
The Mechanism of Microbial-Ferromanganese Nodule Interaction and the Contribution of Biomineralization to the Formation of Oceanic Ferromanganese Nodules

Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Ferromanganese Nodules
2.3. Microbial-Ferromanganese Nodule Interaction Experiments
2.4. Analytical and Statistical Methods
3. Results
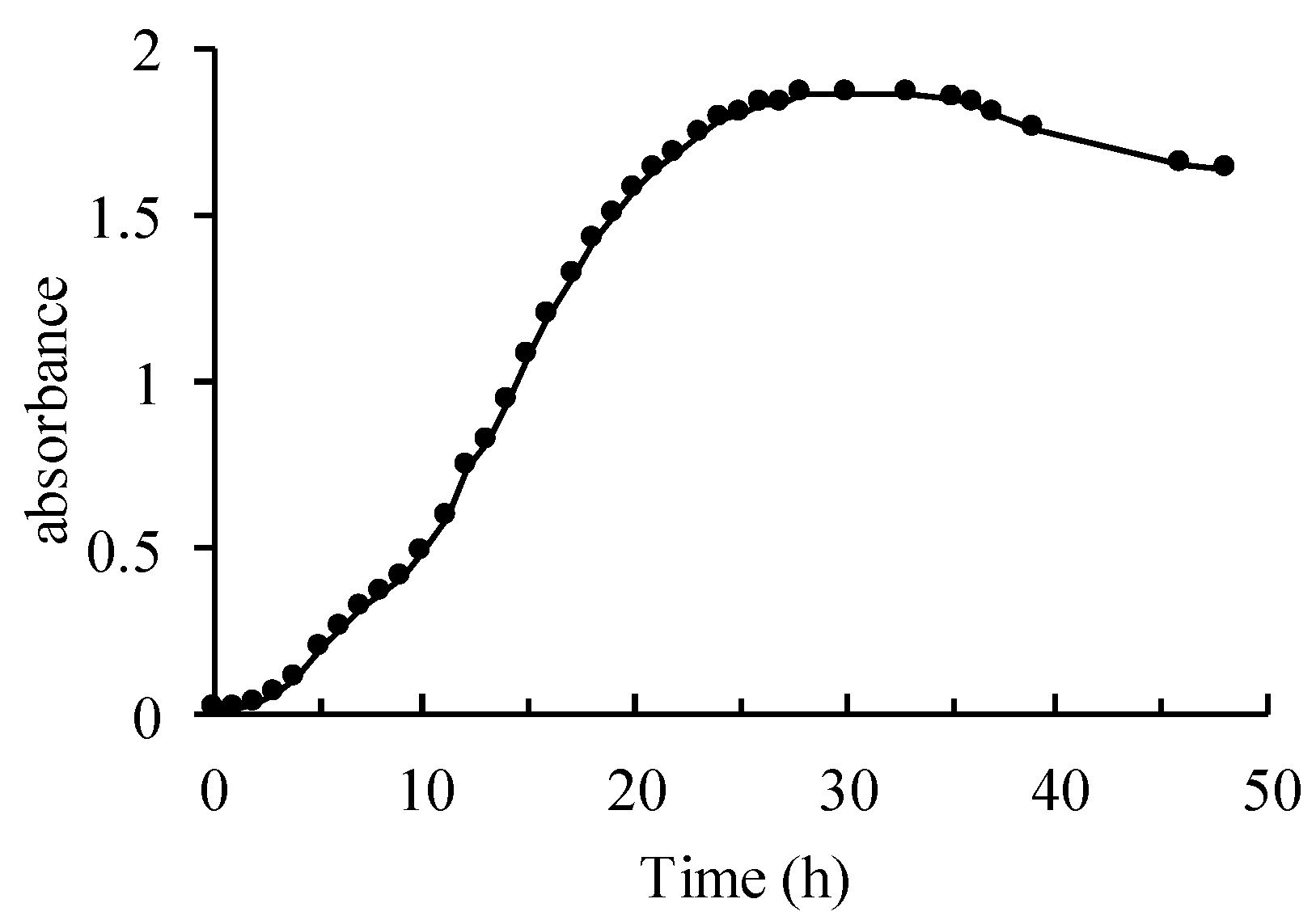
3.1. Growth Curve
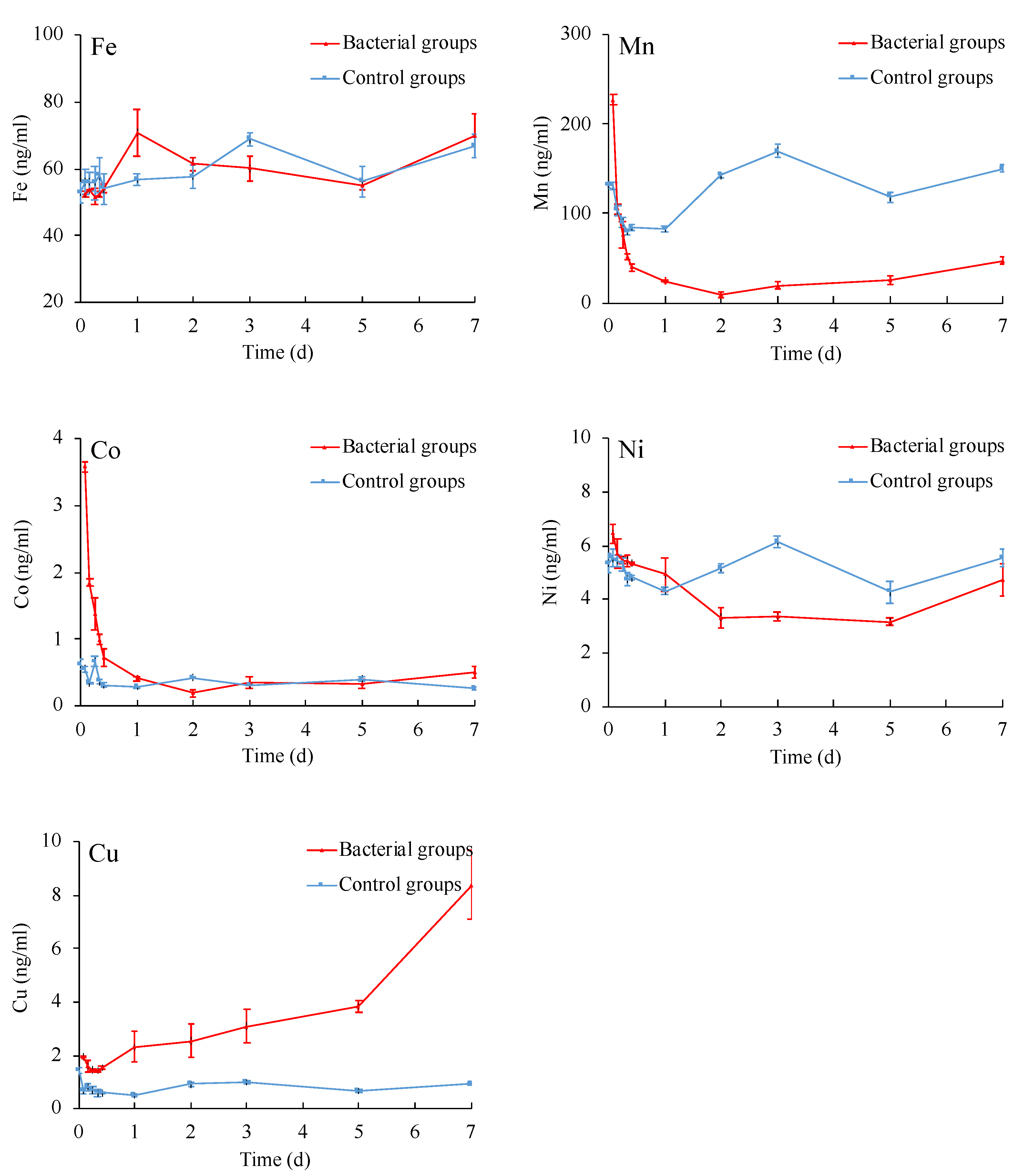
3.2. Elemental Changes of Ferromanganese Nodules
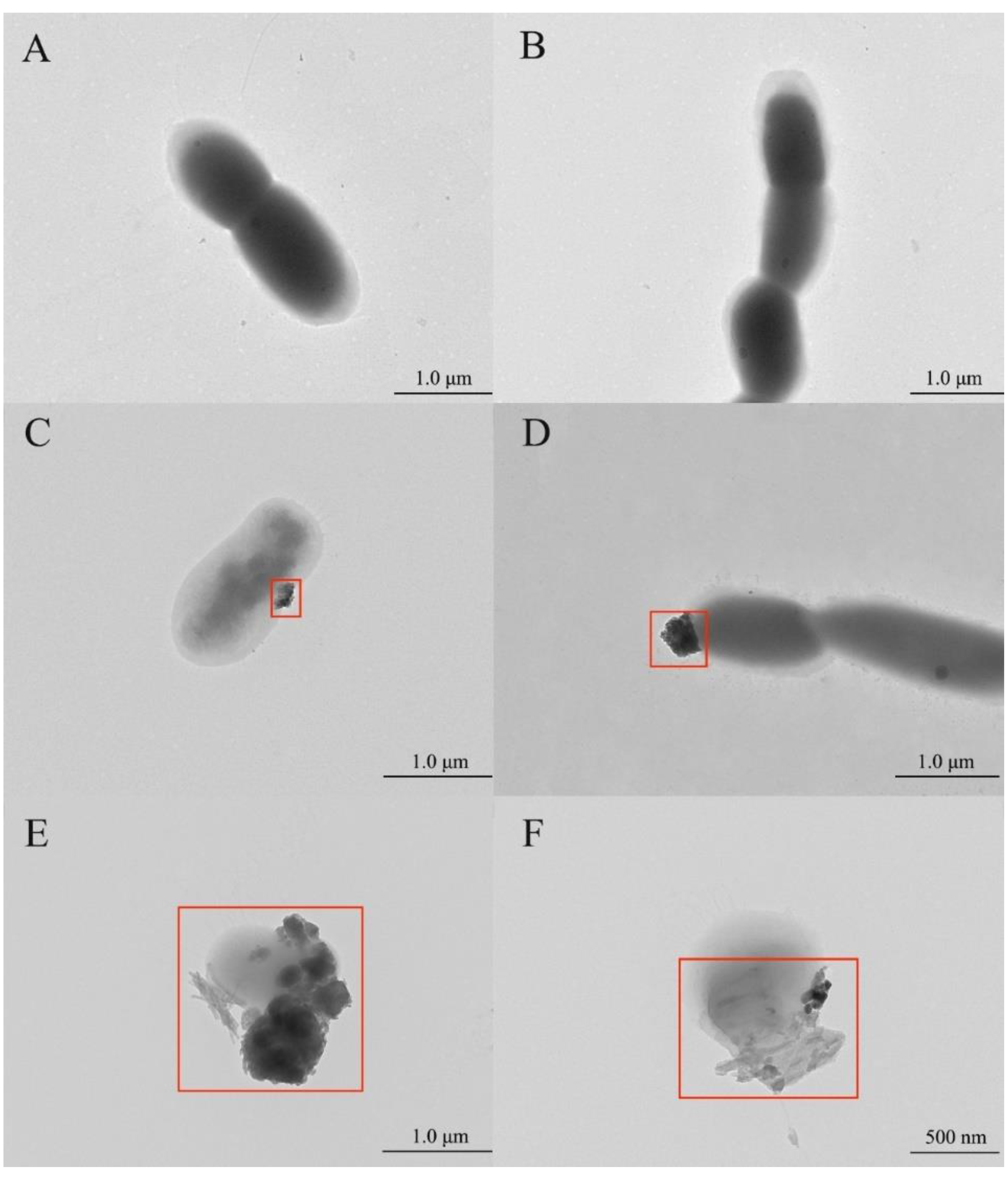
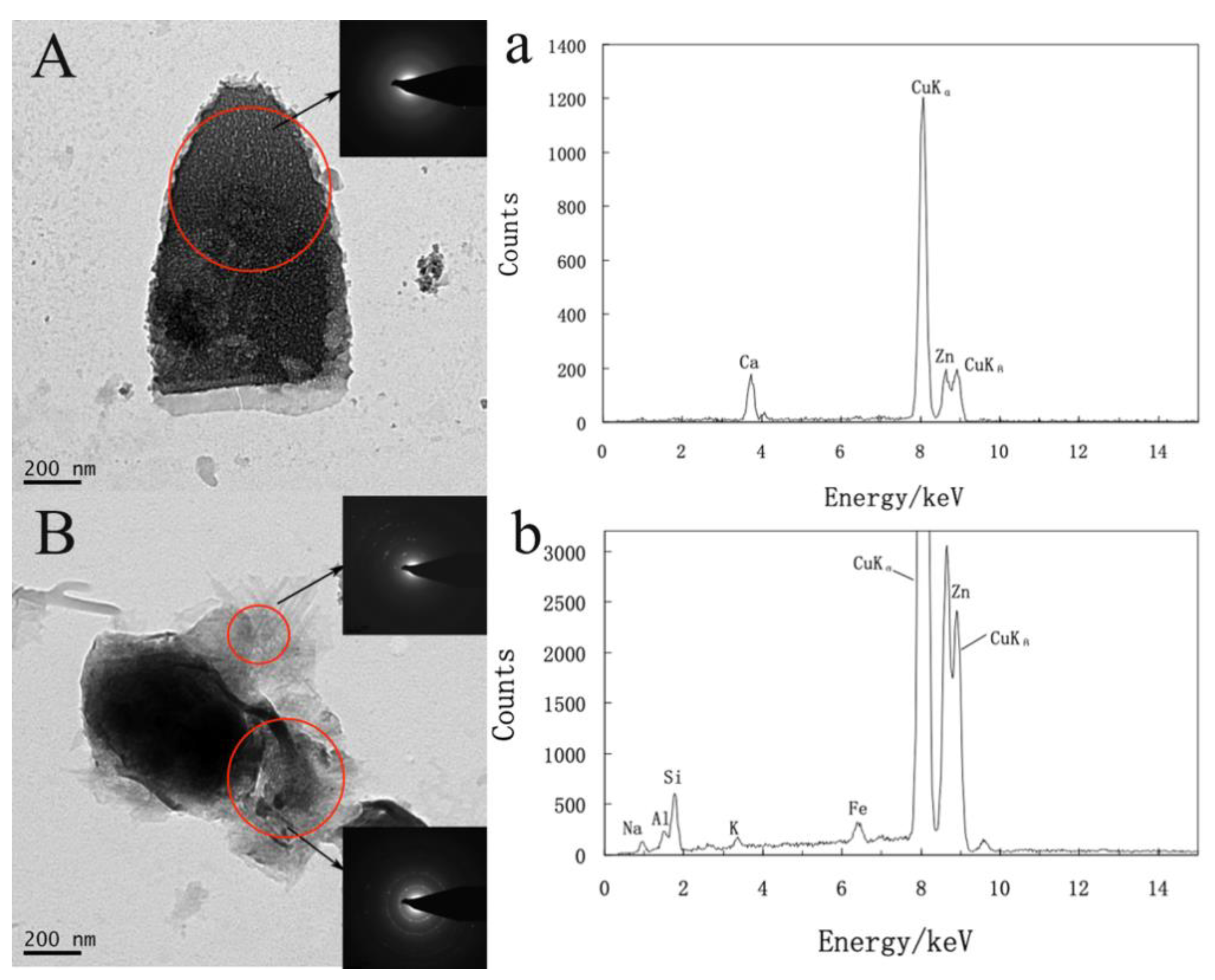
3.3. TEM Observation and Analysis of the Cells
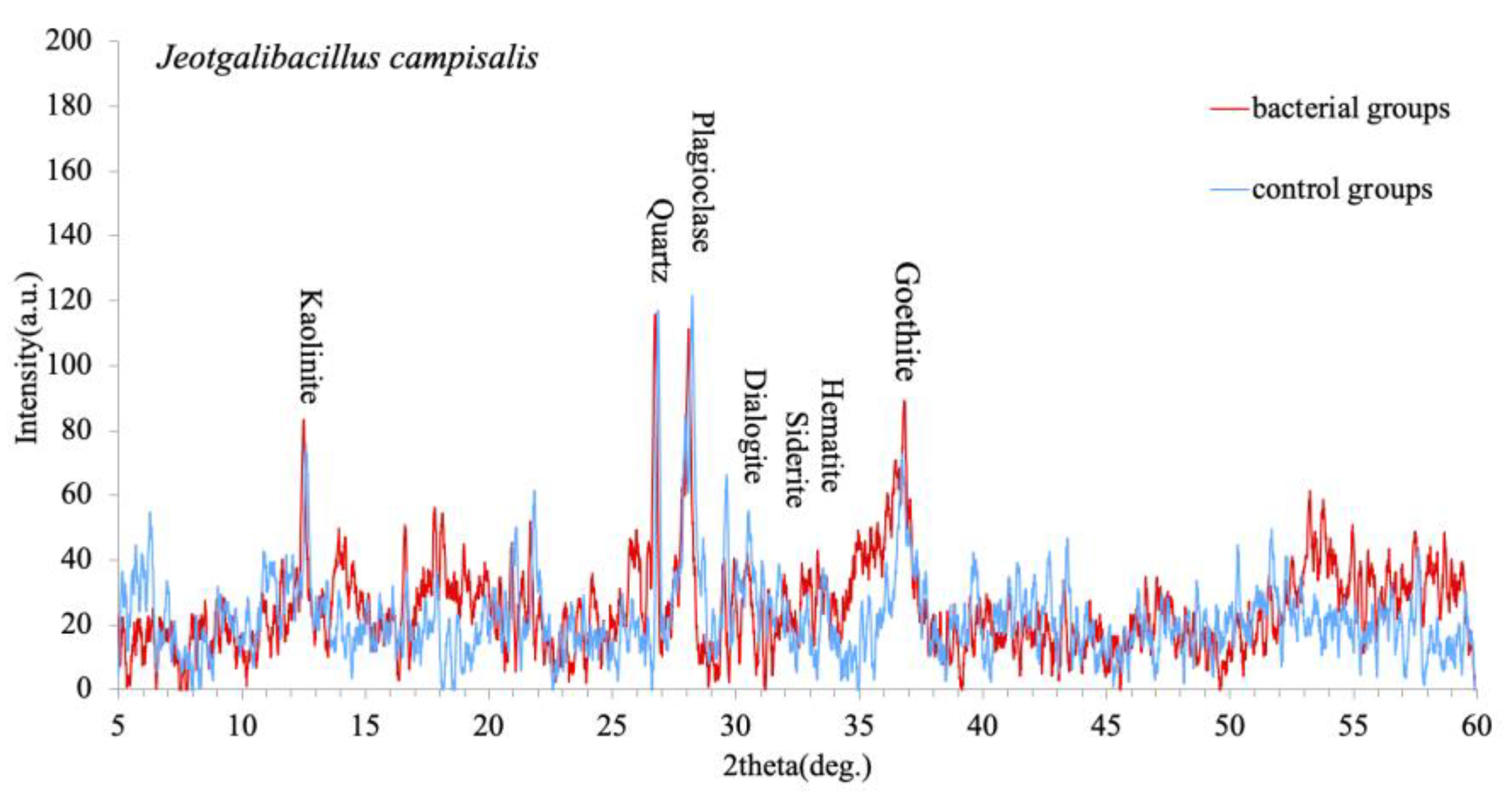
3.4. XRD Analysis of Ferromanganese Nodules Mineral Composition
4. Discussion
4.1. The Mechanism of Microbial-Ferromanganese Nodule Interaction
4.2. Biomineralization Promote the Formation of Ferromanganese Nodules
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hein, J.R.; Koschinsky, A. Deep-Ocean Ferromanganese Crusts and Nodules. In Treatise on Geochemistry; Elsevier Ltd.: Amsterdam, The Netherlands, 2014; pp. 273–291. [Google Scholar]
- Hein, J.R. Manganese Nodules. In Encyclopedia of Marine Geosciences; Harff, J., Meschede, M., Petersen, S., Thiede, J., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 408–412. [Google Scholar]
- Hein, J.R.; Mizell, K.; Koschinsky, A.; Conrad, T.A. Deep-Ocean Mineral Deposits As a Source of Critical Metals for High—and Green-Technology Applications: Comparison With Land-Based Resources. Ore Geol. Rev. 2013, 51, 1–14. [Google Scholar] [CrossRef]
- Verlaan, P.A.; Cronan, D.S.; Morgan, C.L. A comparative analysis of compositional variations in and between marine ferromanganese nodules and crusts in the South Pacific and their environmental controls. Prog. Oceanogr. 2004, 63, 125–158. [Google Scholar] [CrossRef]
- Koschinsky, A.; Halbach, P. Sequential leaching of marine ferromanganese precipitates: Genetic implications. Geochim. Cosmochim. Acta 1995, 59, 5113–5132. [Google Scholar] [CrossRef]
