
A Metzincin and TIMP-Like Protein Pair of a Phage Origin Sensitize Listeria monocytogenes to Phage Lysins and Other Cell Wall Targeting Agents


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Importance
2. Introduction
3. Materials and Methods
3.1. Bacterial Strains and Media
3.2. Generation of Gene Deletion Mutants and Ectopic Expression Strains
3.3. Lm Growth Assays
3.4. Mutanolysin and Triton X-100 Lysis Assays
3.5. Plaque Forming Assay
3.6. Quantitative Real-Time PCR Analysis
3.7. Lm Intracellular Growth
3.8. Transcription Analysis of Intracellular Bacteria
3.9. Biofilm Formation Analysis
3.10. Western Blot Analysis
3.11. Protein Sequencing of Tim
3.12. Transmission Electron Microscopy
3.13. Scanning Electron Microscopy
4. Results
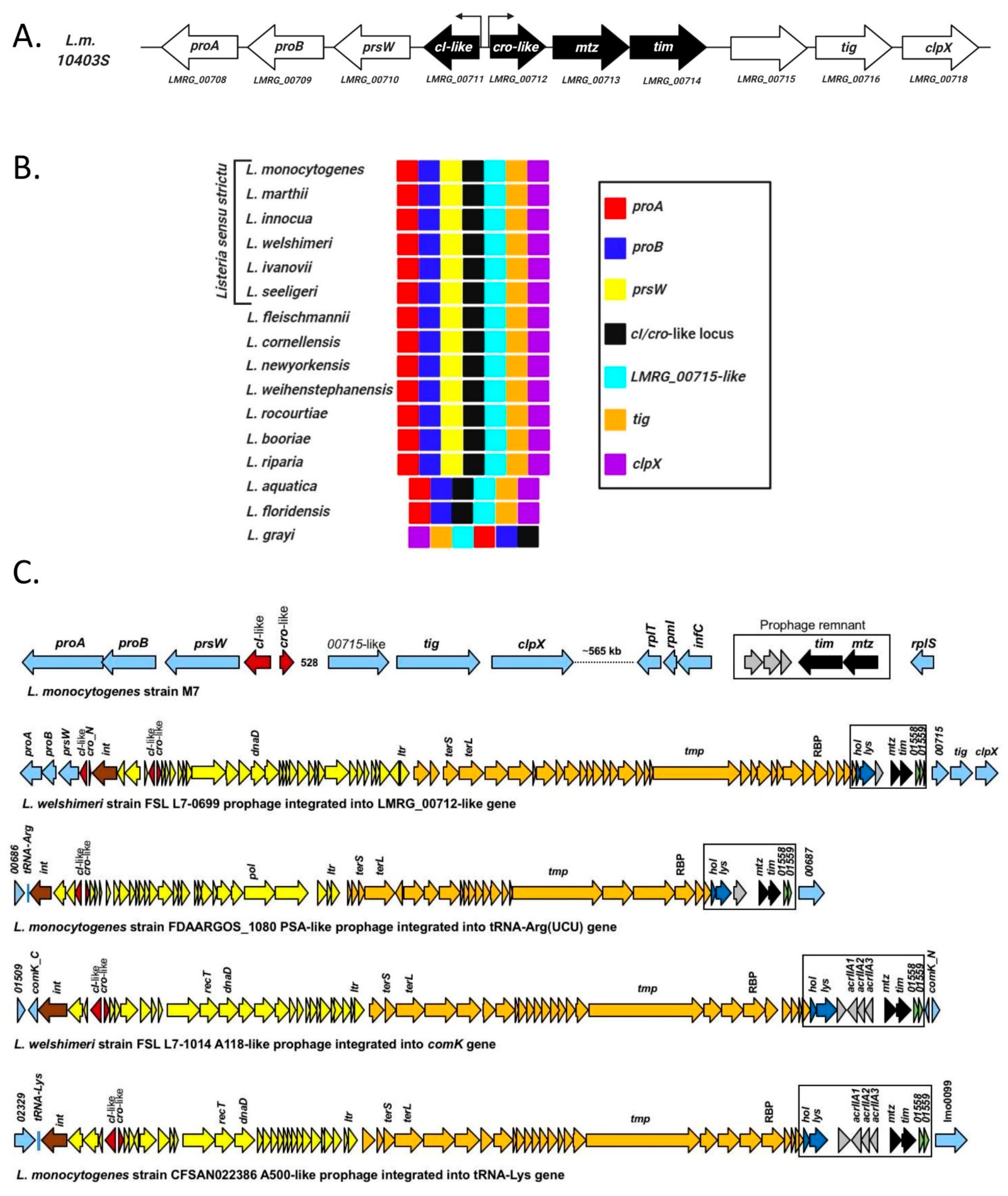
4.1. Listeria monocytogenes Harbors a Conserved Phage Regulatory Switch-Like Locus within Its Chromosome
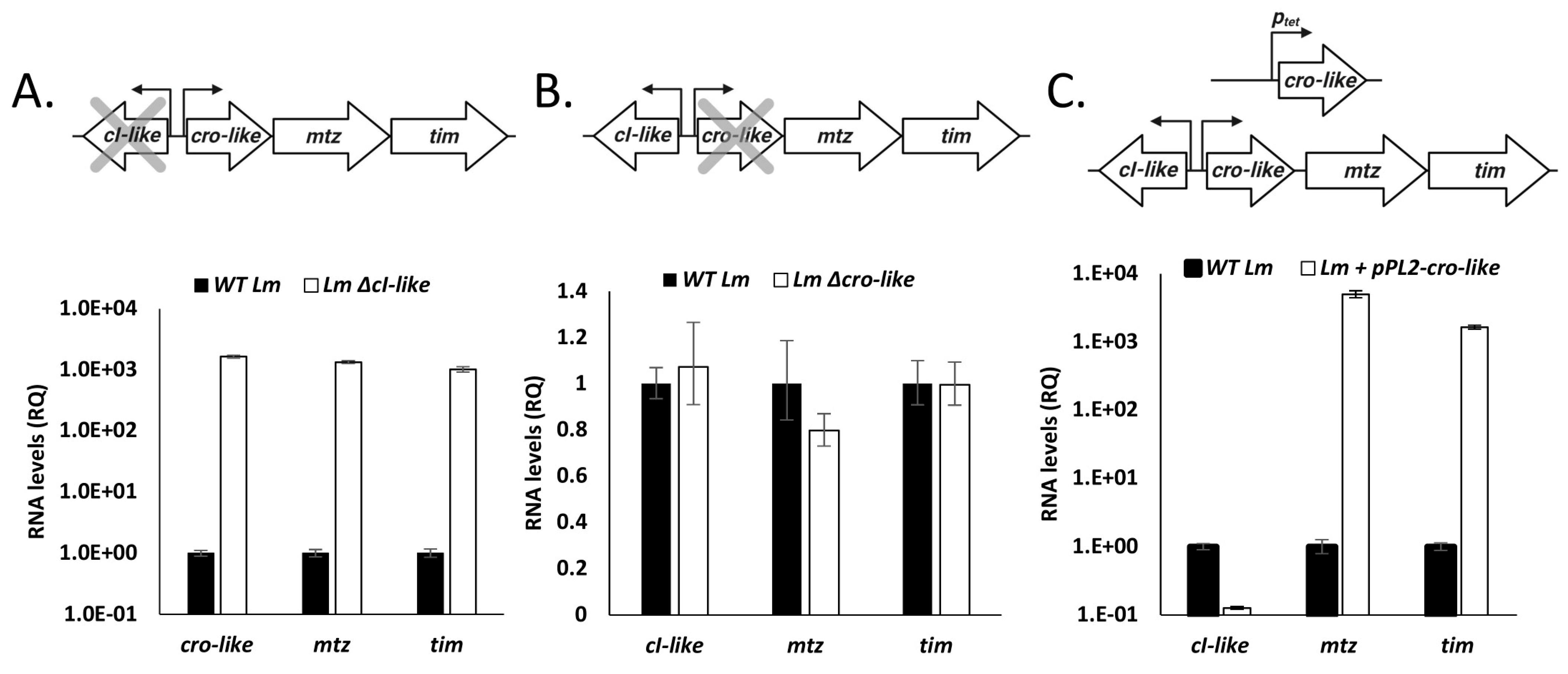
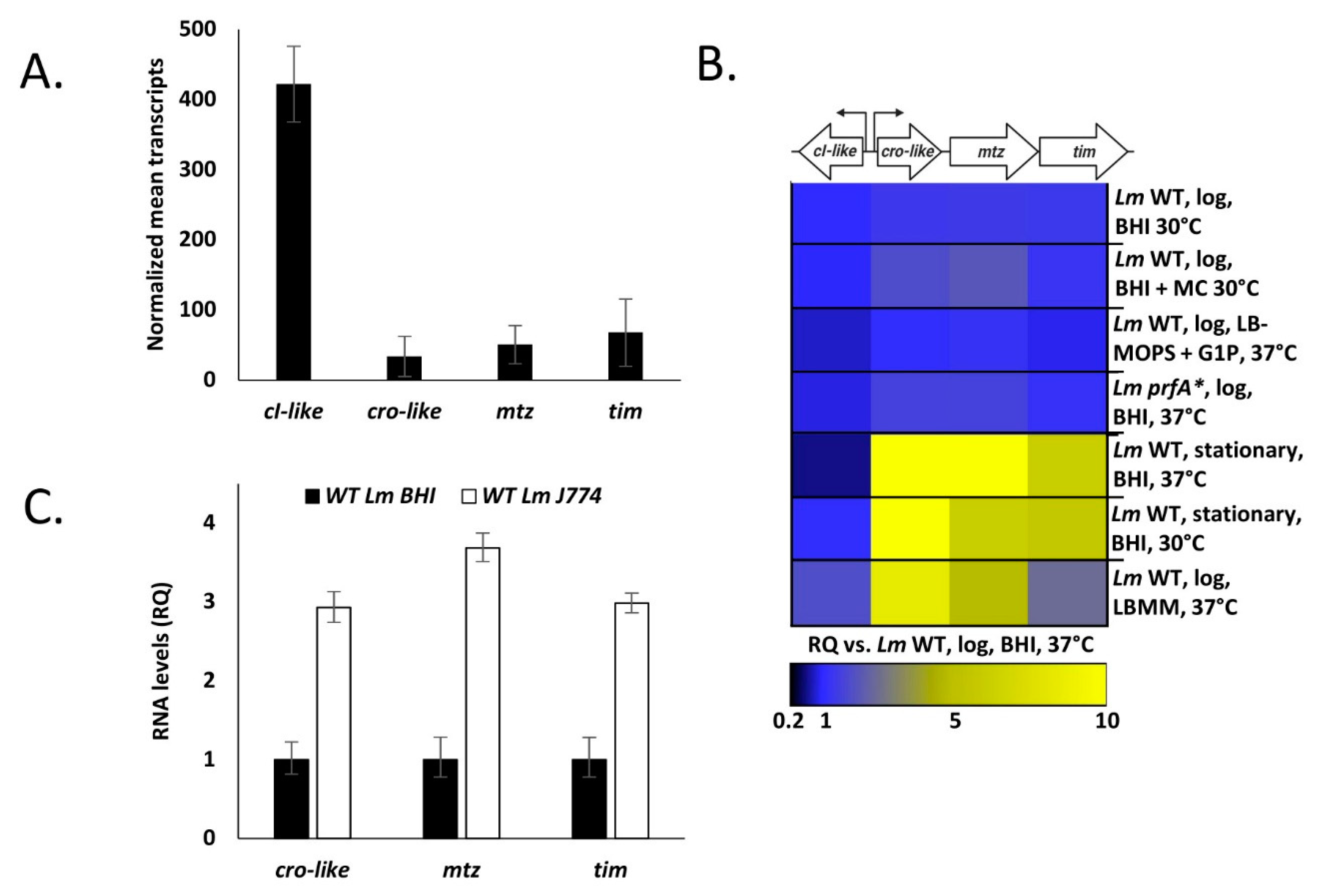
4.2. The cI-cro-Like Locus Possesses the Transcriptional Dynamics of a Regulatory Switch
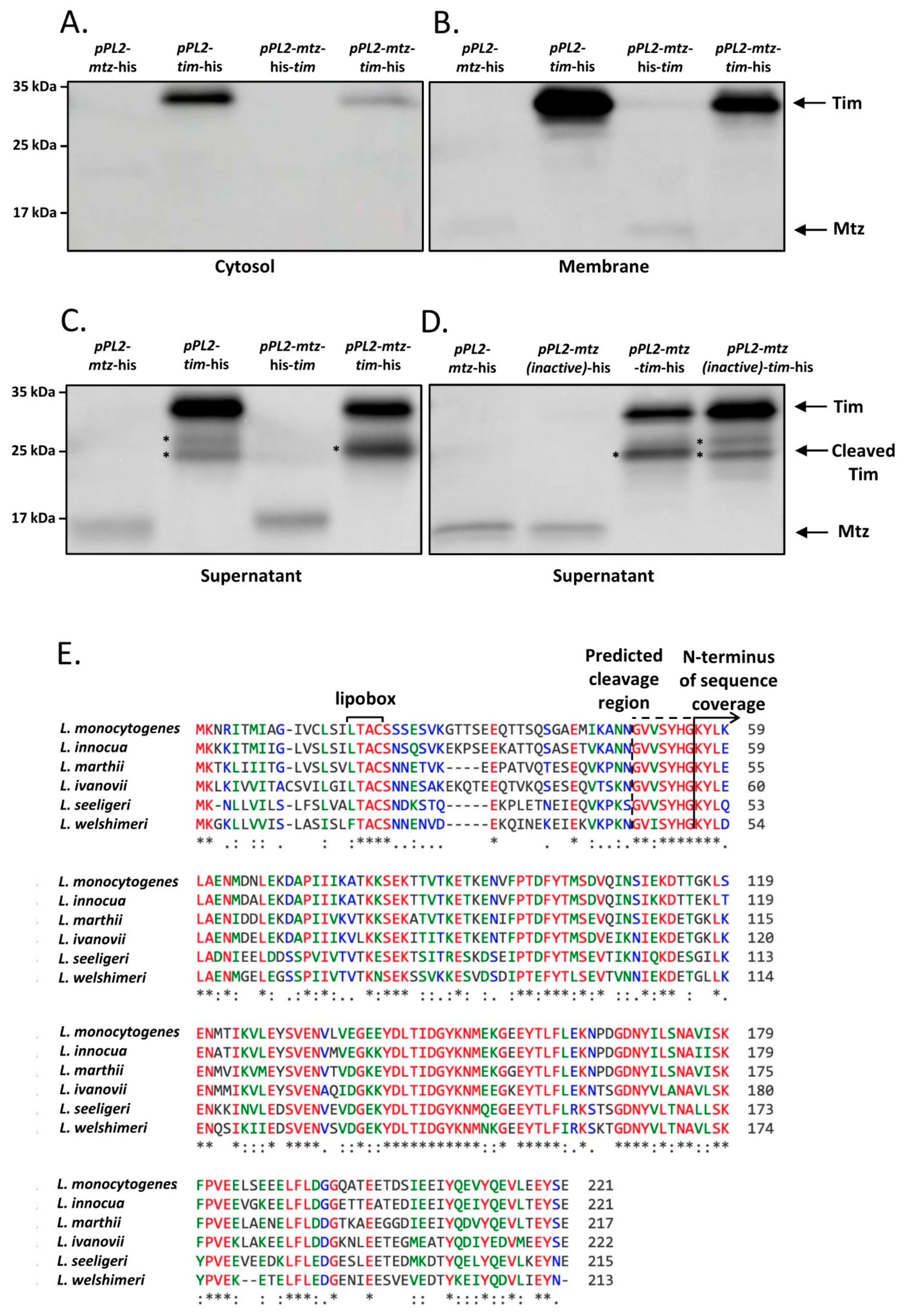
4.3. Mtz and Tim Localize to the Membrane and to the Culture Medium, Where Mtz Likely Cleaves Tim
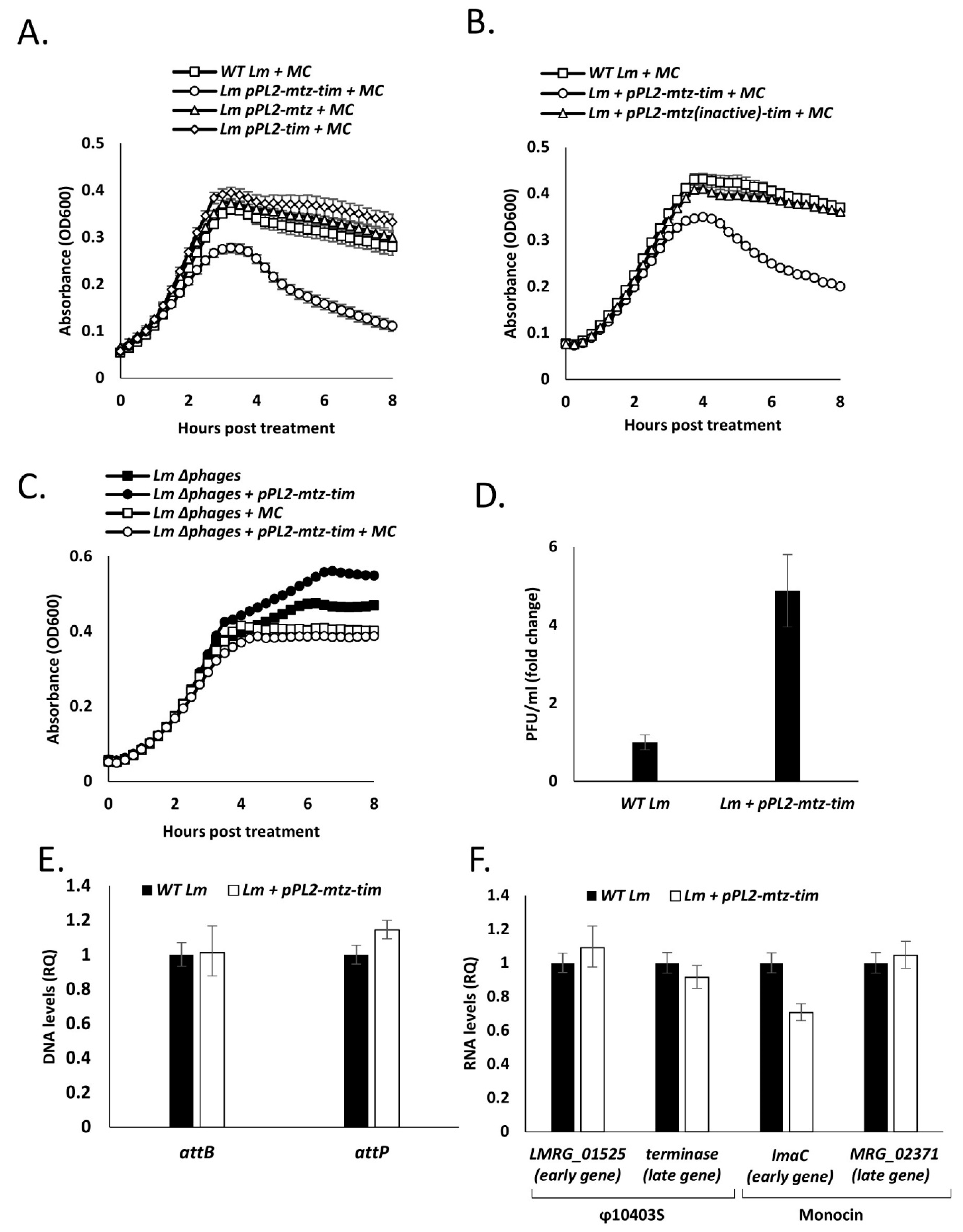
4.4. Expression of Both Mtz and Tim Sensitize the Bacteria to Phage Mediated Bacterial Lysis
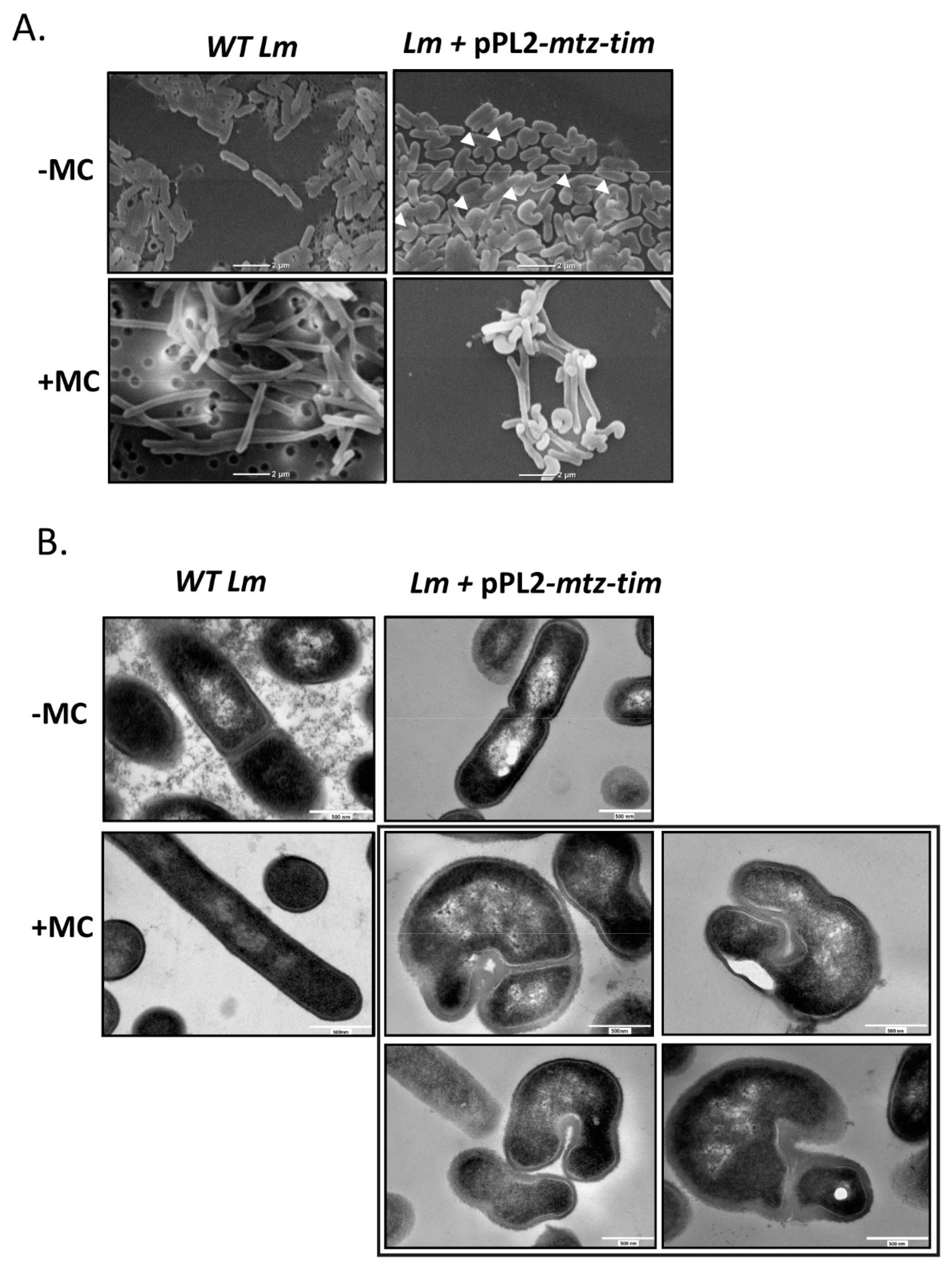
4.5. Expression of Mtz and Tim Changes Lm Cell Morphology
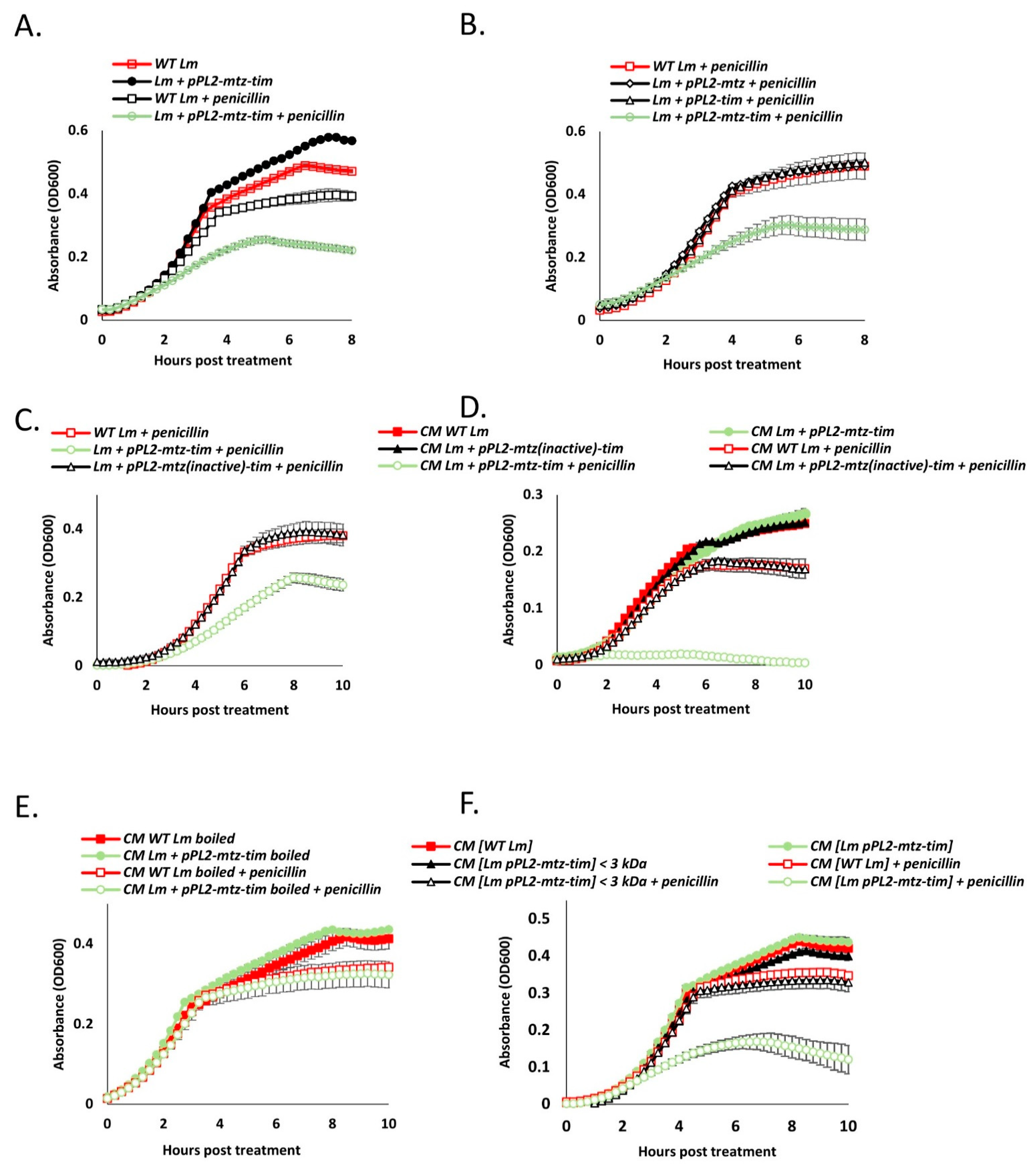
4.6. Secreted Mtz and Tim Increase Bacterial Susceptibility to Cell Wall Targeting Antibiotics, Also in Neighboring Cells
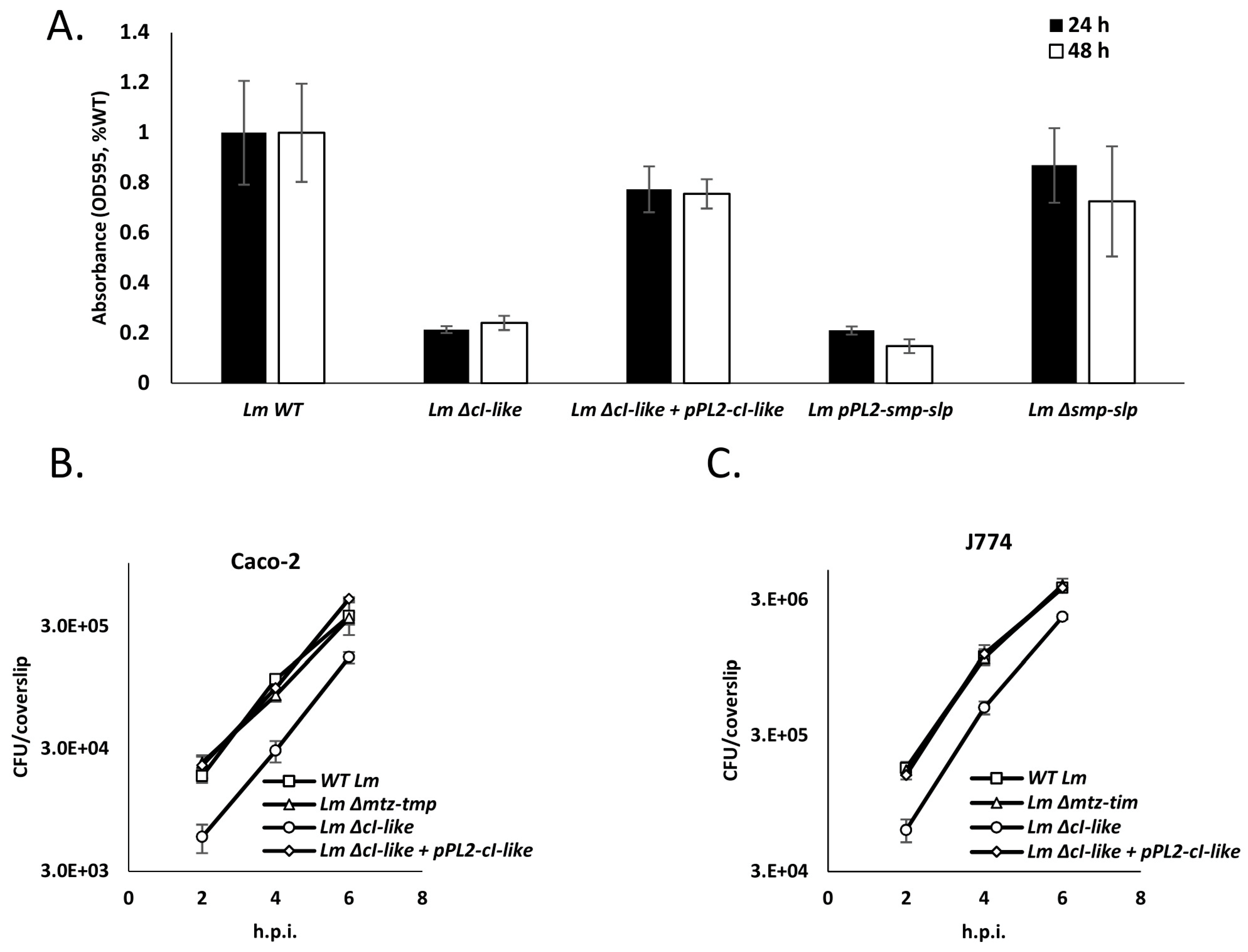
4.7. Expression of Mtz and Tim Hinders Lm Invasion of Mammalian Cells and Biofilm Formation
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef]
- de Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef]
- Breitbart, M.; Rohwer, F. Here a virus, there a virus, everywhere the same virus? Trends Microbiol. 2005, 13, 278–284. [Google Scholar] [CrossRef]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.D.; Buchan, A.; Finke, J.F.; Fuhrman, J.A.; Lennon, J.T.; Middelboe, M.; Suttle, C.A.; Stock, C.; et al. Re-examination of the relationship between marine virus and microbial cell abundances. Nat. Microbiol. 2016, 1. [Google Scholar] [CrossRef]
