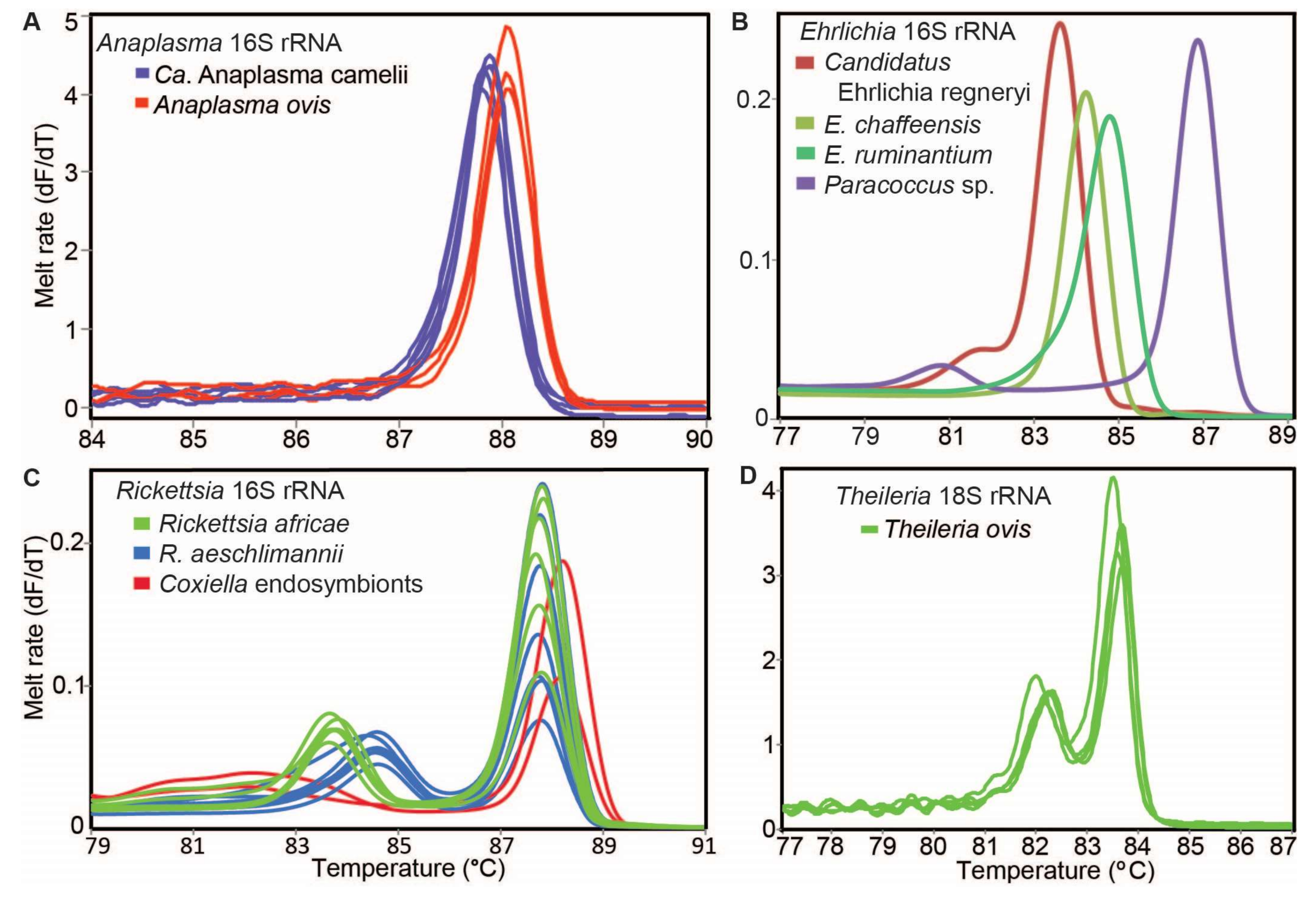
2.7. Molecular Detection of TBPs
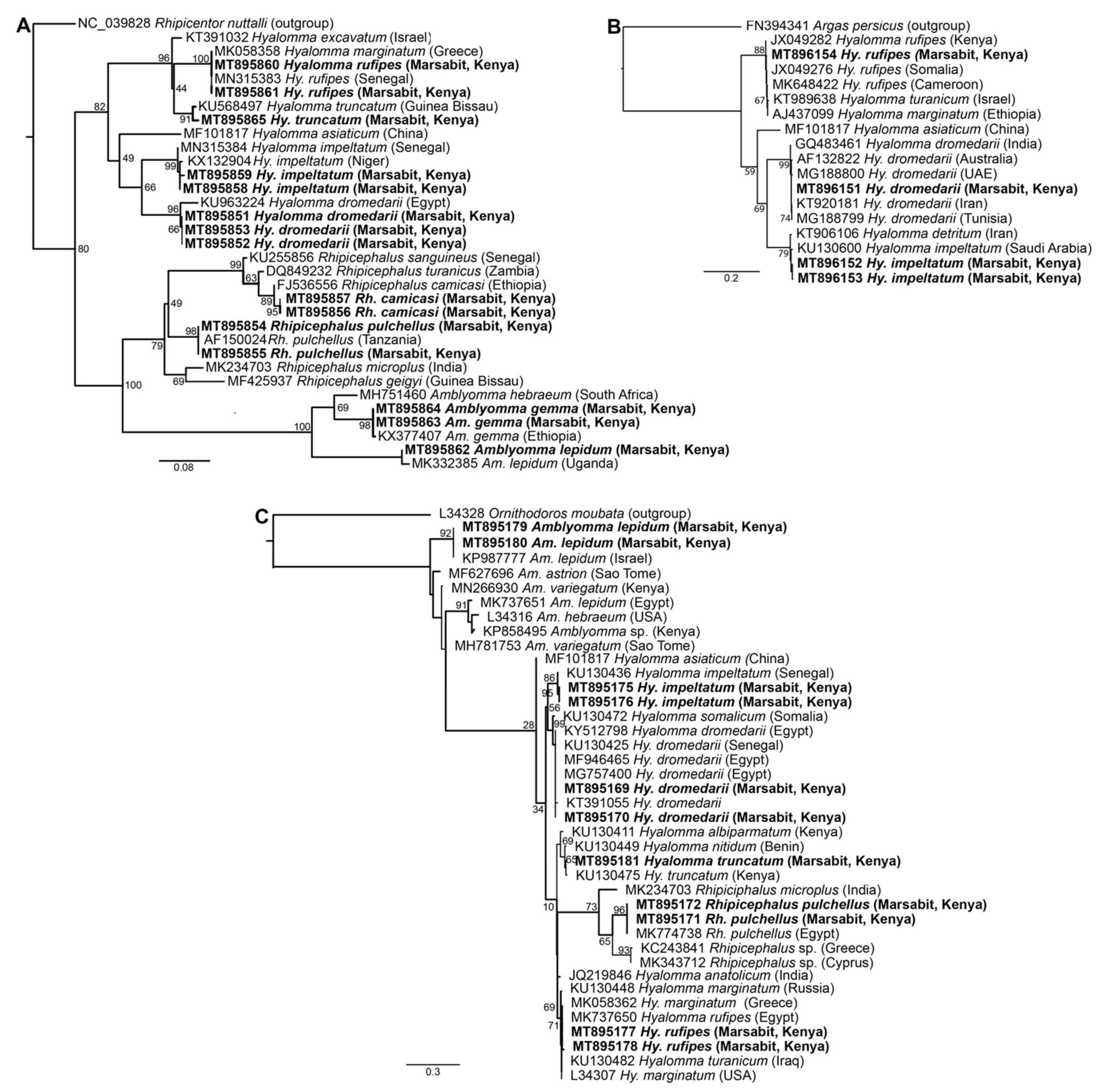
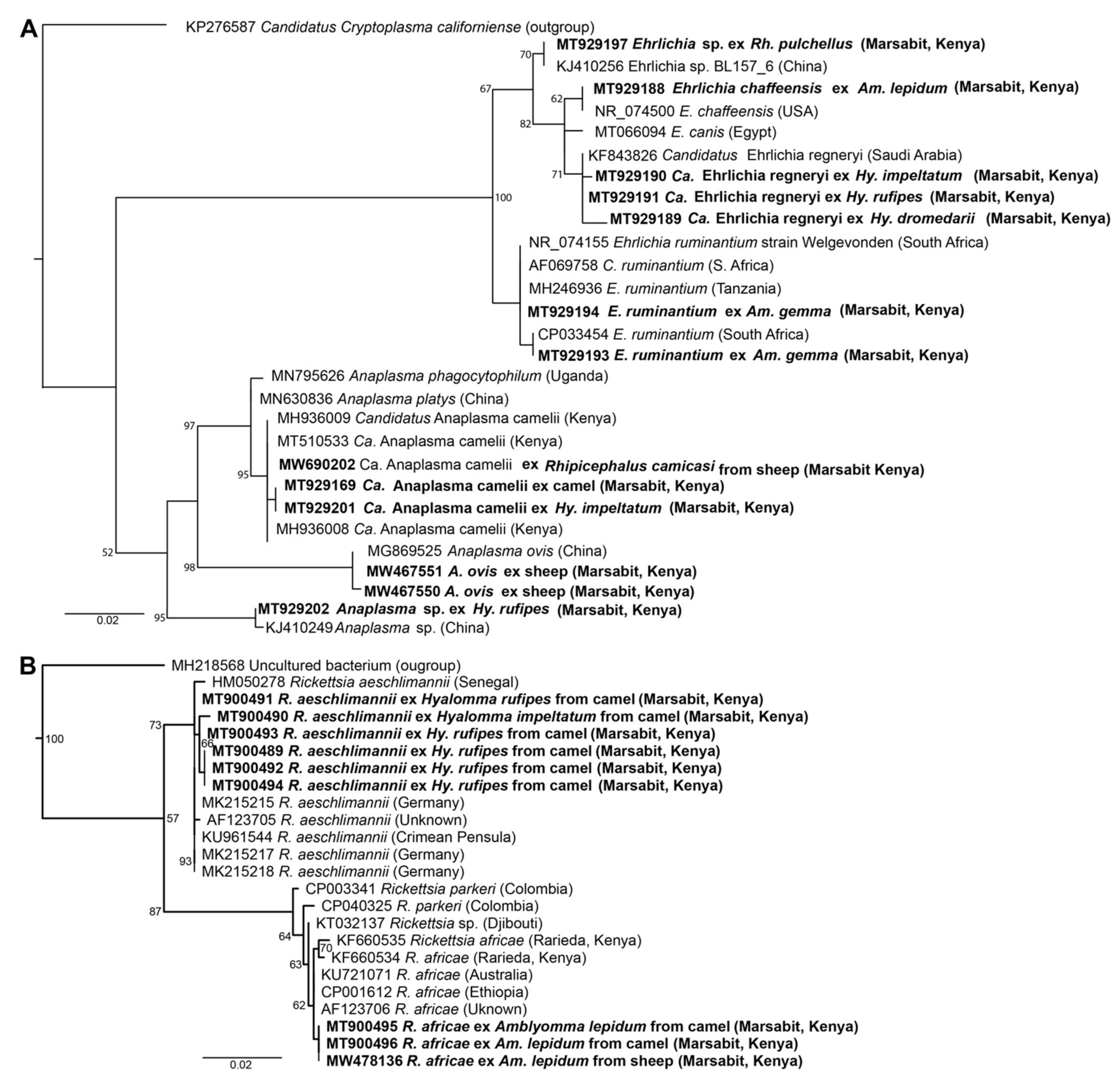
2.8. Phylogenetic Analysis
2.9. Estimation of Tick Infection Rates
3. Results
3.1. Morphological and Molecular Identification of Ticks
3.2. Tick-Borne Pathogens Detected in Camel and Sheep Blood
3.3. Tick-Borne Pathogens and Endosymbionts Detected in Ticks
4. Discussion
4.1. Species Diversity of Ticks Associated with Camels and Co-Herded Sheep in Northern Kenya
4.2. Tick-Borne Bacteria Identified in Ticks, Camels and Co-Herded Sheep in Northern Kenya
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kagunyu, A.W.; Wanjohi, J. Camel rearing replacing cattle production among the Borana community in Isiolo County of northern Kenya, as climate variability bites. Pastoralism 2014, 4, 13. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization Statistical Database. 2016. Available online: http://www.fao.org/faostat/en/#data/QA (accessed on 5 December 2020).