- Benites, M.; Millo, C.; Hein, J.; Nath, B.N.; Murton, B.; Galante, D.; Jovane, L. Integrated geochemical and morphological data provide insights into the genesis of ferromanganese nodules. Minerals 2018, 8, 488. [Google Scholar] [CrossRef]
- Thiel, G.A. Manganese precipitated by micro-organisms. Econ. Geol. 1925, 20, 301–310. [Google Scholar] [CrossRef]
- Shulse, C.N.; Maillot, B.; Smith, C.R.; Church, M.J. Polymetallic nodules, sediments, and deep waters in the equatorial North Pacific exhibit highly diverse and distinct bacterial, archaeal, and microeukaryotic communities. MicrobiologyOpen 2017, 6, e00428. [Google Scholar] [CrossRef]
- Ehrlich, H.L. How microbes influence mineral growth and dissolution. Chem. Geol. 1996, 132, 5–9. [Google Scholar] [CrossRef]
- Xie, S.; Liu, D.; Qiu, X.; Huang, X.; Algeo, T.J. Microbial roles equivalent to geological agents of high temperature and pressure in deep Earth. Sci. China Earth Sci. 2016, 59, 2098–2104. [Google Scholar] [CrossRef]
- Douglas, S.; Beveridge, T.J. Mineral formation by bacteria in natural microbial communities. FEMS Microbiol. Ecol. 1998, 26, 79–88. [Google Scholar] [CrossRef]
- Jiang, M.; Ohnuki, T.; Kozai, N.; Tanaka, K.; Suzuki, Y.; Sakamoto, F.; Kamiishi, E.; Utsunomiya, S. Biological nano-mineralization of Ce phosphate by Saccharomyces cerevisiae. Chem. Geol. 2010, 277, 61–69. [Google Scholar] [CrossRef]
- Jiang, M.; Ohnuki, T.; Tanaka, K.; Kozai, N.; Kamiishi, E.; Utsunomiya, S. Post-adsorption process of Yb phosphate nano-particle formation by Saccharomyces cerevisiae. Geochim. Cosmochim. Acta 2012, 93, 30–46. [Google Scholar] [CrossRef]
- Jiang, M.; Ohnuki, T.; Utsunomiya, S. Biomineralization of Middle Rare Earth Element Samarium in Yeast and Bacteria Systems. Geomicrobiol. J. 2018, 35, 375–384. [Google Scholar] [CrossRef]