- Ofir, G.; Sorek, R. Contemporary Phage Biology: From Classic Models to New Insights. Cell 2018, 172, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, Z.; Abedon, S.T. Diversity of phage infection types and associated terminology: The problem with “Lytic or lysogenic”. FEMS Microbiol. Lett. 2016, 363, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Novak, K. A genetic switch. Nat. Rev. Cancer 2004, 4, 572. [Google Scholar] [CrossRef]
- Oppenheim, A.B.; Kobiler, O.; Stavans, J.; Court, D.L.; Adhya, S. Switches in Bacteriophage Lambda Development. Annu. Rev. Genet. 2005, 39, 409–429. [Google Scholar] [CrossRef]
- Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef]
- Loessner, M.J. Bacteriophage endolysins—Current state of research and applications. Curr. Opin. Microbiol. 2005, 8, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.M. Cryptic Prophages. J. Am. Coll. Radiol. JACR 2011, 10, 175–179. [Google Scholar]
- Rabinovich, L.; Sigal, N.; Borovok, I.; Nir-Paz, R.; Herskovits, A.A. Prophage excision activates listeria competence genes that promote phagosomal escape and virulence. Cell 2012, 150, 792–802. [Google Scholar] [CrossRef]
- Loessner, M.J.; Inman, R.B.; Lauer, P.; Calendar, R. Complete nucleotide sequence, molecular analysis and genome structure of bacteriophage A118 of Listeria monocytogenes: Implications for phage evolution. Mol. Microbiol. 2000, 35, 324–340. [Google Scholar] [CrossRef]
- Lee, G.; Chakraborty, U.; Gebhart, D.; Govoni, G.R.; Zhou, Z.H.; Scholl, D. F-type bacteriocins of Listeria monocytogenes: A new class of phage tail-like structures reveals broad parallel coevolution between tailed bacteriophages and high-molecular-weight bacteriocins. J. Bacteriol. 2016, 198, 2784–2793. [Google Scholar] [CrossRef]
- Zink, R.; Loessner, M.J.; Scherer, S. Characterization of cryptic prophages (monocins) in Listeria and sequence analysis of a holin/endolysin gene. Microbiology 1995, 141, 2577–2584. [Google Scholar] [CrossRef] [PubMed]
