
The Effects of Chemical and Mechanical Stresses on Bacillus cereus and Pseudomonas fluorescens Single- and Dual-Species Biofilm Removal

 ,
,  ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria and Culture Conditions
2.2. Substratum
2.3. Characterisation of Substrate and Bacterial Surfaces
2.3.1. Surface Charge—Zeta Potential
2.3.2. Physicochemical Characterisation: Hydrophobicity and Surface Tension
2.4. Free Energy of Adhesion
2.5. Adhesion Assays
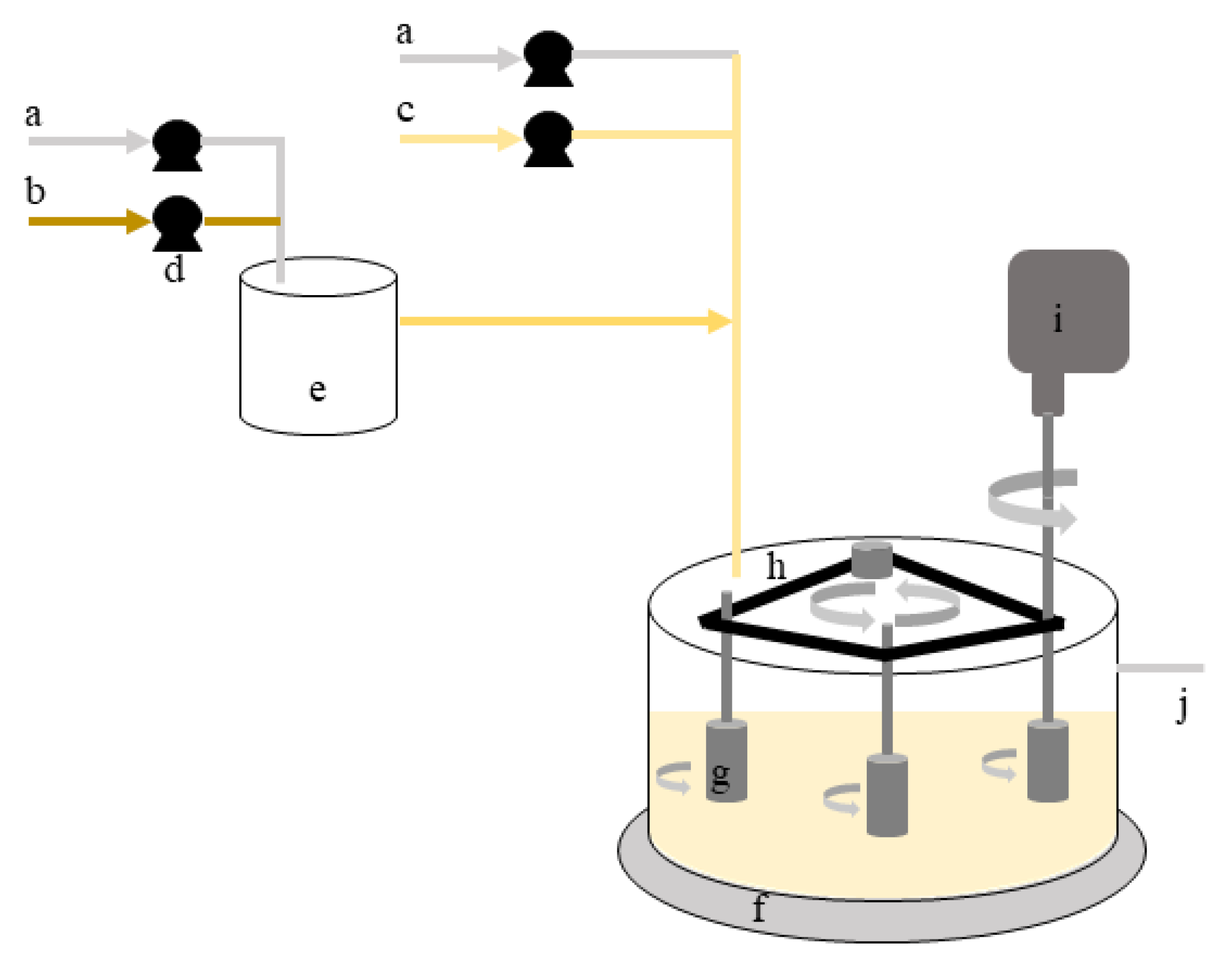
2.6. Biofilm Formation–Experimental Set-Up
2.7. Biofilm Characterization
2.8. Chemical Treatment
2.9. Mechanical Treatment
2.10. Statistical Analysis
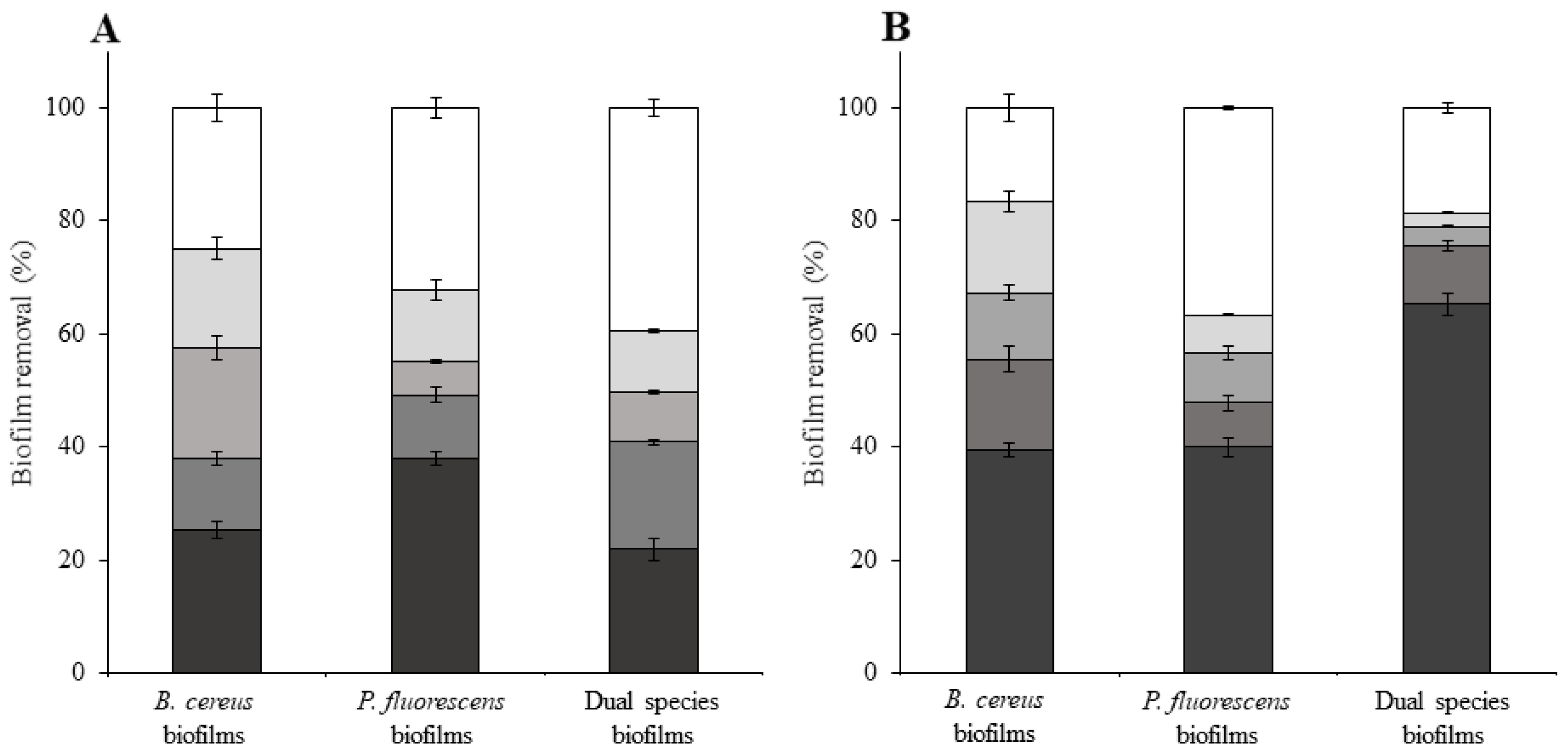
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the Food Industry: Health Aspects and Control Methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef] [PubMed]
- Gomes, I.B.; Simões, M.; Simões, L.C. An overview on the reactors to study drinking water biofilms. Water Res. 2014, 62, 63–87. [Google Scholar] [CrossRef] [PubMed]
- da Silva Fernandes, M.; Kabuki, D.Y.; Kuaye, A.Y. Behavior of Listeria monocytogenes in a multi-species biofilm with Enterococcus faecalis and Enterococcus faecium and control through sanitation procedures. Int. J. Food Microbiol. 2015, 200, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; Gomes, I.B.; Simões, L.C.; Simões, M. Overview on the hydrodynamic conditions found in industrial systems and its impact in (bio)fouling formation. Chem. Eng. J. 2021, 418, 129348. [Google Scholar] [CrossRef]
- Abe, Y.; Skali-Lami, S.; Block, J.C.; Francius, G. Cohesiveness and hydrodynamic properties of young drinking water biofilms. Water Res. 2012, 46, 1155–1166. [Google Scholar] [CrossRef]
- Gomes, I.B.; Lemos, M.; Mathieu, L.; Simões, M.; Simões, L.C. The action of chemical and mechanical stresses on single and dual species biofilm removal of drinking water bacteria. Sci. Total Environ. 2018, 631–632, 987–993. [Google Scholar] [CrossRef]
- Simões, M.; Simões, L.C.; Vieira, M.J. Species association increases biofilm resistance to chemical and mechanical treatments. Water Res. 2009, 43, 229–237. [Google Scholar] [CrossRef]