- Ohnuki, T.; Jiang, M.; Sakamoto, F.; Kozai, N.; Yamasaki, S.; Yu, Q.; Tanaka, K.; Utsunomiya, S.; Xia, X.; Yang, K. Sorption of trivalent cerium by a mixture of microbial cells and manganese oxides: Effect of microbial cells on the oxidation of trivalent cerium. Geochim. Cosmochim. Acta 2015, 163, 1–13. [Google Scholar] [CrossRef]
- Braissant, O.; Decho, A.W.; Dupraz, C.; Glunk, C.; Przekop, K.M.; Visscher, P.T. Exopolymeric substances of sulfate-reducing bacteria: Interactions with calcium at alkaline pH and implication for formation of carbonate minerals. Geobiology 2007, 5, 401–411. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Frankel, R.B. Biologically controlled mineralization of magnetic iron minerals by magnetotactic bacteria. Environ. Microbe-Met. Interact. 2000, 109–144. [Google Scholar] [CrossRef]
- Rodelli, D.; Jovane, L.; Roberts, A.P.; Cypriano, J.; Abreu, F.; Lins, U. Fingerprints of partial oxidation of biogenic magnetite from cultivated and natural marine magnetotactic bacteria using synchrotron radiation. Environ. Microbiol. Rep. 2018, 10, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.B.; Rodelli, D.; Benites, M.; Abreu, F.; Murton, B.; Jovane, L. Presence of biogenic magnetite in ferromanganese nodules. Environ. Microbiol. Rep. 2020, 12, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Nitahara, S.; Kato, S.; Usui, A.; Urabe, T.; Suzuki, K.; Yamagishi, A. Archaeal and bacterial communities in deep-sea hydrogenetic ferromanganese crusts on old seamounts of the northwestern Pacific. PLoS ONE 2017, 12, e0173071. [Google Scholar] [CrossRef]
- Rosson, R.A.; Nealson, K.H. Manganese binding and oxidation by spores of a marine bacillus. J. Bacteriol. 1982, 151, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, S.; Tanner, S.; Bray, R. The molybdenum centre of native xanthine oxidase. Evidence for proton transfer from substrates to the centre and for existence of an anion-binding site. Biochem. J. 1978, 175, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Larock, P.A.; Ehrlich, H.L. Observations of bacterial microcolonies on the surface of ferromanganese nodules from Blake Plateau by scanning electron microscopy. Microb. Ecol. 1975, 2, 84–96. [Google Scholar] [CrossRef]