- Bornstein, S.; Younan, M. Significant veterinary research on the dromedary camels of Kenya: Past and present. J. Camelid Sci. 2013, 6, 1–48. [Google Scholar]
- Khaskheli, A.A. A review on several important aspects of the camels. Aceh J. Anim. Sci. 2020, 5, 129–135. [Google Scholar] [CrossRef]
- Abdisa, T.; Wubishet, Z.; Etsay, K. Study on major constraints of camel production, management and their impacts in and around Yabello District, Oromia Regional State, southern Ethiopia. J. Dairy Vet. Sci. 2017, 3, 555–604. [Google Scholar] [CrossRef]
- Kidambasi, K.O.; Masiga, D.K.; Villinger, J.; Carrington, M.; Bargul, J.L. Detection of blood pathogens in camels and their associated ectoparasitic camel biting keds, Hippobosca camelina: The potential application of keds in xenodiagnosis of camel haemopathogens. AAS Open Res. 2020, 2, 164. [Google Scholar] [CrossRef]
- Getahun, M.N.; Villinger, J.; Bargul, J.L.; Orone, A.; Ngiela, J.; Ahuya, P.O.; Muema, J.M.; Saini, R.K.; Torto, B.; Masiga, D.K. Molecular characterization of pathogenic African trypanosomes in biting flies and camels in surra-endemic areas outside the tsetse fly belt in Kenya. bioRxiv 2020, 15, 68–69. [Google Scholar] [CrossRef]
- Oguntomole, O.; Nwaeze, U.; Eremeeva, M. Tick-, flea-, and louse-borne diseases of public health and veterinary significance in Nigeria. Trop. Med. Infect. Dis. 2018, 3, 3. [Google Scholar] [CrossRef]
- Kernif, T.; Leulmi, H.; Raoult, D.; Parola, P. Emerging tick-borne bacterial pathogens. Microbiol. Spectr. 2016, 4, EI10-0012-2016. [Google Scholar] [CrossRef]
- Wesonga, F.D.; Kitala, P.M.; Gathuma, J.M.; Njenga, M.J.; Ngumi, P.N. An assessment of tick-borne diseases constraints to livestock production in a smallholder livestock production system in Machakos District, Kenya. Livest. Res. Rural Dev. 2010, 22, 6. [Google Scholar]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S1–S4. [Google Scholar] [CrossRef]
- Lwande, O.W.; Lutomiah, J.; Obanda, V.; Gakuya, F.; Mutisya, J.; Mulwa, F.; Michuki, G.; Chepkorir, E.; Fischer, A.; Venter, M.; et al. Isolation of tick and mosquito-borne arboviruses from ticks sampled from livestock and wild animal hosts in Ijara District, Kenya. Vector Borne Zoonotic Dis. 2013, 13, 637–642. [Google Scholar] [CrossRef]
- Raboloko, O.O.; Ramabu, S.S.; Guerrini, L.; Jori, F. Seroprevalence of selected tick-borne pathogens and diversity and abundance of Ixodid ticks (Acari: Ixodidae) at the wildlife-livestock interface in northern Botswana. Front. Vet. Sci. 2020, 7, 187. [Google Scholar] [CrossRef]
- Alsarraf, M.; Mierzejewska, E.J.; Mohallal, E.M.E.; Behnke, J.M.; Bajer, A. Genetic and phylogenetic analysis of the ticks from the Sinai Massif, Egypt, and their possible role in the transmission of Babesia behnkei. Exp. Appl. Acarol. 2017, 72, 415–427. [Google Scholar] [CrossRef]
- Alanazi, A.D.; Nguyen, V.L.; Alyousif, M.S.; Manoj, R.R.S.; Alouffi, A.S.; Donato, R.; Sazmand, A.; Mendoza-Roldan, J.A.; Dantas-Torres, F.; Otranto, D. Ticks and associated pathogens in camels (Camelus dromedarius) from Riyadh Province, Saudi Arabia. Parasites Vectors 2020, 13, 110. [Google Scholar] [CrossRef]
- Alanazi, A.D.; Al-Mohammed, H.I.; Alyousif, M.S.; Said, A.E.; Salim, B.; Abdel-Shafy, S.; Shaapan, R.M. Species diversity and seasonal distribution of hard ticks (Acari: Ixodidae) infesting mammalian hosts in various districts of Riyadh Province, Saudi Arabia. J. Med. Entomol. 2019, 56, 1027–1032. [Google Scholar] [CrossRef]
- Dioli, M.; Jean-Baptiste, S.; Fox, M. Ticks (Acari: Ixodidae) of the one-humped camel (Camelus dromedarius) in Kenya and southern Ethiopia: Species composition, attachment sites. Rev. Elev. Med. Vet. Pays. Trop. 2001, 54, 115–122. [Google Scholar] [CrossRef][Green Version]
- Younan, M.; Ouso, D.O.; Bodha, B.; Keitany, E.K.; Wesonga, H.O.; Sitawa, R.; Kimutai, J.; Kuria, W.; Sake, W.S.; Svitek, N.; et al. Ehrlichia spp. close to Ehrlichia ruminantium, Ehrlichia canis, and “Candidatus Ehrlichia regneryi” linked to heartwater-like disease in Kenyan camels (Camelus dromedarius). Trop. Anim. Health Prod. 2021, 53, 147. [Google Scholar] [CrossRef]
- van Vliet, A.H.M.; Zeijst, B.A.M.; Camus, E.; Mahan, S.M.; Martinez, D.; Jongejan, F. Use of a specific immunogenic region on the Cowdria ruminantium MAP1 protein in a serological assay. J. Clin. Microbiol. 1995, 33, 2405–2410. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Koney, E.B.M.; Dogbey, O.; Walker, A.R. Ehrlichia ruminantium seroprevalence in domestic ruminants in Ghana; I. Longitudinal survey in the Greater Accra region. Vet. Microbiol. 2004, 100, 175–188. [Google Scholar] [CrossRef]
- County Government of Marsabit CIDP. County Government of Marsabit: First County Integrated Development Plan 2013–2017. 2013, p. 284. Available online: http://www.kpda.or.ke/documents/CIDP/Marsabit.pdf (accessed on 29 March 2021).