- Douterelo, I.; Sharpe, R.L.; Boxall, J.B. Influence of hydraulic regimes on bacterial community structure and composition in an experimental drinking water distribution system. Water Res. 2013, 47, 503–516. [Google Scholar] [CrossRef]
- Chisti, Y.; Moo-Young, M. Clean-in-place systems for industrial bioreactors: Design, validation and operation. J. Ind. Microbiol. 1994, 13, 201–207. [Google Scholar] [CrossRef]
- Joseph, B.; Otta, S.K.; Karunasagar, I.; Karunasagar, I. Biofilm formation by Salmonella spp. on food contact surfaces and their sensitivity to sanitizers. Int. J. Food Microbiol. 2001, 64, 367–372. [Google Scholar] [CrossRef]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef]
- Rodríguez, A.; McLandsborough, L.A. Evaluation of the transfer of Listeria monocytogenes from stainless steel and high-density polyethylene to Bologna and American cheese. J. Food Protect. 2007, 70, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Coulier, L.; Orbons, H.G.M.; Rijk, R. Analytical protocol to study the food safety of (multiple-)recycled high-density polyethylene (HDPE) and polypropylene (PP) crates: Influence of recycling on the migration and formation of degradation products. Polym. Degrad. Stab. 2007, 92, 2016–2025. [Google Scholar] [CrossRef]
- Moerman, F.; Partington, E. Materials of construction for food processing equipment and services: Requirements, strengths and weaknesses. J. Hyg. Eng. Des. 2014, 6, 10–37. [Google Scholar]
- Paul, E.; Ochoa, J.C.; Pechaud, Y.; Liu, Y.; Liné, A. Effect of shear stress and growth conditions on detachment and physical properties of biofilms. Water Res. 2012, 46, 5499–5508. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Simoes, L.C.; Pereira, M.O.; Vieira, M.J. Antagonism between Bacillus cereus and Pseudomonas fluorescens in planktonic systems and in biofilms. Biofouling 2008, 24, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tang, L.; Zhang, X.; Dong, J. A review of factors affecting the efficiency of clean-in-place procedures in closed processing systems. Energy 2019, 178, 57–71. [Google Scholar] [CrossRef]