- Ehrlich, H.L. Geomicrobiology: Its significance for geology. Earth-Sci. Rev. 1998, 45, 45–60. [Google Scholar] [CrossRef]
- Melton, E.D.; Swanner, E.D.; Behrens, S.; Schmidt, C.; Kappler, A. The interplay of microbially mediated and abiotic reactions in the biogeochemical Fe cycle. Nat. Rev. Microbiol. 2014, 12, 797–808. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Kappler, A.; Roden, E.E. Iron in microbial metabolisms. Elements 2011, 7, 89–93. [Google Scholar] [CrossRef]
- Shiraishi, F.; Mitsunobu, S.; Suzuki, K.; Hoshino, T.; Morono, Y.; Inagaki, F. Dense microbial community on a ferromanganese nodule from the ultra-oligotrophic South Pacific Gyre: Implications for biogeochemical cycles. Earth Planet. Sci. Lett. 2016, 447, 10–20. [Google Scholar] [CrossRef]
- Tully, B.J.; Heidelberg, J.F. Microbial communities associated with ferromanganese nodules and the surrounding sediments. Front. Microbiol. 2013, 4, 161. [Google Scholar] [CrossRef] [PubMed]
- Blöthe, M.; Wegorzewski, A.; Müller, C.; Simon, F.; Kuhn, T.; Schippers, A. Manganese-cycling microbial communities inside deep-sea manganese nodules. Environ. Sci. Technol. 2015, 49, 7692–7700. [Google Scholar] [CrossRef] [PubMed]
- Lindh, M.V.; Maillot, B.M.; Shulse, C.N.; Gooday, A.J.; Amon, D.J.; Smith, C.R.; Church, M.J. From the surface to the deep-sea: Bacterial distributions across polymetallic nodule fields in the clarion-clipperton zone of the Pacific Ocean. Front. Microbiol. 2017, 8, 1696. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.; Das, S.K.; Munda, P. Biogenic signature and ultra microfossils in ferromanganese nodules of the Central Indian Ocean Basin. J. Asian Earth Sci. 2013, 73, 296–305. [Google Scholar] [CrossRef]