- Siciliano, G.; Bigi, V.; Vigna, I.; Comino, E.; Rosso, M.; Cristofori, E.; Demarchi, A.; Pezzoli, A. Comparison of multiple maximum and minimum temperature datasets at local level: The case study of North Horr sub-County, Kenya. Climate 2021, 9, 62. [Google Scholar] [CrossRef]
- Walker, A.R.; Bouattour, A.; Camicas, J.; Estrada-Peña, A.; Horak, I.G.; Latif, A.A.; Pegram, R.G.; Preston, P.M. Ticks of Domestic Animals in Africa: A Guide to Identification of Tick Species; Bioscience Reports: Edinburgh, UK, 2003; p. 227. [Google Scholar]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Sim on, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Brahma, R.K.; Dixit, V.; Sangwan, A.K.; Doley, R. Identification and characterization of Rhipicephalus (Boophilus) microplus and Haemaphysalis bispinosa ticks (Acari: Ixodidae) of North East India by ITS2 and 16S rDNA sequences and morphological analysis. Exp. Appl. Acarol. 2014, 62, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Nijhof, A.M.; Bodaan, C.; Postigo, M.; Nieuwenhuijs, H.; Opsteegh, M.; Franssen, L.; Jebbink, F.; Jongejan, F. Ticks and associated pathogens collected from domestic animals in the Netherlands. Vector-Borne Zoonotic Dis. 2007, 7, 585–595. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef]
- Hoover, T.A.; Vodkin, M.H.; Williams, J.C. A Coxiella burnetii repeated DNA element resembling a bacterial insertion sequence. J. Bacteriol. 1992, 174, 5540–5548. [Google Scholar] [CrossRef]
- Tokarz, R.; Kapoor, V.; Samuel, J.E.; Bouyer, D.H.; Briese, T.; Lipkin, W.I. Detection of tick-borne pathogens by masstag polymerase chain reaction. Vector-Borne Zoonotic Dis. 2009, 9, 147–151. [Google Scholar] [CrossRef]
- Goodman, J.L.; Nelson, C.; Vitale, B.; Madigan, J.E.; Dumler, J.S.; Kurtti, T.J.; Munderloh, U.G. Direct cultivation of the causative agent of human granulocytic ehrlichiosis. N. Engl. J. Med. 1996, 334, 209–215. [Google Scholar] [CrossRef]
- Parola, P.; Roux, V.; Camicas, J.L.; Baradji, I.; Brouqui, P.; Raoult, D. Detection of ehrlichiae in African ticks by polymerase chain reaction. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 707–708. [Google Scholar] [CrossRef]
- Reysenbach, A.L.; Giver, L.J.; Wickham, G.S.; Pace, N.R. Differential amplification of rRNA genes by polymerase chain reaction. Appl. Environ. Microbiol. 1992, 58, 3417–3418. [Google Scholar] [CrossRef]
- Mwamuye, M.M.; Kariuki, E.; Omondi, D.; Kabii, J.; Odongo, D.; Masiga, D.; Villinger, J. Novel Rickettsia and emergent tick-borne pathogens: A molecular survey of ticks and tick-borne pathogens in Shimba Hills National Reserve, Kenya. Ticks Tick Borne Dis. 2017, 8, 208–218. [Google Scholar] [CrossRef]
- Gubbels, J.M.; De Vos, A.P.; Van Der Weide, M.; Viseras, J.; Schouls, L.M.; De Vries, E.; Jongejan, F. Simultaneous detection of bovine Theileria and Babesia species by reverse line blot hybridization. J. Clin. Microbiol. 1999, 37, 1782–1789. [Google Scholar] [CrossRef]
- Oundo, J.W.; Villinger, J.; Jeneby, M.; Ong’amo, G.; Otiende, M.Y.; Makhulu, E.E.; Musa, A.A.; Ouso, D.O.; Wambua, L. Pathogens, endosymbionts, and blood-meal sources of host-seeking ticks in the fast-changing Maasai Mara wildlife eco-system. PLoS ONE 2020, 15, e0228366. [Google Scholar] [CrossRef]
- Chiuya, T.; Masiga, D.K.; Falzon, L.C.; Bastos, A.D.S.; Fèvre, E.M.; Villinger, J. Tick-borne pathogens, including Crimean-Congo haemorrhagic fever virus, at livestock markets and slaughterhouses in western Kenya. Transbound. Emerg. Dis. 2020. Online ahead of print. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree; Version 1.4.4; University of Edinburgh: Edinburgh, UK, 2020. [Google Scholar]
- Okal, M.N.; Odhiambo, B.K.; Otieno, P.; Bargul, J.L.; Masiga, D.; Villinger, J.; Kalayou, S. Anaplasma and Theileria pathogens in cattle of Lambwe Valley, Kenya: A case for pro-active surveillance in the wildlife—Livestock interface. Microorganisms 2020, 8, 1830. [Google Scholar] [CrossRef]
- Wikel, S.K. Ticks and tick-borne infections: Complex ecology, agents, and host interactions. Vet. Sci. 2018, 5, 60. [Google Scholar] [CrossRef]
- Kanduma, E.G.; Mwacharo, J.M.; Mwaura, S.; Njuguna, J.N.; Nzuki, I.; Kinyanjui, P.W.; Githaka, N.; Heyne, H.; Hanotte, O.; Skilton, R.A.; et al. Multi-locus genotyping reveals absence of genetic structure in field populations of the brown ear tick (Rhipicephalus appendiculatus) in Kenya. Ticks Tick Borne Dis. 2016, 7, 26–35. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef]
- Elghali, A.; Hassan, S.M. Ticks (Acari: Ixodidae) infesting camels (Camelus dromedarius) in northern Sudan. Onderstepoort J. Vet. Res. 2009, 76, 177–185. [Google Scholar] [CrossRef]
- Youssef, S.Y.; Yasien, S.; Mousa, W.M.A.; Nasr, S.M.; El-Kelesh, E.A.M.; Mahran, K.M.; Abd-El-Rahman, A.H. Vector iden-tification and clinical, hematological, biochemical, and parasitological characteristics of camel (Camelus dromedarius) theileriosis in Egypt. Trop. Anim. Health Prod. 2015, 47, 649–656. [Google Scholar] [CrossRef]