- Moreira, J.M.R.; Simões, M.; Melo, L.F.; Mergulhão, F.J. The combined effects of shear stress and mass transfer on the balance between biofilm and suspended cell dynamics. Des. Water Treat. 2015, 53, 3348–3354. [Google Scholar] [CrossRef]
- Ochoa, J.-C.; Coufort, C.; Escudié, R.; Liné, A.; Paul, E. Influence of non-uniform distribution of shear stress on aerobic biofilms. Chem. Eng. Sci. 2007, 62, 3672–3684. [Google Scholar] [CrossRef]
- Mathieu, L.; Bertrand, I.; Abe, Y.; Angel, E.; Block, J.C.; Skali-Lami, S.; Francius, G. Drinking water biofilm cohesiveness changes under chlorination or hydrodynamic stress. Water Res. 2014, 55, 175–184. [Google Scholar] [CrossRef]
- Dogan, B.; Boor, K.J. Genetic diversity and spoilage potentials among Pseudomonas spp. isolated from fluid milk products and dairy processing plants. Appl. Environ. Microbiol. 2003, 69, 130–138. [Google Scholar] [CrossRef]
- Kreske, A.C.; Ryu, J.H.; Pettigrew, C.A.; Beuchat, L.R. Lethality of chlorine, chlorine dioxide, and a commercial produce sanitizer to Bacillus cereus and Pseudomonas in a liquid detergent, on stainless steel, and in biofilm. J. Food Protect. 2006, 69, 2621–2634. [Google Scholar] [CrossRef]
- Peng, J.-S.; Tsai, W.-C.; Chou, C.-C. Inactivation and removal of Bacillus cereus by sanitizer and detergent. Int. J. Food Microbiol. 2002, 77, 11–18. [Google Scholar] [CrossRef]
- Ferreira, C.; Rosmaninho, R.; Simões, M.; Pereira, M.C.; Bastos, M.M.; Nunes, O.C.; Coelho, M.; Melo, L.F. Biofouling control using microparticles carrying a biocide. Biofouling 2010, 26, 205–212. [Google Scholar] [CrossRef]
- Simões, L.C.; Azevedo, N.; Pacheco, A.; Keevil, C.W.; Vieira, M.J. Drinking water biofilm assessment of total and culturable bacteria under different operating conditions. Biofouling 2006, 22, 91–99. [Google Scholar] [CrossRef]
- Lemos, M.; Gomes, I.; Mergulhão, F.; Melo, L.; Simões, M. The effects of surface type on the removal of Bacillus cereus and Pseudomonas fluorescens single and dual species biofilms. Food Bioprod. Process. 2015, 93, 234–241. [Google Scholar] [CrossRef][Green Version]