- Argov, T.; Azulay, G.; Pasechnek, A.; Stadnyuk, O.; Ran-Sapir, S.; Borovok, I.; Sigal, N.; Herskovits, A.A. Temperate bacteriophages as regulators of host behavior. Curr. Opin. Microbiol. 2017, 38, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Pasechnek, A.; Rabinovich, L.; Stadnyuk, O.; Azulay, G.; Mioduser, J.; Argov, T.; Borovok, I.; Sigal, N.; Herskovits, A.A. Active Lysogeny in Listeria Monocytogenes Is a Bacteria-Phage Adaptive Response in the Mammalian Environment. Cell Rep. 2020, 32, 107956. [Google Scholar] [CrossRef] [PubMed]
- Feiner, R.; Argov, T.; Rabinovich, L.; Sigal, N.; Borovok, I.; Herskovits, A.A. A new perspective on lysogeny: Prophages as active regulatory switches of bacteria. Nat. Rev. Microbiol. 2015, 13, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Argov, T.; Sapir, S.R.; Pasechnek, A.; Azulay, G.; Stadnyuk, O.; Rabinovich, L.; Sigal, N.; Borovok, I.; Herskovits, A.A. Coordination of cohabiting phage elements supports bacteria–phage cooperation. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Ripio, M.T.; Brehm, K.; Lara, M.; Suarez, M.; Vazquez-Boland, J.A. Glucose-1-phosphate utilization by Listeria monocytogenes is PrfA dependent and coordinately expressed with virulence factors. J. Bacteriol. 1997, 179, 7174–7180. [Google Scholar] [CrossRef]
- Lobel, L.; Sigal, N.; Borovok, I.; Ruppin, E.; Herskovits, A.A. Integrative Genomic Analysis Identifies Isoleucine and CodY as Regulators of Listeria monocytogenes Virulence. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.; Youngman, P. Use of a new integrational vector to investigate compartment-specific expression of the Bacillus subtilis spollM gene. Biochimie 1992, 74, 705–711. [Google Scholar] [CrossRef]
- Lauer, P.; Chow, M.Y.N.; Loessner, M.J.; Portnoy, D.A.; Calendar, R. Construction, characterization, and use of two Listeria monocytogenes site-specific phage integration vectors. J. Bacteriol. 2002, 184, 4177–4186. [Google Scholar] [CrossRef] [PubMed]
- Popowska, M.; Kusio, M.; Szymañska, P.; Markiewicz, Z. Inactivation of the wall-associated De-N-acetylase (PgdA) of Listeria monocytogenes results in greater susceptibility of the cells to induced autolysis. J. Microbiol. Biotechnol. 2009, 19, 932–945. [Google Scholar] [CrossRef] [PubMed]
- Sigal, N.; Pasechnek, A.; Herskovits, A.A. RNA purification from intracellularly grown listeria monocytogenes in macrophage cells. J. Vis. Exp. 2016, 2016, 54044. [Google Scholar] [CrossRef] [PubMed]