- Jiang, X.-D.; Sun, X.-M.; Guan, Y. Biogenic mineralization in the ferromanganese nodules and crusts from the South China Sea. J. Asian Earth Sci. 2019, 171, 46–59. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Kang, S.-J.; Schumann, P.; Oh, T.-K. Jeotgalibacillus salarius sp. nov., isolated from a marine saltern, and reclassification of Marinibacillus marinus and Marinibacillus campisalis as Jeotgalibacillus marinus comb. nov. and Jeotgalibacillus campisalis comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2010, 60, 15–20. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yoon, J.-H.; Kim, I.-G.; Schumann, P.; Oh, T.-K.; Park, Y.-H. Marinibacillus campisalis sp. nov., a moderate halophile isolated from a marine solar saltern in Korea, with emended description of the genus Marinibacillus. Int. J. Syst. Evol. Microbiol. 2004, 54, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.-R.; Lu, D.-C.; Sun, X.-K.; Sun, Y.-Y.; Saren, G.; Yu, X.-K.; Du, Z.-J. Seonamhaeicola maritimus sp. nov., isolated from coastal sediment. Int. J. Syst. Evol. Microbiol. 2020, 70, 902–908. [Google Scholar] [CrossRef]
- Schwertmann, U. Transformation of hematite to goethite in soils. Nature 1971, 232, 624–625. [Google Scholar] [CrossRef]
- Stumm, W.; Morgan, J. Aquatic Chemistry, Chemical Equilibria and Rates in Natural Waters; John Wiley & Sons, Inc.: New York, NY, USA, 1996. [Google Scholar]
- Albery, W.J.; Knowles, J.R. Evolution of enzyme function and the development of catalytic efficiency. Biochemistry 1976, 15, 5631–5640. [Google Scholar] [CrossRef]
- Ferris, F. Microbe-Metal Interactions in Sediments. In Microbial Sediments; Springer: Berlin/Heidelberg, Germany, 2000; pp. 121–126. [Google Scholar]
- Baker, M.; Lalonde, S.; Konhauser, K.; Foght, J. Role of extracellular polymeric substances in the surface chemical reactivity of Hymenobacter aerophilus, a psychrotolerant bacterium. Appl. Environ. Microbiol. 2010, 76, 102–109. [Google Scholar] [CrossRef]