- Bayoudh, S.; Othmane, A.; Mora, L.; Ben Ouada, H. Assessing bacterial adhesion using DLVO and XDLVO theories and the jet impingement technique. Colloids Surf. B Biointerfaces 2009, 73, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Busscher, H.J.; Weerkamp, A.H.; van der Mei, H.C.; van Pelt, A.W.; de Jong, H.P.; Arends, J. Measurement of the surface free energy of bacterial cell surfaces and its relevance for adhesion. Appl. Environ. Microbiol. 1984, 48, 980–983. [Google Scholar] [CrossRef]
- Simões, M.; Simões, L.C.; Cleto, S.; Machado, I.; Pereira, M.O.; Vieira, M.J. Antimicrobial mechanisms of ortho-phthalaldehyde action. J. Basic Microbiol. 2007, 47, 230–242. [Google Scholar] [CrossRef]
- Janczuk, B.; Chibowski, E.; Bruque, J.M.; Kerkeb, M.L.; Caballero, F.G. On the consistency of surface free energy components as calculated from contact angles of different liquids: An application to the cholesterol surface. J. Colloid Interface Sci. 1993, 159, 421–428. [Google Scholar] [CrossRef]
- Simões, L.C.; Simões, M.; Oliveira, R.; Vieira, M.J. Potential of the adhesion of bacteria isolated from drinking water to materials. J. Basic Microbiol. 2007, 47, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Simões, L.C.; Cleto, S.; Pereira, M.O.; Vieira, M.J. The effects of a biocide and a surfactant on the detachment of Pseudomonas fluorescens from glass surfaces. Int. J. Food Microbiol. 2008, 121, 335–341. [Google Scholar] [CrossRef][Green Version]
- Johnston, M.D.; Lambert, R.J.; Hanlon, G.W.; Denyer, S.P. A rapid method for assessing the suitability of quenching agents for individual biocides as well as combinations. J. Appl. Microbiol. 2002, 92, 784–789. [Google Scholar] [CrossRef]
- Simões, M.; Pereira, M.O.; Sillankorva, S.; Azeredo, J.; Vieira, M.J. The effect of hydrodynamic conditions on the phenotype of Pseudomonas fluorescens biofilms. Biofouling 2007, 23, 249–258. [Google Scholar] [CrossRef]