- Bode, W.; Grams, F.; Reinemer, P.; Gomis-Rüth, F.X.; Baumann, U.; McKay, D.B.; Stöcker, W. The metzincin-superfamily of zinc-peptidases. Adv. Exp. Med. Biol. 1996, 389, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Palmer, T.; Harrington, D.J.; Sutcliffe, I.C. Lipoprotein biogenesis in Gram-positive bacteria: Knowing when to hold ’em, knowing when to fold ’em. Trends Microbiol. 2009, 17, 13–21. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2016, 10, 845–858. [Google Scholar] [CrossRef]
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef]
- Schmid, M.W.; Ng, E.Y.W.; Lampidis, R.; Emmerth, M.; Walcher, M.; Kreft, J.; Goebel, W.; Wagner, M.; Schleifer, K.H. Evolutionary history of the genus Listeria and its virulence genes. Syst. Appl. Microbiol. 2005, 28, 1–18. [Google Scholar] [CrossRef]
- Rieu, A.; Weidmann, S.; Garmyn, D.; Piveteau, P.; Guzzo, J. Agr system of Listeria monocytogenes EGD-e: Role in adherence and differential expression pattern. Appl. Environ. Microbiol. 2007, 73, 6125–6133. [Google Scholar] [CrossRef]
- Ripio, M.T.; Domínguez-Bernal, G.; Lara, M.; Suárez, M.; Vázquez-Boland, J.A. A Gly145Ser substitution in the transcriptional activator PrfA causes constitutive overexpression of virulence factors in Listeria monocytogenes. J. Bacteriol. 1997, 179, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Lobel, L.; Sigal, N.; Borovok, I.; Belitsky, B.R.; Sonenshein, A.L.; Herskovits, A.A. The metabolic regulator CodY links Listeria monocytogenes metabolism to virulence by directly activating the virulence regulatory gene prfA. Mol. Microbiol. 2015, 95, 624–644. [Google Scholar] [CrossRef] [PubMed]
- McGwire, B.S.; Chang, K.P. Posttranslational regulation of a Leishmania HEXXH metalloprotease (gp63): The effects of site-specific mutagenesis of catalytic, zinc binding, N-glycosylation, and glycosyl phosphatidylinositol addition sites on N-terminal end cleavage, intracellular sta. J. Biol. Chem. 1996, 271, 7903–7909. [Google Scholar] [CrossRef] [PubMed]
