
Characterization of the RNA-Binding Protein TcSgn1 in Trypanosoma cruzi




Abstract
1. Introduction
2. Materials and Methods
2.1. Cloning and Expression in T. cruzi
2.2. T. cruzi Growth Conditions and Cloning
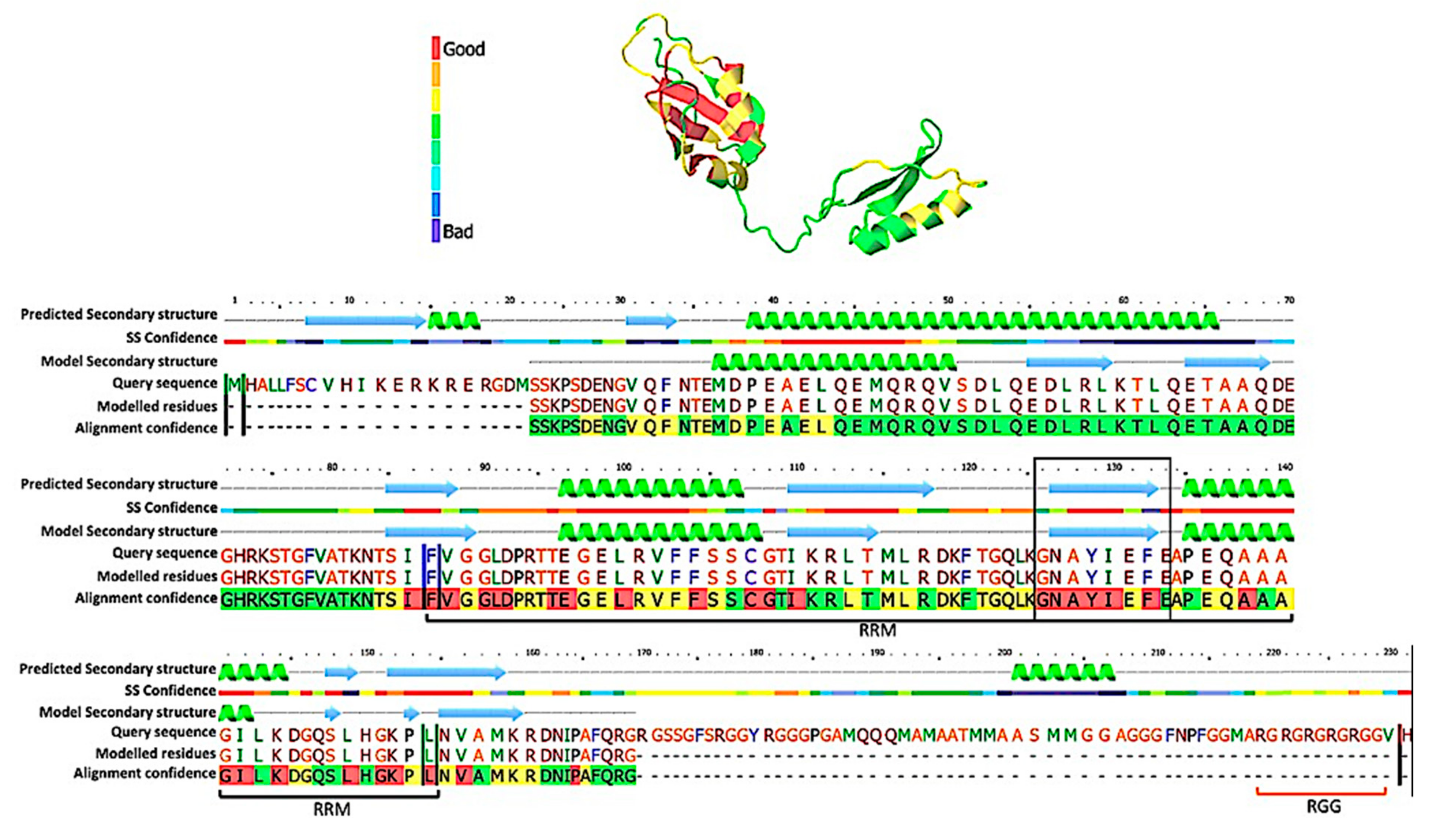
2.3. Bioinformatics/Structural Predictions
2.4. Immunoprecipitation of Protein Complexes Associated with TcSgn1
2.5. Fluorescent RNA Electrophoresis Mobility Shift Assay (RNA-EMSA)
2.6. RNA-Seq Data Analysis
3. Results
3.1. TcSgn1 Is a Nuclear RNA-Binding Protein
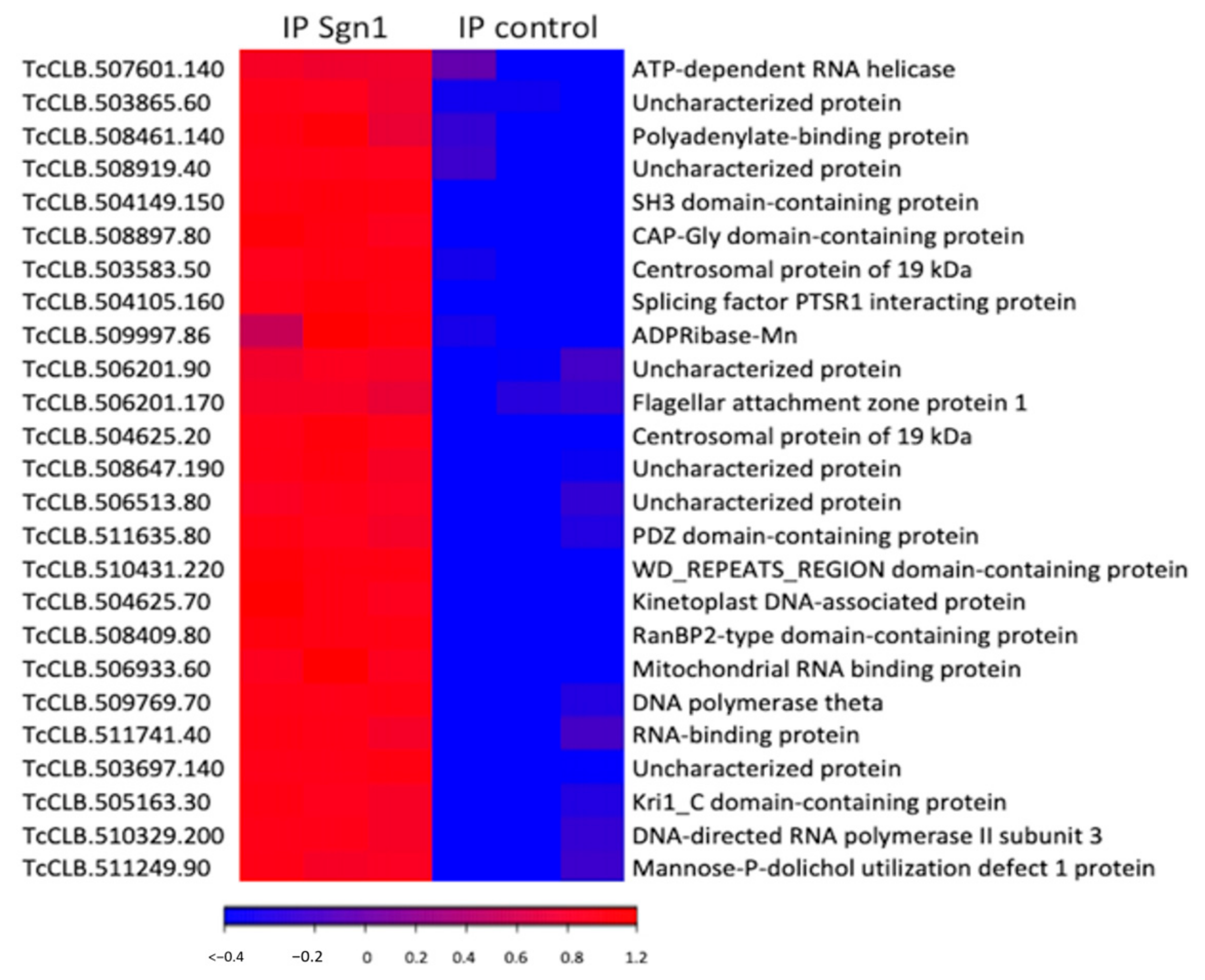
3.2. TcSgn1 Ribonucleoprotein Complex
3.3. TcSgn1 Binds to the Poly(A) Tail of mRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chagas, C. Nova Tripanozomiaze Humana: Estudos Sobre a Morfolojia e o Ciclo Evolutivo Do Schizotrypanum Cruzi n. Gen., n. Sp., Ajente Etiolojico de Nova Entidade Morbida Do Homem. Mem. Inst. Oswaldo Cruz 1909, 1, 159–218. [Google Scholar] [CrossRef]
- De Souza, W. Cell Biology of Trypanosoma Cruzi. Int. Rev. Cytol. 1984. [Google Scholar] [CrossRef]
- de Souza, W.; de Carvalho, T.M.; Barrias, E.S. Review on Trypanosoma Cruzi: Host Cell Interaction. Int. J. Cell. Biol. 2010, 2010, 295394. [Google Scholar] [CrossRef] [PubMed]
- Clayton, C.E. Life without Transcriptional Control? From Fly to Man and Back Again. EMBO J. 2002, 21, 1881–1888. [Google Scholar] [CrossRef]
- Clayton, C. The Regulation of Trypanosome Gene Expression by RNA-Binding Proteins. PLoS Pathog. 2013, 9, e1003680. [Google Scholar] [CrossRef]
- Clayton, C.; Shapira, M. Post-Transcriptional Regulation of Gene Expression in Trypanosomes and Leishmanias. Mol. Biochem. Parasitol. 2007, 156, 93–101. [Google Scholar] [CrossRef]
- Romagnoli, B.A.A.; Holetz, F.B.; Alves, L.R.; Goldenberg, S. RNA Binding Proteins and Gene Expression Regulation in Trypanosoma Cruzi. Front. Cell. Infect. Microbiol. 2020. [Google Scholar] [CrossRef]