- Lemos, M.; Mergulhão, F.; Melo, L.; Simões, M. The effect of shear stress on the formation and removal of Bacillus cereus biofilms. Food Bioprod. Process. 2015, 93, 242–248. [Google Scholar] [CrossRef]
- Geankoplis, C.J. Transport. Processes and Unit Operations; Prentice-Hall International, Inc.: Hoboken, NJ, USA, 1993. [Google Scholar]
- Perry, R.H.; Green, D.W. Perry’s Chemical Engineers’ Handbook; McGraw-Hill Professional Publishing: New York, NY, USA, 1999. [Google Scholar]
- Gabe, D.R.; Walsh, F.C. The rotating cylinder electrode: A review of development. J. Appl. Electrochem. 1983, 13, 3–21. [Google Scholar] [CrossRef]
- Simões, M.; Pereira, M.O.; Vieira, M.J. Effect of mechanical stress on biofilms challenged by different chemicals. Water Res. 2005, 39, 5142–5152. [Google Scholar] [CrossRef]
- Teodósio, J.S.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Flow cell hydrodynamics and their effects on E. coli biofilm formation under different nutrient conditions and turbulent flow. Biofouling 2011, 27, 1–11. [Google Scholar] [CrossRef]
- Frølund, B.; Palmgren, R.; Keiding, K.; Nielsen, P.H. Extraction of extracellular polymers from activated sludge using a cation exchange resin. Water Res. 1996, 30, 1749–1758. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Peterson, G.L. Review of the folin phenol protein quantitation method of lowry, rosebrough, farr and randall. Anal. Biochem. 1979, 100, 201–220. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- APHA; AWWA; WPCF. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA; American Water Works Association: Denver, CO, USA; Water Environment Federation: Alexandria, VA, USA, 1989. [Google Scholar]
- Melo, L.F.; Bott, T.R. Biofouling in water systems. Exp. Therm. Fluid Sci. 1997, 14, 375–381. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed]
- Busscher, H.J.; van der Mei, H.C. How do bacteria know they are on a surface and regulate their response to an adhering state? PLoS Pathog. 2012, 8, e1002440. [Google Scholar] [CrossRef] [PubMed]