- Klimpel, K.R.; Arora, N.; Leppla, S.H. Anthrax toxin lethal factor contains a zinc metalloprotease consensus sequence which is required for lethal toxin activity. Mol. Microbiol. 1994, 13, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Catalão, M.J.; Gil, F.; Moniz-Pereira, J.; São-José, C.; Pimentel, M. Diversity in bacterial lysis systems: Bacteriophages Show the way. FEMS Microbiol. Rev. 2013, 37, 554–571. [Google Scholar] [CrossRef] [PubMed]
- Korsak, D.; Markiewicz, Z.; Gutkind, G.O.; Ayala, J.A. Identification of the full set of Listeria monocytogenes penicillin-binding proteins and characterization of PBPD2 (Lmo2812). BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef]
- Guinane, C.M.; Cotter, P.D.; Ross, R.P.; Hill, C. Contribution of penicillin-binding protein homologs to antibiotic resistance, cell morphology, and virulence of Listeria monocytogenes EGDe. Antimicrob. Agents Chemother. 2006, 50, 2824–2828. [Google Scholar] [CrossRef]
- Tomasz, A. The mechanism of the irreversible antimicrobial effects of penicillins: How the beta-lactam antibiotics kill and lyse bacteria. Annu. Rev. Microbiol. 1979, 33, 113–137. [Google Scholar] [CrossRef]
- Aframian, N.; Eldar, A. A Bacterial Tower of Babel: Quorum-Sensing Signaling Diversity and Its Evolution. Annu. Rev. Microbiol. 2020, 74, 587–606. [Google Scholar] [CrossRef]
- Ouyang, Y.; Li, J.; Dong, Y.; Blakely, L.V.; Cao, M. Genome-wide screening of genes required for Listeria monocytogenes biofilm formation. J. Biotech Res. 2012, 4, 13–25. [Google Scholar]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef]
- Mira, A.; Ochman, H.; Moran, N.A. Deletional bias and the evolution of bacterial genomes. Trends Genet. 2001, 17, 589–596. [Google Scholar] [CrossRef]
- Lee, M.C.; Marx, C.J. Repeated, selection-driven genome reduction of accessory genes in experimental populations. PLoS Genet. 2012, 8, e1002651. [Google Scholar] [CrossRef]
- Matos, R.C.; Lapaque, N.; Rigottier-Gois, L.; Debarbieux, L.; Meylheuc, T.; Gonzalez-Zorn, B.; Repoila, F.; de Lopes, M.F.; Serror, P. Enterococcus faecalis Prophage Dynamics and Contributions to Pathogenic Traits. PLoS Genet. 2013, 9, e1003539. [Google Scholar] [CrossRef]
- Asadulghani, M.; Ogura, Y.; Ooka, T.; Itoh, T.; Sawaguchi, A.; Iguchi, A.; Nakayama, K.; Hayashi, T. The defective prophage pool of Escherichia coli O157: Prophage-prophage interactions potentiate horizontal transfer of virulence determinants. PLoS Pathog. 2009, 5, e1000408. [Google Scholar] [CrossRef]