- Wallménius, K.; Barboutis, C.; Fransson, T.; Jaenson, T.G.; Lindgren, P.E.; Nyström, F.; Olsen, B.; Salaneck, E.; Nilsson, K. Spotted fever Rickettsia species in Hyalomma and Ixodes ticks infesting migratory birds in the European Mediterranean area. Parasites Vectors 2014, 7, 318. [Google Scholar] [CrossRef]
- Kleinerman, G.; Baneth, G.; Mumcuoglu, K.Y.; Van Straten, M.; Berlin, D.; Apanaskevich, D.A.; Abdeen, Z.; Nasereddin, A.; Harrus, S. Molecular detection of Rickettsia africae, Rickettsia aeschlimannii, and Rickettsia sibirica mongolitimonae in camels and Hyalomma spp. ticks from Israel. Vector Borne Zoonotic Dis. 2013, 13, 851–856. [Google Scholar] [CrossRef]
- Kernif, T.; Djerbouh, A.; Mediannikov, O.; Ayach, B.; Rolain, J.M.; Raoult, D.; Parola, P.; Bitam, I. Rickettsia africae in Hyalomma dromedarii ticks from sub-Saharan Algeria. Ticks Tick Borne Dis. 2012, 3, 377–379. [Google Scholar] [CrossRef]
- Bellabidi, M.; Benaissa, M.H.; Bissati-Bouafia, S.; Harrat, Z.; Brahmi, K.; Kernif, T. Coxiella burnetii in camels (Camelus dromedarius) from Algeria: Seroprevalence, molecular characterization, and ticks (Acari: Ixodidae) vectors. Acta Trop. 2020, 206, 105443. [Google Scholar] [CrossRef] [PubMed]
- Schelling, E.; Diguimbaye, C.; Daoud, S.; Nicolet, J.; Boerlin, P.; Tanner, M.; Zinsstag, J. Brucellosis and Q-fever seroprev-alences of nomadic pastoralists and their livestock in Chad. Prev. Vet. Med. 2003, 61, 279–293. [Google Scholar] [CrossRef]
- Selmi, R.; Ben Said, M.; Dhibi, M.; Ben Yahia, H.; Messadi, L. Improving specific detection and updating phylogenetic data related to Anaplasma platys-like strains infecting camels (Camelus dromedarius) and their ticks. Ticks Tick Borne Dis. 2019, 10, 101260. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, N.H.; Abdel-Moein, K.A.; Zaher, H.M. Molecular detection of Francisella spp. among ticks attached to camels in Egypt. Vector Borne Zoonotic Dis. 2017, 17, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Moshaverinia, A.; Moghaddas, E. Prevalence of tick infestation in dromedary camels (Camelus dromedarius) brought for slaughter in Mashhad abattoir, Iran. J. Parasit. Dis. 2015, 39, 452–455. [Google Scholar] [CrossRef]
- Rehman, A.; Nijhof, A.M.; Sauter-Louis, C.; Schauer, B.; Staubach, C.; Conraths, F.J. Distribution of ticks infesting ruminants and risk factors associated with high tick prevalence in livestock farms in the semi-arid and arid agro-ecological zones of Pakistan. Parasites Vectors 2017, 10, 190. [Google Scholar] [CrossRef]
- Shemshad, M.; Shemshad, K.; Sedaghat, M.M.; Shokri, M.; Barmaki, A.; Baniardalani, M.; Rafinejad, J. First survey of hard ticks (Acari: Ixodidae) on cattle, sheep and goats in Boeen Zahra and Takistan counties, Iran. Asian Pac. J. Trop. Biomed. 2012, 2, 489–492. [Google Scholar] [CrossRef]
- Chitimia-Dobler, L.; Schaper, S.; Rieß, R.; Bitterwolf, K.; Frangoulidis, D.; Bestehorn, M.; Springer, A.; Oehme, R.; Drehmann, M.; Lindau, A.; et al. Imported Hyalomma ticks in Germany in 2018. Parasites Vectors 2019, 12, 134. [Google Scholar] [CrossRef]
- Omondi, D.; Masiga, D.K.; Fielding, B.C.; Kariuki, E.; Ajamma, Y.U.; Mwamuye, M.M.; Ouso, D.O.; Villinger, J. Molecular detection of tick-borne pathogen diversities in ticks from livestock and reptiles along the shores and adjacent islands of Lake Victoria and Lake Baringo, Kenya. Front. Vet. Sci. 2017, 4, 73. [Google Scholar] [CrossRef]
- Kamani, J.; Baneth, G.; Mumcuoglu, K.Y.; Waziri, N.E.; Eyal, O.; Guthmann, Y.; Harrus, S. Molecular detection and characterization of tick-borne pathogens in dogs and ticks from Nigeria. PLoS Negl. Trop. Dis. 2013, 7, e2108. [Google Scholar] [CrossRef]
- Kariuki, E.K.; Penzhorn, B.L.; Horak, I.G. Ticks (Acari: Ixodidae) infesting cattle and African buffaloes in the Tsavo conservation area, Kenya. Onderstepoort J. Vet. Res. 2012, 79, 437–441. [Google Scholar] [CrossRef]
- Dohm, D.J.; Logan, T.M.; Linthicum, K.J.; Rossi, C.A.; Turell, M.J. Transmission of Crimean-Congo hemorrhagic fever virus by Hyalomma impeltatum (Acari: Ixodidae) after experimental infection. J. Med. Entomol. 1996, 33, 848–851. [Google Scholar] [CrossRef]
- Allsopp, B.A. Heartwater—Ehrlichia ruminantium infection. Rev. Sci. Tech. 2015, 34, 557–568. [Google Scholar] [CrossRef]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.J.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combi. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef]
- Walker, J.B.; Olwage, A. The tick vectors of Cowdria ruminantium (Ixodoidea, Ixodidae, genus Amblyomma) and their distribution. Onderstepoort J. Vet. Res. 1987, 54, 353–379. [Google Scholar] [PubMed]
- Postigo, M.; Bell-Sakyi, L.; Paxton, E.; Sumption, K. Kinetics of experimental infection of sheep with Ehrlichia ruminantium cultivated in tick and mammalian cell lines. Exp. Appl. Acarol. 2002, 28, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Andrew, H.R.; Norval, R.A.I. The carrier status of sheep, cattle and African buffalo recovered from heartwater. Vet. Parasitol. 1989, 34, 261–266. [Google Scholar] [CrossRef]
- Bechir, A. OIE Immediate Notification Report (31/12/2013). 2013. Available online: http://www.oie.int/wahis_2/public/wahid.php/Reviewreport/Review?page_refer=MapFullEventReport&reportid=14588 (accessed on 15 November 2020).