- Roberts, J.A.; Fowle, D.A.; Hughes, B.T.; Kulczycki, E. Attachment behavior of Shewanella putrefaciens onto magnetite under aerobic and anaerobic conditions. Geomicrobiol. J. 2006, 23, 631–640. [Google Scholar] [CrossRef]
- Maurice, P.A.; Haack, E.A.; Mishra, B. Siderophore sorption to clays. Biometals 2009, 22, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, O.W.; Sposito, G. Siderophore-promoted dissolution of synthetic and biogenic layer-type Mn oxides. Chem. Geol. 2007, 242, 497–508. [Google Scholar] [CrossRef]
- Peña, J.; Duckworth, O.W.; Bargar, J.R.; Sposito, G. Dissolution of hausmannite (Mn3O4) in the presence of the trihydroxamate siderophore desferrioxamine B. Geochim. Cosmochim. Acta 2007, 71, 5661–5671. [Google Scholar] [CrossRef]
- Welch, S.; Barker, W.; Banfield, J. Microbial extracellular polysaccharides and plagioclase dissolution. Geochim. Cosmochim. Acta 1999, 63, 1405–1419. [Google Scholar] [CrossRef]
- Barker, W.; Welch, S.; Chu, S.; Banfield, J. Experimental observations of the effects of bacteria on aluminosilicate weathering. Am. Mineral. 1998, 83, 1551–1563. [Google Scholar] [CrossRef]
- Kendall, B.; Anbar, A.D.; Kappler, A.; Konhauser, K.O. The global iron cycle. Fundam. Geobiol. 2012, 1, 65–92. [Google Scholar]
- Holland, H.D. The oceans; a possible source of iron in iron-formations. Econ. Geol. 1973, 68, 1169–1172. [Google Scholar] [CrossRef]
- Morris, R. Genetic modelling for banded iron-formation of the Hamersley Group, Pilbara Craton, Western Australia. Precambrian Res. 1993, 60, 243–286. [Google Scholar] [CrossRef]
- Xiao-quan, S.; Radziuk, B.; Welz, B.; Vyskočilová, O. Determination of manganese in river and sea-water samples by electrothermal atomic absorption spectrometry with a tungsten atomizer. J. Anal. At. Spectrom. 1993, 8, 409–413. [Google Scholar] [CrossRef]
- Anthoni, J.F. The chemical composition of seawater. Magnesium 2006, 2701, 96-062. [Google Scholar]