- Michel-Briand, Y.; Baysse, C. The pyocins of Pseudomonas aeruginosa. Biochimie 2002, 84, 499–510. [Google Scholar] [CrossRef]
- Leiman, P.G.; Basler, M.; Ramagopal, U.A.; Bonanno, J.B.; Sauder, J.M.; Pukatzki, S.; Burley, S.K.; Almo, S.C.; Mekalanos, J.J. Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc. Natl. Acad. Sci. USA 2009, 106, 4154–4159. [Google Scholar] [CrossRef]
- Tree, J.J.; Granneman, S.; McAteer, S.P.; Tollervey, D.; Gally, D.L. Identification of Bacteriophage-Encoded Anti-sRNAs in Pathogenic Escherichia coli. Mol. Cell 2014, 55, 199–213. [Google Scholar] [CrossRef]
- Paul, J.H. Prophages in marine bacteria: Dangerous molecular time bombs or the key to survival in the seas? ISME J. 2008, 2, 579–589. [Google Scholar] [CrossRef]
- Duong, F.; Lazdunski, A.; Carni, B.; Murgier, M. Sequence of a cluster of genes controlling synthesis and secretion of alkaline protease in Pseudomonas aeruginosa: Relationships to other secretory pathways. Gene 1992, 121, 47–54. [Google Scholar] [CrossRef]
- Hege, T.; Feltzer, R.E.; Gray, R.D.; Baumann, U. Crystal structure of a complex between Pseudomonas aeruginosa alkaline protease and its cognate inhibitor: Inhibition by a zinc-NH2 coordinative bond. J. Biol. Chem. 2001, 276, 35087–35092. [Google Scholar] [CrossRef]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Gayathri, P.; Gunasekaran, P.; Jagannadham, M.V.; Rajendhran, J. Comprehensive proteomic analysis and pathogenic role of membrane vesicles of Listeria monocytogenes serotype 4b reveals proteins associated with virulence and their possible interaction with host. Int. J. Med. Microbiol. 2019, 309, 199–212. [Google Scholar] [CrossRef]
- Davis, K.L.; Wise, K.S. Site-specific proteolysis of the MALP-404 lipoprotein determines the release of a soluble selective lipoprotein-associated motif-containing fragment and alteration of the surface phenotype of Mycoplasma fermentans. Infect. Immun. 2002, 70, 1129–1135. [Google Scholar] [CrossRef]