- Palenchar, J.B.; Bellofatto, V. Gene Transcription in Trypanosomes. Mol. Biochem. Parasitol. 2006, 146, 135–141. [Google Scholar] [CrossRef]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A Brave New World of RNA-Binding Proteins. Nat. Rev. Mol. Cell. Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Keene, J.D. RNA Regulons: Coordination of Post-Transcriptional Events. Nat. Rev. Genet. 2007, 8, 533–543. [Google Scholar] [CrossRef]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A Census of Human RNA-Binding Proteins. Nat. Rev. Genet. 2014. [Google Scholar] [CrossRef]
- Cléry, A.; Frédéric, H.-T.A. From Structure to Function of RNA Binding Domains. Landes Biosci. Austin 2012, 137, 146–158. [Google Scholar]
- Oliveira, C.; Faoro, H.; Alves, L.R.; Goldenberg, S. RNA-Binding Proteins and Their Role in the Regulation of Gene Expression in Trypanosoma Cruzi and Saccharomyces Cerevisiae. Genet. Mol. Biol. 2017, 40. [Google Scholar] [CrossRef]
- Winstall, E.; Sadowski, M.; Kühn, U.; Wahle, E.; Sachs, A.B. The Saccharomyces Cerevisiae RNA-Binding Protein Rbp29 Functions in Cytoplasmic MRNA Metabolism. J. Biol. Chem. 2000, 275, 21817–21826. [Google Scholar] [CrossRef]
- Hogan, D.J.; Riordan, D.P.; Gerber, A.P.; Herschlag, D.; Brown, P.O. Diverse RNA-Binding Proteins Interact with Functionally Related Sets of RNAs, Suggesting an Extensive Regulatory System. PLoS Biol. 2008, 6, e255. [Google Scholar] [CrossRef]
- Alves, L.R.; Avila, A.R.; Correa, A.D.; Holetz, F.B.; Mansur, F.C.; Manque, P.A.; Menezes, J.P.B.; Buck, G.; Krieger, M.A.; Goldenberg, S. Proteomic Analysis Reveals the Dynamic Associationof Proteins with Translated MRNAs in Trypanosoma Cruzi. Gene 2010, 1, 6. [Google Scholar]
- Batista, M.; Marchini, F.K.; Celedon, P.A.; Fragoso, S.P.; Probst, C.M.; Preti, H.; Ozaki, L.S.; Buck, G.A.; Goldenberg, S.; Krieger, M.A. A High-Throughput Cloning System for Reverse Genetics in Trypanosoma Cruzi. BMC Microbiol. 2010, 10, 259. [Google Scholar] [CrossRef]