- Ngumi, P.N.; Rumberia, R.M.; Williamson, S.M.; Sumption, K.J.; Lesan, A.C.; Kariuki, D.P. Isolation of the causative agent of heartwater (Cowdria ruminantium) from three Amblyomma species in eight districts of Kenya. Vet. Rec. 1997, 140, 13–16. [Google Scholar] [CrossRef]
- Paddock, C.D.; Childs, J.E. Ehrlichia chaffeensis: A prototypical emerging pathogen. Clin. Microbiol. Rev. 2003, 16, 37–64. [Google Scholar] [CrossRef]
- Proboste, T.; Kalema-Zikusoka, G.; Altet, L.; Solano-Gallego, L.; Fernández De Mera, I.G.; Chirife, A.D.; Muro, J.; Bach, E.; Piazza, A.; Cevidanes, A.; et al. Infection and exposure to vector-borne pathogens in rural dogs and their ticks, Uganda. Parasites Vectors 2015, 8, 306. [Google Scholar] [CrossRef]
- Ndip, L.M.; Ndip, R.N.; Esemu, S.N.; Walker, D.H.; McBride, J.W. Predominance of Ehrlichia chaffeensis in Rhipicephalus sanguineus ticks from kennel-confined dogs in Limbe, Cameroon. Exp. Appl. Acarol. 2010, 50, 163–168. [Google Scholar] [CrossRef]
- Iweriebor, B.C.; Mmbaga, E.J.; Adegborioye, A.; Igwaran, A.; Obi, L.C.; Okoh, A.I. Genetic profiling for Anaplasma and Ehrlichia species in ticks collected in the Eastern Cape Province of South Africa. BMC Microbiol. 2017, 17, 45. [Google Scholar] [CrossRef]
- Bastos, A.D.S.; Mohammed, O.B.; Bennett, N.C.; Petevinos, C.; Alagaili, A.N. Molecular detection of novel Anaplasmataceae closely related to Anaplasma platys and Ehrlichia canis in the dromedary camel (Camelus dromedarius). Vet. Microbiol. 2015, 179, 310–314. [Google Scholar] [CrossRef]
- Frangoulidis, D.; Kahlhofer, C.; Said, A.S.; Osman, A.Y.; Chitimia-Dobler, L.; Shuaib, Y.A. High prevalence and new genotype of Coxiella burnetii in ticks infesting camels in Somalia. Pathogens 2021, 10, 741. [Google Scholar] [CrossRef]
- Devaux, C.A.; Osman, I.O.; Million, M.; Raoult, D. Coxiella burnetii in dromedary camels (Camelus dromedarius): A possible threat for humans and livestock in North Africa and the Near and Middle East? Front Vet Sci. 2020, 7, 558481. [Google Scholar] [CrossRef]
- Selmi, R.; Ben Said, M.; Mamlouk, A.; Ben Yahia, H.; Messadi, L. Molecular detection and genetic characterization of the potentially pathogenic Coxiella burnetii and the endosymbiotic Candidatus Midichloria mitochondrii in ticks infesting camels (Camelus dromedarius) from Tunisia. Microb. Pathog. 2019, 136, 103655. [Google Scholar] [CrossRef]
- Songer, J.G.; Post, K.W. Coxiella and Cowdria. In Veterinary Microbiology: Bacterial and Fungal Agents of Animal Disease, 1st ed.; Songer, J.G., Post, K.W., Eds.; Elsevier/Saunders: MO, USA, 2005; pp. 339–342. [Google Scholar]
- Browne, A.S.; Fèvre, E.M.; Kinnaird, M.; Muloi, D.M.; Wang, C.A.; Larsen, P.S.; O’Brien, T.; Deem, S.L. Serosurvey of Coxiella burnetii (Q fever) in dromedary camels (Camelus dromedarius) in Laikipia County, Kenya. Zoonoses Public Health 2017, 64, 543–549. [Google Scholar] [CrossRef]
- Kirkan, S.; Kaya, O.; Tekbiyik, S.; Parin, U. Detection of Coxiella burnetii in cattle by PCR. Turkish. J. Vet. Anim. Sci. 2008, 32, 215–220. [Google Scholar]
- Duron, O.; Sidi-Boumedine, K.; Rousset, E.; Moutailler, S.; Jourdain, E. The importance of ticks in Q Fever transmission: What has (and has not) been demonstrated? Trends Parasitol. 2015, 31, 536–552. [Google Scholar] [CrossRef]
- Knobel, D.L.; Maina, A.N.; Cutler, S.J.; Ogola, E.; Feikin, D.R.; Junghae, M.; Halliday, J.E.B.; Richards, J.L.; Breiman, R.S.; Cleaveland, S.; et al. Coxiella burnetii in humans, domestic ruminants, and ticks in rural Western Kenya. Am. J. Trop. Med. Hyg. 2013, 88, 513–518. [Google Scholar] [CrossRef]
- Koka, H.; Sang, R.; Kutima, H.L.; Musila, L. Coxiella burnetii detected in tick samples from pastoral communities in Kenya. Biomed. Res. Int. 2018, 54, 774–780. [Google Scholar] [CrossRef]
- Ndeereh, D.; Muchemi, G.; Thaiyah, A.; Otiende, M.; Angelone-Alasaad, S.; Jowers, M.J. Molecular survey of Coxiella burnetii in wildlife and ticks at wildlife–livestock interfaces in Kenya. Exp. Appl. Acarol. 2017, 72, 277–289. [Google Scholar] [CrossRef]
- Mediannikov, O.; Fenollar, F.; Socolovschi, C.; Diatta, G.; Bassene, H.; Molez, J.F.; Sokhna, C.; Trape, J.-S.; Raoult, D. Coxiella burnetii in humans and ticks in rural Senegal. PLoS Negl. Trop. Dis. 2010, 4, e654. [Google Scholar] [CrossRef]
- Neare, K.; Janson, M.; Hütt, P.; Lassen, B.; Viltrop, A. Coxiella burnetii antibody prevalence and risk factors of infection in the human population of Estonia. Microorganisms 2019, 7, 629. [Google Scholar] [CrossRef]
- Szymańska-Czerwińska, M.; Galińska, E.M.; Niemczuk, K.; Knap, J.P. Prevalence of Coxiella burnetii infection in humans occupationally exposed to animals in Poland. Vector Borne Zoonotic Dis. 2015, 15, 261–267. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Rot, A.; Mahagna, M.; Kapri, E.; Behar, A.; Gottlieb, Y. Coxiella-like endosymbiont of Rhipicephalus sanguineus is required for physiological processes during ontogeny. Front. Microbiol. 2020, 11, 493. [Google Scholar] [CrossRef]
- Zhong, J.; Jasinskas, A.; Barbour, A.G. Antibiotic treatment of the tick vector Amblyomma americanum reduced reproductive fitness. PLoS ONE 2007, 2, e405. [Google Scholar] [CrossRef]