- Antelmann, H.; Tjalsma, H.; Voigt, B.; Ohlmeier, S.; Bron, S.; Van Dijl, J.M.; Hecker, M. A proteomic view on genome-based signal peptide predictions. Genome Res. 2001, 11, 1484–1502. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.B.K.; Lampen, J.O. β-Lactamase III of Bacillus cereus 569: Membrane Lipoprotein and Secreted Protein. Biochemistry 1983, 22, 4652–4656. [Google Scholar] [CrossRef]
- Imanaka, T.; Oshihara, W.; Himeno, T.; Aiba, S. Comparative studies on extracellular penicillinases of the same structural gene, penP, expressed in Bacillus licheniformis and Bacillus subtilis. J. Gen. Microbiol. 1983, 129, 2621–2628. [Google Scholar] [CrossRef]
- Christopher, A.M.L.S. Integrative and Conjugative Elements (ICEs): What They Do and How They Work. Physiol. Behav. 2016, 176, 100–106. [Google Scholar] [CrossRef]
- Møretrø, T.; Langsrud, S. Listeria monocytogenes: Biofilm formation and persistence in food-processing environments. Biofilms 2004, 1, 107–121. [Google Scholar] [CrossRef]
- Hong, Y.; Ghebrehiwet, B. Effect of Pseudomonas aeruginosa elastase and alkaline protease on serum complement and isolated components C1q and C3. Clin. Immunol. Immunopathol. 1992, 62, 133–138. [Google Scholar] [CrossRef]
- Pel, M.J.C.; Van Dijken, A.J.H.; Bardoel, B.W.; Seidl, M.F.; Van Der Ent, S.; Van Strijp, J.A.G.; Pieterse, C.M.J. Pseudomonas syringae evades host immunity by degrading flagellin monomers with alkaline protease AprA. Mol. Plant Microbe Interact. 2014, 27, 603–610. [Google Scholar] [CrossRef]
- Parmely, M.; Gale, A.; Clabaugh, M.; Horvat, R.; Zhou, W.W. Proteolytic inactivation of cytokines by Pseudomonas aeruginosa. Infect. Immun. 1990, 58, 3009–3014. [Google Scholar] [CrossRef]
- Temple, M.E.; Nahata, M.C. Treatment of listeriosis. Ann. Pharmacother. 2000, 34, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.F.; Mobashery, S. β-Lactam resistance mechanisms: Gram-positive bacteria and mycobacterium tuberculosis. Cold Spring Harb. Perspect. Med. 2016, 6, 1–20. [Google Scholar] [CrossRef]
- Sanders, C.C.; Eugene Sanders, W. ß-lactam resistance in gram-negative bacteria: Global trends and clinical impact. Clin. Infect. Dis. 1992, 15, 824–839. [Google Scholar] [CrossRef]