- Oliveira, C.; Carvalho, P.C.; Alves, L.R.; Goldenberg, S. The Role of the Trypanosoma Cruzi TcNRBD1 Protein in Translation. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Contreras, V.T.; Araujo-Jorge, T.C.; Bonaldo, M.C.; Thomaz, N.; Barbosa, H.S.; Meirelles, M.N.; Goldenberg, S. Biological Aspects of the Dm 28c Clone of Trypanosoma Cruzi after Metacyclogenesis in Chemically Defined Media. Mem. Inst. Oswaldo Cruz 1988, 83, 123–133. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Fridy, P.C.; Li, Y.; Keegan, S.; Thompson, M.K.; Nudelman, I.; Scheid, J.F.; Oeffinger, M.; Nussenzweig, M.C.; Fenyö, D.; Chait, B.T.; et al. A Robust Pipeline for Rapid Production of Versatile Nanobody Repertoires. Nat. Methods 2014, 11, 1253–1260. [Google Scholar] [CrossRef]
- Hakhverdyan, Z.; Domanski, M.; Hough, L.E.; Oroskar, A.A.; Oroskar, A.R.; Keegan, S.; Dilworth, D.J.; Molloy, K.R.; Sherman, V.; Aitchison, J.D.; et al. Rapid, Optimized Interactomic Screening. Nat. Methods 2015, 12, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.R.; Oliveira, C.; Mörking, P.A.; Kessler, R.L.; Martins, S.T.; Romagnoli, B.A.A.; Marchini, F.K.; Goldenberg, S. The MRNAs Associated to a Zinc Finger Protein from Trypanosoma Cruzi Shift during Stress Conditions. RNA Biol. 2014, 11, 921–933. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A Scaling Normalization Method for Differential Expression Analysis of RNA-Seq Data. Genome Biol. 2010. [Google Scholar] [CrossRef]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-Throughput Functional Annotation and Data Mining with the Blast2GO Suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New Perspectives on Genomes, Pathways, Diseases and Drugs. Nucleic Acids Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019. [Google Scholar] [CrossRef]
- de Godoy, L.M.; Marchini, F.K.; Pavoni, D.P.; Rampazzo, R.C.P.; Probst, C.M.; Goldenberg, S.; Krieger, M.A. Quantitative Proteomics of Trypanosoma Cruzi during Metacyclogenesis. Proteomics 2012, 12, 2694–2703. [Google Scholar] [CrossRef]
- Holetz, F.B.; Alves, L.R.; Probst, C.M.; Dallagiovanna, B.; Marchini, F.K.; Manque, P.; Buck, G.; Krieger, M.A.; Correa, A.; Goldenberg, S. Protein and MRNA Content of TcDHH1-Containing MRNPs in Trypanosoma Cruzi. FEBS J. 2010, 277. [Google Scholar] [CrossRef]