- Nielsen, A.E.; Söhnel, O. Interfacial tensions electrolyte crystal-aqueous solution, from nucleation data. J. Cryst. Growth 1971, 11, 233–242. [Google Scholar] [CrossRef]
- Konhauser, K.O. Introduction to Geomicrobiology; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Konhauser, K.; Riding, R. Bacterial biomineralization. In Fundamentals of Geobiology; Wiley Online Library: Hoboken, NJ, USA, 2012; pp. 105–130. [Google Scholar]
- Dove, P.; De Yoreo, J.; Weiner, S. (Eds.) Biomineralization; Mineralogical Society of America: Washington, DC, USA, 2003. [Google Scholar]
- Beveridge, T.; Murray, R. Uptake and retention of metals by cell walls of Bacillus subtilis. J. Bacteriol. 1976, 127, 1502–1518. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, T.; Murray, R. Sites of metal deposition in the cell wall of Bacillus subtilis. J. Bacteriol. 1980, 141, 876–887. [Google Scholar] [CrossRef]
- Kashefi, K.; Lovley, D.R. Reduction of Fe (III), Mn (IV), and toxic metals at 100 °C by Pyrobaculum islandicum. Appl. Environ. Microbiol. 2000, 66, 1050–1056. [Google Scholar] [CrossRef]
- Wang, X.-H.; Gan, L.; Müller, W.E. Contribution of biomineralization during growth of polymetallic nodules and ferromanganese crusts from the Pacific Ocean. Front. Mater. Sci. China 2009, 3, 109–123. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Hamade, T.; Raiswell, R.; Morris, R.C.; Ferris, F.G.; Southam, G.; Canfield, D.E. Could bacteria have formed the Precambrian banded iron formations? Geology 2002, 30, 1079–1082. [Google Scholar] [CrossRef]
- Kappler, A.; Pasquero, C.; Konhauser, K.O.; Newman, D.K. Deposition of banded iron formations by anoxygenic phototrophic Fe (II)-oxidizing bacteria. Geology 2005, 33, 865–868. [Google Scholar] [CrossRef]
- Posth, N.R.; Hegler, F.; Konhauser, K.O.; Kappler, A. Alternating Si and Fe deposition caused by temperature fluctuations in Precambrian oceans. Nat. Geosci. 2008, 1, 703–708. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Si | Fe | Al | Ca | Mg | Na | K | Mn | Cl | S |