- Aldridge, K.E.; Sanders, C.V.; Marier, R.L. Variation in the potentiation of/Hactam antibiotic activity by clavulanic add and solbactam against multiply antibiotic-resistant bacteria Chemical modification of the side chains of the/Mactams has produced compounds that are more resistant to/Mact. J. Antimicrob. Chemother. 1986, 17, 463–469. [Google Scholar] [CrossRef]
- Corbett, D.; Wise, A.; Langley, T.; Skinner, K.; Trimby, E.; Birchall, S.; Dorali, A.; Sandiford, S.; Williams, J.; Warn, P.; et al. Potentiation of antibiotic activity by a novel cationic peptide: Potency and spectrum of activity of SPR741. Antimicrob. Agents Chemother. 2017, 61, 1–10. [Google Scholar] [CrossRef]
- Abreu, A.C.; Serra, S.C.; Borges, A.; Saavedra, M.J.; Salgado, A.J.; Simões, M. Evaluation of the best method to assess antibiotic potentiation by phytochemicals against Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2014, 79, 125–134. [Google Scholar] [CrossRef][Green Version]
- Zanini, S.F.; Silva-Angulo, A.B.; Rosenthal, A.; Rodrigo, D.; Martínez, A. Effect of citral and carvacrol on the susceptibility of Listeria monocytogenes and Listeria innocua to antibiotics. Lett. Appl. Microbiol. 2014, 58, 486–492. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boichis, E.; Sigal, N.; Borovok, I.; Herskovits, A.A. A Metzincin and TIMP-Like Protein Pair of a Phage Origin Sensitize Listeria monocytogenes to Phage Lysins and Other Cell Wall Targeting Agents. Microorganisms 2021, 9, 1323. https://doi.org/10.3390/microorganisms9061323
Boichis E, Sigal N, Borovok I, Herskovits AA. A Metzincin and TIMP-Like Protein Pair of a Phage Origin Sensitize Listeria monocytogenes to Phage Lysins and Other Cell Wall Targeting Agents. Microorganisms. 2021; 9(6):1323. https://doi.org/10.3390/microorganisms9061323
Chicago/Turabian StyleBoichis, Etai, Nadejda Sigal, Ilya Borovok, and Anat A. Herskovits. 2021. "A Metzincin and TIMP-Like Protein Pair of a Phage Origin Sensitize Listeria monocytogenes to Phage Lysins and Other Cell Wall Targeting Agents" Microorganisms 9, no. 6: 1323. https://doi.org/10.3390/microorganisms9061323
APA StyleBoichis, E., Sigal, N., Borovok, I., & Herskovits, A. A. (2021). A Metzincin and TIMP-Like Protein Pair of a Phage Origin Sensitize Listeria monocytogenes to Phage Lysins and Other Cell Wall Targeting Agents. Microorganisms, 9(6), 1323. https://doi.org/10.3390/microorganisms9061323