- Bullitt, E.; Makowski, L. Structural polymorphism of bacterial adhesion pili. Nature 1995, 373, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.E.; Nilsson, L.M.; Forero, M.; Sokurenko, E.V.; Vogel, V. Shear-dependent ‘stick-and-roll’ adhesion of type 1 fimbriated Escherichia coli. Mol. Microbiol. 2004, 53, 1545–1557. [Google Scholar] [CrossRef] [PubMed]
- An, Y.H.; Friedman, R.J. Concise review of mechanisms of bacterial adhesion to biomaterial surfaces. J. Biomed. Mat. Res. 1998, 43, 338–348. [Google Scholar] [CrossRef]
- Busscher, H.J.; van der Mei, H.C. Microbial adhesion in flow displacement systems. Clin. Microbiol. Rev. 2006, 19, 127–141. [Google Scholar] [CrossRef]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Melo, L.F. Biofilm physical structure, internal diffusivity and tortuosity. Water Sci. Technol. 2005, 52, 77–84. [Google Scholar] [CrossRef]
- Stewart, P.S. Theoretical aspects of antibiotic diffusion into microbial biofilms. Antimicrob. Agents Chemother. 1996, 40, 2517–2522. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S. Diffusion in Biofilms. J. Bacteriol. 2003, 185, 1485. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Stewart, P.S. Biofilm removal caused by chemical treatments. Water Res. 2000, 34, 4229–4233. [Google Scholar] [CrossRef]
- Behnke, S.; Camper, A.K. Chlorine dioxide disinfection of single and dual species biofilms, detached biofilm and planktonic cells. Biofouling 2012, 28, 635–647. [Google Scholar] [CrossRef]
- Hornemann, J.A.; Lysova, A.A.; Codd, S.L.; Seymour, J.D.; Busse, S.C.; Stewart, P.S.; Brown, J.R. Biopolymer and water dynamics in microbial biofilm extracellular polymeric substance. Biomacromolecules 2008, 9, 2322–2328. [Google Scholar] [CrossRef]
- Pechaud, Y.; Marcato-Romain, C.E.; Girbal-Neuhauser, E.; Queinnec, I.; Bessiere, Y.; Paul, E. Combining hydrodynamic and enzymatic treatments to improve multi-species thick biofilm removal. Chem. Eng. Sci. 2012, 80, 109–118. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (s−1) | ||
|---|---|---|
| 3.84 | 2400 | 0.70 |
| 6.40 | 4000 | 1.66 |
| 13.0 | 8100 | 5.50 |
| 19.4 | 12,100 | 10.9 |