- Holetz, F.B.; Correa, A.; Avila, A.R.; Nakamura, C.V.; Krieger, M.A.; Goldenberg, S. Evidence of P-Body-like Structures in Trypanosoma Cruzi. Biochem. Biophys. Res. Commun. 2007, 356, 1062–1067. [Google Scholar] [CrossRef]
- Smircich, P.; Eastman, G.; Bispo, S.; Duhagon, M.A.; Guerra-Slompo, E.P.; Garat, B.; Goldenberg, S.; Munroe, D.J.; Dallagiovanna, B.; Holetz, F.; et al. Ribosome Profiling Reveals Translation Control as a Key Mechanism Generating Differential Gene Expression in Trypanosoma Cruzi. BMC Genom. 2015, 16. [Google Scholar] [CrossRef]
- Nadler, J.P.; Lederer, E.; Wittner, M.; Baum, S.G.; Tanowitz, H.B. The Effect of Adenosine Analogues on the in Vitro Growth of Trypanosoma Cruzi. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 285–287. [Google Scholar] [CrossRef]
- Avila, J.L.; Polegre, M.A.; Avila, A. Inhibitory Effects of Sinefungin and Its Cyclic Analog on the Multiplication of Trypanosoma Cruzi Isolates. Am. J. Trop. Med. Hyg. 1993, 48, 112–119. [Google Scholar] [CrossRef]
- da Costa Lima, T.D.; Moura, D.M.N.; Reis, C.R.S.; Vasconcelos, J.R.C.; Ellis, L.; Carrington, M.; Figueiredo, R.C.B.Q.; de Melo Neto, O.P. Functional Characterization of Three Leishmania Poly(A) Binding Protein Homologues with Distinct Binding Properties to RNA and Protein Partners. Eukaryot. Cell 2010. [Google Scholar] [CrossRef]
- Alves, L.R. RNA-Binding Proteins Related to Stress Response and Differentiation in Protozoa. World J. Biol. Chem. 2016, 7, 78. [Google Scholar] [CrossRef]
- da Costa, J.F.; Ferrarini, M.G.; Nardelli, S.C.; Goldenberg, S.; Ávila, A.R.; Holetz, F.B. Trypanosoma Cruzi XRNA Granules Colocalise with Distinct MRNP Granules at the Nuclear Periphery. Mem. Inst. Oswaldo Cruz 2018. [Google Scholar] [CrossRef]
- Sotelo, N.S.; Schepens, J.T.G.; Valiente, M.; Hendriks, W.J.A.J.; Pulido, R. PTEN–PDZ Domain Interactions: Binding of PTEN to PDZ Domains of PTPN13. Methods 2015, 77–78, 147–156. [Google Scholar] [CrossRef]
- Burri, L.; Lithgow, T. A Complete Set of SNAREs in Yeast: A Complete Set of SNAREs in Yeast. Traffic 2004, 5, 45–52. [Google Scholar] [CrossRef]