|---|---|---|---|---|---|---|---|---|---|---|
| Content (PPM) | 194,180 | 202,440 | 24,122.4 | 30,773.4 | 9120 | 8904 | 5808.6 | 104,174.6 | 132 | 229.1 |
| Element | Ti | P | Ni | Co | Ba | Pb | Ce | Sr | Cu | V |
| Content (PPM) | 10,483.9 | 4431.9 | 2117.8 | 2125.4 | 321 | 1277.7 | 1285.3 | 1098 | 1285.3 | 764.2 |
| Element | Zr | Zn | As | Nd | La | Mo | Y | Cr | W | Nb |
| Content (PPM) | 541 | 496.8 | 489.6 | 445.9 | 321 | 297 | 160.6 | 83 | 67.2 | 45.6 |
| Element | Th | Sb | Hf | Cd | Bi | Rb | Sc | Ga | U | Br |
| Content (PPM) | 42.1 | 12 | 14.8 | — | 8.4 | 7.2 | 4.5 | 2.7 | 0.1 | 0 |
| Fe | Mn | Co | Ni | Cu |
|---|---|---|---|---|
| 0.4922 ns | 0.04883 * | 0.03654 * | 0.625 ns | 0.001953 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, J.; Yu, X.; Jiang, M.; Cao, W.; Saren, G.; Chang, F. The Mechanism of Microbial-Ferromanganese Nodule Interaction and the Contribution of Biomineralization to the Formation of Oceanic Ferromanganese Nodules. Microorganisms 2021, 9, 1247. https://doi.org/10.3390/microorganisms9061247
Lyu J, Yu X, Jiang M, Cao W, Saren G, Chang F. The Mechanism of Microbial-Ferromanganese Nodule Interaction and the Contribution of Biomineralization to the Formation of Oceanic Ferromanganese Nodules. Microorganisms. 2021; 9(6):1247. https://doi.org/10.3390/microorganisms9061247
Chicago/Turabian StyleLyu, Jing, Xinke Yu, Mingyu Jiang, Wenrui Cao, Gaowa Saren, and Fengming Chang. 2021. "The Mechanism of Microbial-Ferromanganese Nodule Interaction and the Contribution of Biomineralization to the Formation of Oceanic Ferromanganese Nodules" Microorganisms 9, no. 6: 1247. https://doi.org/10.3390/microorganisms9061247
APA StyleLyu, J., Yu, X., Jiang, M., Cao, W., Saren, G., & Chang, F. (2021). The Mechanism of Microbial-Ferromanganese Nodule Interaction and the Contribution of Biomineralization to the Formation of Oceanic Ferromanganese Nodules. Microorganisms, 9(6), 1247. https://doi.org/10.3390/microorganisms9061247