| 25.8 | 16,000 | 17.7 |
| B. cereus | P. fluorescens | |||
|---|---|---|---|---|
| Control | BDMDAC | Control | BDMDAC | |
| HDPE | −13.1 | −6.5 | 7.8 | −5.8 |
| B. cereus | P. fluorescens | Dual Species | |
|---|---|---|---|
| Thickness (µm) | 526 ± 8 | 278 ± 71 | 880 ± 90 |
| Dry mass (mg/cm2) | 0.191 ± 0.02 | 0.365 ± 0.07 | 0.324 ± 0.09 |
| Volumetric density (mg/cm3) | 3.6 ± 0.3 | 13.1 ± 1.9 | 3.7 ± 1.1 |
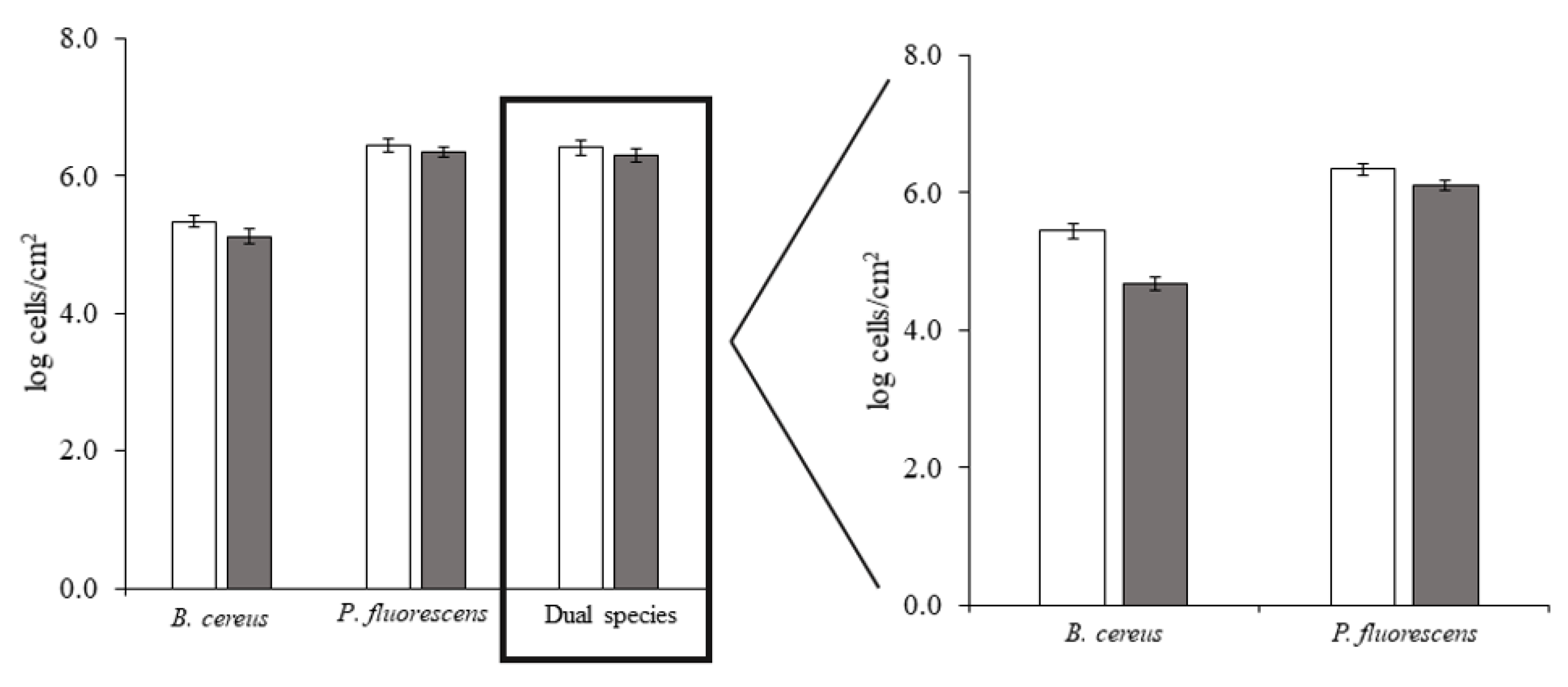
| Cellular density (log cells/cm2) | 6.67 ± 0.11 | 6.94 ± 0.20 | 6.90 ± 0.04 * |
| Water content (% of total biofilm mass) | 97.9 ± 0.7 | 95.0 ± 3.1 | 99.3 ± 2.5 |
| Extracellular polysaccharides (% of total biofilm polysaccharides) | 72.1 ± 1.5 | 69.8 ± 2.1 | 59.4 ± 0.7 |
| Extracellular proteins (% of total biofilm proteins) | 50.9 ± 0.5 | 34.3 ± 0.6 | 72.0 ± 4.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, I.B.; Lemos, M.; Fernandes, S.; Borges, A.; Simões, L.C.; Simões, M. The Effects of Chemical and Mechanical Stresses on Bacillus cereus and Pseudomonas fluorescens Single- and Dual-Species Biofilm Removal. Microorganisms 2021, 9, 1174. https://doi.org/10.3390/microorganisms9061174
Gomes IB, Lemos M, Fernandes S, Borges A, Simões LC, Simões M. The Effects of Chemical and Mechanical Stresses on Bacillus cereus and Pseudomonas fluorescens Single- and Dual-Species Biofilm Removal. Microorganisms. 2021; 9(6):1174. https://doi.org/10.3390/microorganisms9061174
Chicago/Turabian StyleGomes, Inês B., Madalena Lemos, Susana Fernandes, Anabela Borges, Lúcia C. Simões, and Manuel Simões. 2021. "The Effects of Chemical and Mechanical Stresses on Bacillus cereus and Pseudomonas fluorescens Single- and Dual-Species Biofilm Removal" Microorganisms 9, no. 6: 1174. https://doi.org/10.3390/microorganisms9061174
APA StyleGomes, I. B., Lemos, M., Fernandes, S., Borges, A., Simões, L. C., & Simões, M. (2021). The Effects of Chemical and Mechanical Stresses on Bacillus cereus and Pseudomonas fluorescens Single- and Dual-Species Biofilm Removal. Microorganisms, 9(6), 1174. https://doi.org/10.3390/microorganisms9061174