- Belkahia, H.; Said, M.B.; Sayahi, L.; Alberti, A.; Messadi, L. Detection of novel strains genetically related to Anaplasma platys in Tunisian one-humped camels (Camelus dromedarius). J. Infect. Dev. Ctries. 2015, 9, 1117–1125. [Google Scholar] [CrossRef]
- Azmat, M.; Ijaz, M.; Farooqi, S.H.; Ghaffar, A.; Ali, A.; Masud, A.; Saleem, S.; Rehman, A.; Ali, M.M.; Mehmood, K.; et al. Molecular epidemiology, associated risk factors, and phylogenetic analysis of anaplasmosis in camel. Microb. Pathog. 2018, 123, 377–384. [Google Scholar] [CrossRef]
- Bahrami, S.; Hamidinejat, H.; Ganjali, A.R. First molecular detection of Anaplasma phagocytophilum in dromedaries (Camelus dromedarius). J. Zoo Wildl. Med. 2018, 49, 844–848. [Google Scholar] [CrossRef]
- Brown, W.C. Adaptive immunity to Anaplasma pathogens and immune dysregulation: Implications for bacterial persistence. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 241–252. [Google Scholar] [CrossRef][Green Version]
- Bargul, J.; Kidambasi, K.; Getahun, M.; Villinger, J.; Copeland, R.; Muema, J.; Carrington, M.; Masiga, D.K. Transmission of ‘Candidatus Anaplasma camelii’ to laboratory animals by camel-specific keds, Hippobosca camelina. PLOS Negl. Trop. Dis. 2021, in press. [Google Scholar]
- Lacasta, D.; Ferrer, L.M.; Sanz, S.; Labanda, R.; González, J.M.; Benito, A.Á.; Ruiz, H.; Rodríguez-Largo, A.; Ramos, J.J. Anaplasmosis outbreak in lambs: First report causing carcass condemnation. Animals 2020, 10, 1851. [Google Scholar] [CrossRef] [PubMed]
- Koka, H.; Sang, R.; Kutima, H.L.; Musila, L.; Macaluso, K. The detection of spotted fever group Rickettsia DNA in tick samples from pastoral communities in Kenya. J. Med. Entomol. 2017, 54, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Selmi, R.; Ben Said, M.; Ben Yahia, H.; Abdelaali, H.; Messadi, L. Molecular epidemiology and phylogeny of spotted fever group Rickettsia in camels (Camelus dromedarius) and their infesting ticks from Tunisia. Transbound. Emerg. Dis. 2019, 67, 733–744. [Google Scholar] [CrossRef]
- Gebrekidan, H.; Hailu, A.; Kassahun, A.; Rohoušová, I.; Maia, C.; Talmi-Frank, D.; Warburg, A.; Baneth, G. Theileria infection in domestic ruminants in northern Ethiopia. Vet. Parasitol. 2014, 200, 31–38. [Google Scholar] [CrossRef]
- El Imam, A.H.; Hassan, S.M.; Gameel, A.A.; Abdelrahim, M.; Hussein, E.; Taha, K.M.; Oosthuizen, M.C. Molecular identi-fication of different Theileria and Babesia species infecting sheep in Sudan. Polish Parasitol. Soc. 2016, 62, 47–54. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Koney, E.B.M.; Dogbey, O.; Walker, A.R. Incidence and prevalence of tick-borne haemoparasites in domestic ruminants in Ghana. Vet. Parasitol. 2004, 124, 25–42. [Google Scholar] [CrossRef]
- Machado-Ferreira, E.; Piesman, J.; Zeidner, N.S.; Soares, C.A.G. A prevalent alpha-proteobacterium Paracoccus sp. in a population of the Cayenne ticks (Amblyomma cajennense) from Rio de Janeiro, Brazil. Genet. Mol. Biol. 2012, 35, 862–867. [Google Scholar] [CrossRef]
- Egyed, L.; Makrai, L. Cultivable internal bacterial flora of ticks isolated in Hungary. Exp. Appl. Acarol. 2014, 63, 107–122. [Google Scholar] [CrossRef]
- Rojas-Jaimes, J.; Lindo-Seminario, D.; Correa-Nunez, G.; Diringer, B. Characterization of the bacterial microbiome of Rhipicephalus (Boophilus) microplus collected from Pecari tajacu “Sajino” Madre de Dios, Peru. Sci. Rep. 2021, 11, 6661. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Target Gene | Sequence (5’–3’) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| Tick COI F Tick COI R | Tick COI | ATTCAACCAATCATAAAGATATTGG TAAACTTCTGGATGTCCAAAAAATCA | 658 | [24] |
| SR-J-14199F SR-N-14594R | Tick 12S rRNA | TACTATGTTACGACTTAT AAACTAGGATTAGATACCC | 430 | [25] |
| Tick 16S Tick 16S | Tick 16S rRNA | AATTGCTGTAGTATTTTGAC TCTGAACTCAGATCAAGTAG | 450 | [26] |
| Rick-F Rick-R | Rickettsia 16S rRNA | GAACGCTATCGGTATGCTTAACACA CATCACTCACTCGGTATTGCTGGA | 364 | [27] |
| 120–2788 120–3599 | Rickettsia ompB | AAACAATAATCAAGGTACTGT TACTTCCGGTTACAGCAAAGT | 856 | [28] |
| Trans1 Trans2 | Coxiella IS1111 | TGGTATTCTTGCCGATGAC GATCGTAACTGCTTAATAAACCG | 687 | [29] |
| Ehrlichia16S F Ehrlichia16S R | Ehrlichia 16S rRNA | CGTAAAGGGCACGTAGGTGGACTA CACCTCAGTGTCAGTATCGAACCA | 200 | [30] |
| PER1 PER2 | Ehrlichia 16S rRNA | TTTATCGCTATTAGATGAGCCTATG CTCTACACTAGGAATTCCGCTAT | 451 | [31] |
| EHR16SD 1492R | Anaplasma/Ehrlichia 16S rRNA | GGTACCYACAGAAGAAGTCC GGTTACCTTGTTACGACTT | 1030 | [32,33] |
| AnaplasmaJV F AnaplasmaJV R | Anaplasma 16S rRNA | CGGTGGAGCATGTGGTTTAATTC CGRCGTTGCAACCTATTGTAGTC | 300 | [34] |
| RLB F RLB R | Theileria/Babesia 18S rRNA | GAGGTAGTGACAAGAAATAACAATA TCTTCGATCCCCTAACTTTC | 460–520 bp | [35] |
| From 296 Camels | From 77 Co-Herded Sheep | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Male | Female | No. of Pools | No. of Ticks | Percent (%) | Male | Female | No. of Pools | No. of Ticks | Percent (%) |
| Amblyomma gemma | 80 | 49 | 87 | 129 | 4.95 | 11 | 4 | 12 | 15 | 17.44 |
| Amblyomma lepidum | 186 | 144 | 120 | 330 | 12.64 | 20 | 4 | 12 | 24 | 27.91 |