- Cunha-e-Silva, N.; Sant’Anna, C.; Pereira, M.G.; Porto-Carreiro, I.; Jeovanio, A.L.; de Souza, W. Reservosomes: Multipurpose Organelles? Parasitol. Res. 2006, 99, 325–327. [Google Scholar] [CrossRef]
- Zoltner, M.; Krienitz, N.; Field, M.C.; Kramer, S. Comparative Proteomics of the Two T. Brucei PABPs Suggests That PABP2 Controls Bulk MRNA. PLoS Negl. Trop. Dis. 2018, 12, e0006679. [Google Scholar] [CrossRef]
- Sharan, R.; Suthram, S.; Kelley, R.M.; Kuhn, T.; McCuine, S.; Uetz, P.; Sittler, T.; Karp, R.M.; Ideker, T. Conserved Patterns of Protein Interaction in Multiple Species. Proc. Natl. Acad. Sci. USA 2005, 102, 1974–1979. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Term | p Value | Fold Enrichment | FDR | Fisher Exact |
|---|---|---|---|---|---|
| GOTERM_MF_DIRECT | Nucleic acid binding | 4.10 × 10−9 | 2.5 | 0% | 1.50 × 10−9 |
| GOTERM_BP_DIRECT | Transcription, DNA-templated | 5.20 × 10−5 | 3.4 | 1% | 1.30 × 10−5 |
| GOTERM_MF_DIRECT | RNA binding | 1.10 × 10−4 | 2.3 | 1% | 4.50 × 10−5 |
| GOTERM_MF_DIRECT | DNA-directed RNA polymerase activity | 9.60 × 10−4 | 3.2 | 5% | 2.50 × 10−4 |
| KEGG_PATHWAY | Endocytosis | 1.70 × 10−3 | 2.9 | 8% | 5.00 × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, C.; Gerber, A.P.; Goldenberg, S.; Alves, L.R. Characterization of the RNA-Binding Protein TcSgn1 in Trypanosoma cruzi. Microorganisms 2021, 9, 986. https://doi.org/10.3390/microorganisms9050986
Oliveira C, Gerber AP, Goldenberg S, Alves LR. Characterization of the RNA-Binding Protein TcSgn1 in Trypanosoma cruzi. Microorganisms. 2021; 9(5):986. https://doi.org/10.3390/microorganisms9050986
Chicago/Turabian StyleOliveira, Camila, André P. Gerber, Samuel Goldenberg, and Lysangela R. Alves. 2021. "Characterization of the RNA-Binding Protein TcSgn1 in Trypanosoma cruzi" Microorganisms 9, no. 5: 986. https://doi.org/10.3390/microorganisms9050986
APA StyleOliveira, C., Gerber, A. P., Goldenberg, S., & Alves, L. R. (2021). Characterization of the RNA-Binding Protein TcSgn1 in Trypanosoma cruzi. Microorganisms, 9(5), 986. https://doi.org/10.3390/microorganisms9050986