| Hyalomma dromedarii | 624 | 295 | 233 | 919 | 35.21 | |||||
| Hyalomma rufipes | 557 | 253 | 251 | 810 | 31.03 | 1 | 1 | 1 | 1.16 | |
| Hyalomma truncatum | 19 | 6 | 12 | 25 | 0.96 | |||||
| Hyalomma impeltatum | 153 | 68 | 44 | 221 | 8.47 | |||||
| Rhipicephalus pulchellus | 73 | 31 | 66 | 104 | 3.98 | 1 | 1 | 1 | 1.16 | |
| Rhipicephalus camicasi | 30 | 42 | 24 | 72 | 2.76 | 22 | 23 | 22 | 45 | 52.33 |
| Total | 1734 | 876 | 858 | 2610 | 55 | 31 | 48 | 86 | ||
| Bacterial Species (Target Gene) | TBP Detection in Ticks—Number of Positive Pools (Minimum Infection Rate) | Camels with TBPs (Infection Rate) | GenBank Accessions | Nucleotide Sequence Identity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hy. dromedarii | Hy. rufipes | Hy. impeltatum | Hy. truncatum | Am. gemma | Am. lepidum | Rh. camicasi | Rh. pulchellus | Study Sequences | Reference GenBank Accessions | |||
| No. of individuals | 919 ticks | 810 ticks | 221 ticks | 25 ticks | 129 ticks | 330 ticks | 72 ticks | 104 ticks | 296 camels | |||
| Number of tick pools | 254 | 251 | 44 | 12 | 87 | 120 | 24 | 66 | ||||
| Ehrlichia ruminantium(16S rRNA) | 16 (12.40%) | 17 (5.15%) | MT929193-MT929195 | NR_074155, KU721071, CP001612 | 100% | |||||||
| Ca. Ehrlichia regneryi (16S rRNA) | 22 (2.39%) | 46 (5.68%) | 6 (2.72%) | 43 (14.53%) | MT929189-MT929192 | KF843826 | 100% | |||||
| Ehrlichia chaffeensis (16S rRNA) | 2 (0.61%) | MT929188 | NR_074500, NR_074501, CP007473-CP007480 | 100% | ||||||||
| Ehrlichia sp. (16S rRNA) | 1 (0.12%) | 1 (0.78%) | 3 (4.17%) | 18 (17.31%) | MT929196-MT929197 | MN726921, KJ410256 | 100% | |||||
| Candidatus Anaplasma camelii (16S rRNA) | 25 (2.72%) | 27 (3.33%) | 6 (2.72%) | 1 (4%) | 11 (8.53%) | 20 (6.06%) | 6 (8.33%) | 7 (6.73%) | 233 (78.72%) | MT929199-MT929201, MT929169-MT929177 | MT510533, MK388297 | 100% |
| Anaplasma sp. (16S rRNA) | 1 (0.12%) | MT929202 | KJ410248, KJ410249 | 100% | ||||||||
| Rickettsia africae (ompB) | 14 (10.85%) | 31 (9.39%) | MT900495-MT900496 | KU721071, KT032136, CP0011612 | 100% | |||||||
| Rickettsia aeschlimannii(ompB) | 3 (0.33%) | 87 (1.07%) | 6 (2.72%) | 1 (4.00%) | 5 (4.81%) | MT900489-MT900494 | MK215215-MK215218 | 100% | ||||
| Coxiella burnetii (IS1111) | 11 (1.20%) | 12 (1.50%) | 5 (4.81%) | 10 (3.38%) | MT900497-MT900501 | DQ379976, KT954146, MT268529 | 100% | |||||
| Coxiella endo- symbiont (16S rRNA) | 12 (9.30%) | 16 (22.22%) | 6 (5.77%) | MW541904-MW541911 | EU143670, JX846589, MK026405 | 98–100% | ||||||
| Paracoccus sp. (16S rRNA) 1 | 2 (0.22%) | 8 (1.00%) | 2 (0.90%) | 1 (4.00%) | 1 (0.78%) | 3 (2.88%) | 2 | KP003988 | 99% | |||
| Bacterial Species (Target Gene) | TBP Detection in Ticks—Number of Positive Pools (Minimum Infection Rate) | Sheep with TBPs (Infection Rate) | GenBank Accessions | Nucleotide Sequence Identity | |||||
|---|---|---|---|---|---|---|---|---|---|
| Hy. rufipes | Am. gemma | Am. lepidum | Rh. camicasi | Rh. pulchellus | Study Sequences | Reference GenBank Accessions | |||
| No. of individuals | 1 tick | 14 ticks | 24 ticks | 45 ticks | 1 tick | 77 sheep | |||
| No. of tick pools | 1 | 12 | 12 | 22 | 1 | ||||
| Ehrlichia ruminantium (16S rRNA) | 2 (14.29%) | 1 (4.17%) | 1 (1.30%) | MW467546 | NR_074155, MH246936, U03776 | 100% | |||
| Ehrlichia chaffeensis (16S rRNA) | 2 (2.60%) | NR_074501 | 100% | ||||||
| Anaplasma ovis (16S rRNA) | 2 (14.29%) | 2 (8.33%) | 7 (15.56%) | 1 (100%) | 68 (88.31%) | MW467547-MW467552 | MG869525 | 100% | |
| Candidatus Anaplasma camelii (16S rRNA) | 1 (2.22%) | MW690202 | MN630836 | 100% | |||||
| Rickettsia africae (ompB) | 2 (14.29%) | 4 (16.67%) | MW478135-MW478138 | KU721071 | 100% | ||||
| Theileria ovis (18S rRNA) | 1 (2.22%) | 62 (80.52%) | MW467555-MW467561 | MN712508, KX273858, MG738321 | 100% | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Getange, D.; Bargul, J.L.; Kanduma, E.; Collins, M.; Bodha, B.; Denge, D.; Chiuya, T.; Githaka, N.; Younan, M.; Fèvre, E.M.; et al. Ticks and Tick-Borne Pathogens Associated with Dromedary Camels (Camelus dromedarius) in Northern Kenya. Microorganisms 2021, 9, 1414. https://doi.org/10.3390/microorganisms9071414
Getange D, Bargul JL, Kanduma E, Collins M, Bodha B, Denge D, Chiuya T, Githaka N, Younan M, Fèvre EM, et al. Ticks and Tick-Borne Pathogens Associated with Dromedary Camels (Camelus dromedarius) in Northern Kenya. Microorganisms. 2021; 9(7):1414. https://doi.org/10.3390/microorganisms9071414
Chicago/Turabian StyleGetange, Dennis, Joel L. Bargul, Esther Kanduma, Marisol Collins, Boku Bodha, Diba Denge, Tatenda Chiuya, Naftaly Githaka, Mario Younan, Eric M. Fèvre, and et al. 2021. "Ticks and Tick-Borne Pathogens Associated with Dromedary Camels (Camelus dromedarius) in Northern Kenya" Microorganisms 9, no. 7: 1414. https://doi.org/10.3390/microorganisms9071414
APA StyleGetange, D., Bargul, J. L., Kanduma, E., Collins, M., Bodha, B., Denge, D., Chiuya, T., Githaka, N., Younan, M., Fèvre, E. M., Bell-Sakyi, L., & Villinger, J. (2021). Ticks and Tick-Borne Pathogens Associated with Dromedary Camels (Camelus dromedarius) in Northern Kenya. Microorganisms, 9(7), 1414. https://doi.org/10.3390/microorganisms9071414